
Expression of Nerve Growth Factor and Its Receptor TrkA in the Reproductive System of Adult Zebrafish


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Animals and Gonads Dissection
2.2. Histology (Haematoxylin and Eosin Staining)
2.3. Chromogenic and Fluorescence In Situ Hybridization (ISH)
- ngf F: GGAGCACAGGAGATCTACGC and R: CGTGGAAAAACCCAACTCAT;
- trka F: AGTTGTTGCTTGCAGGGTGG and R: TGGGTCAATCATGACCTCAG.
3. Results
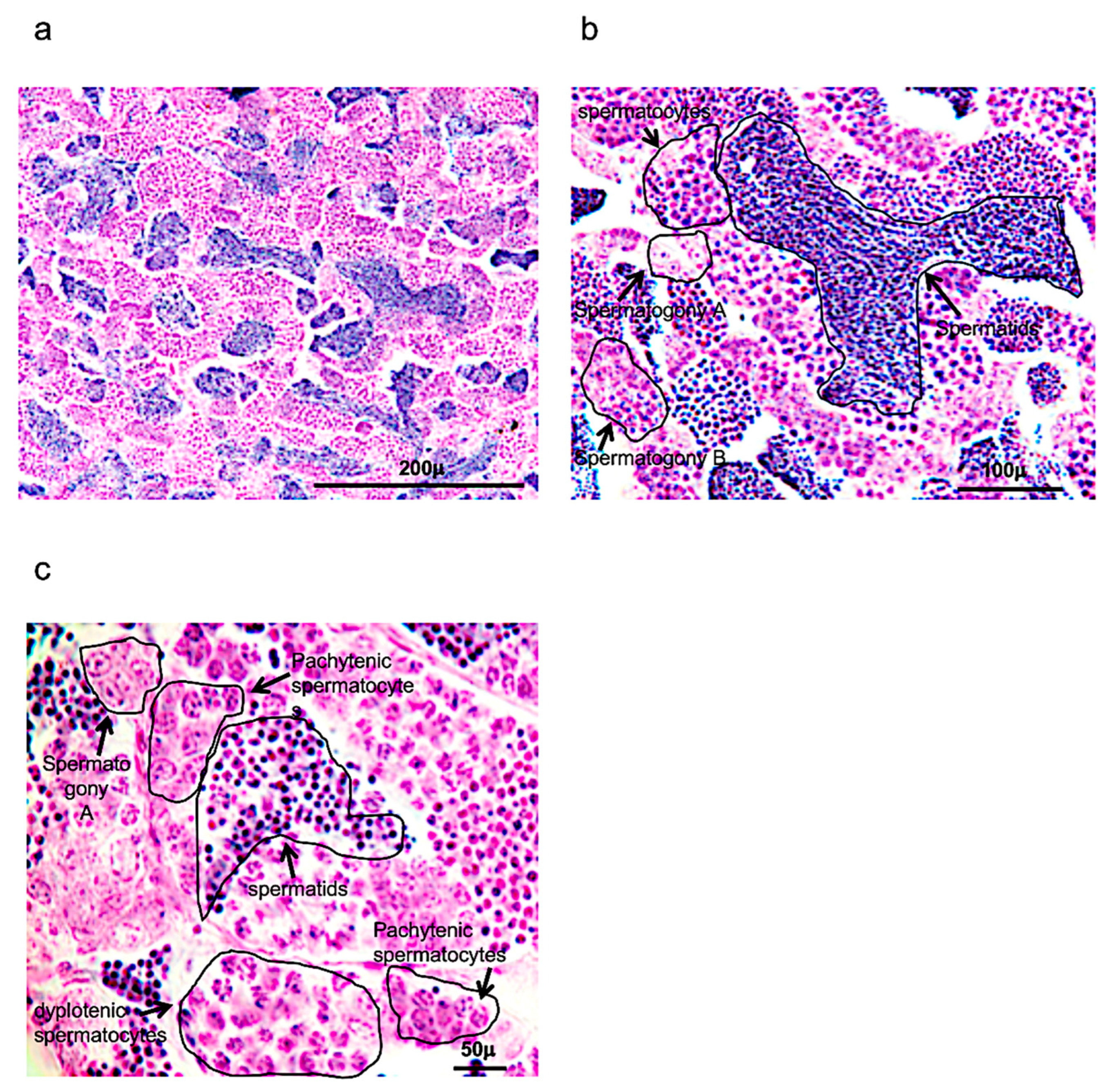
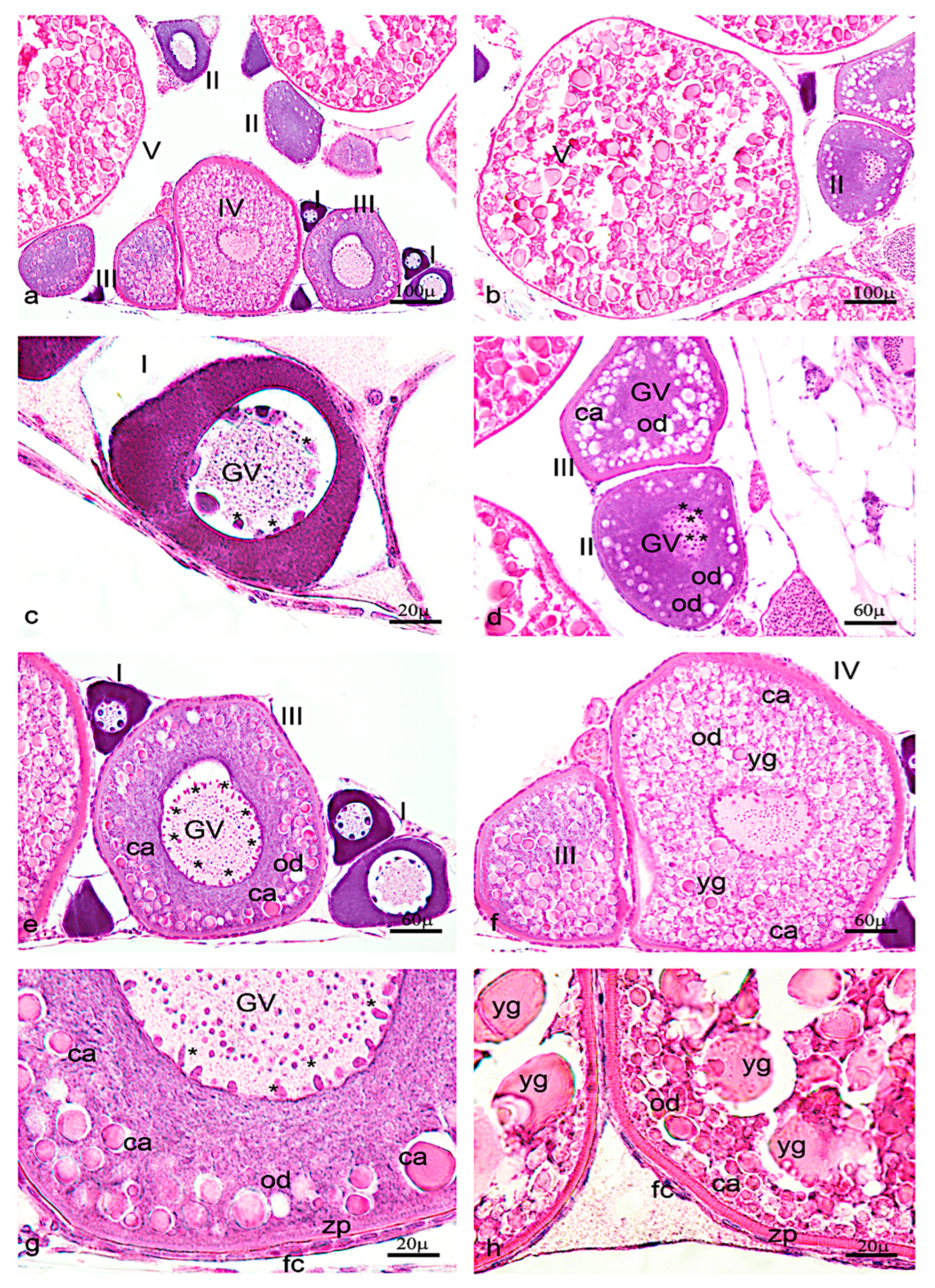
3.1. Morphological Analysis of Adult Zebrafish Testis
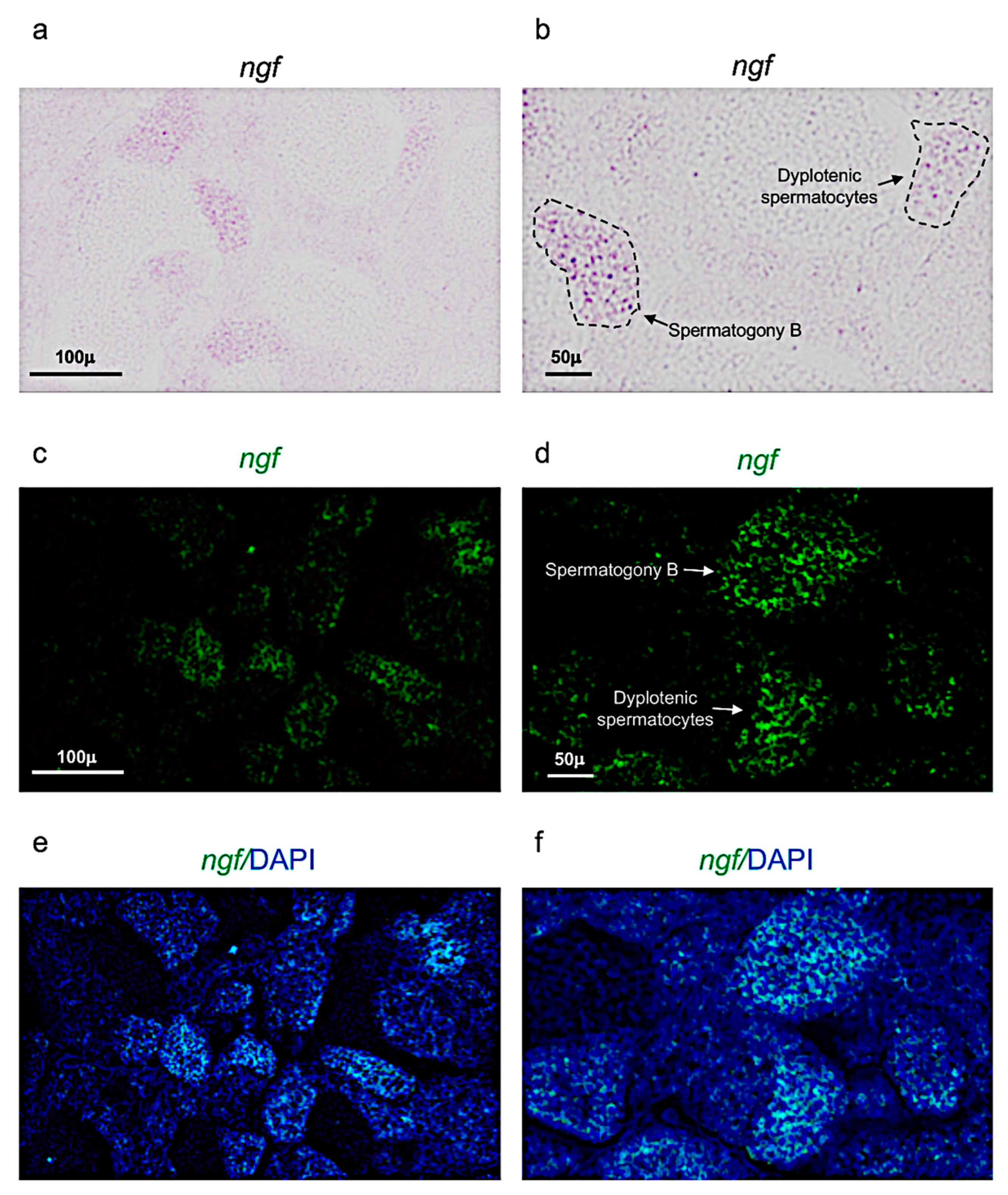
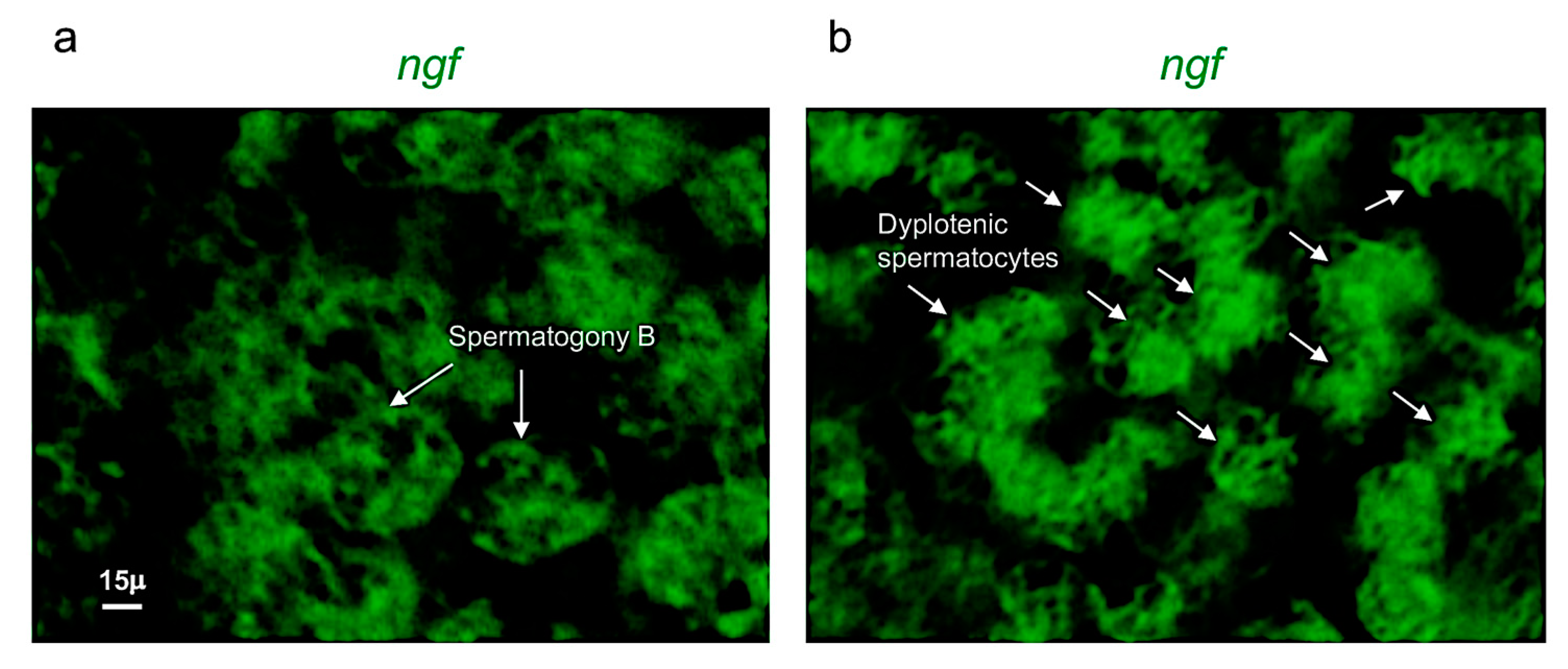
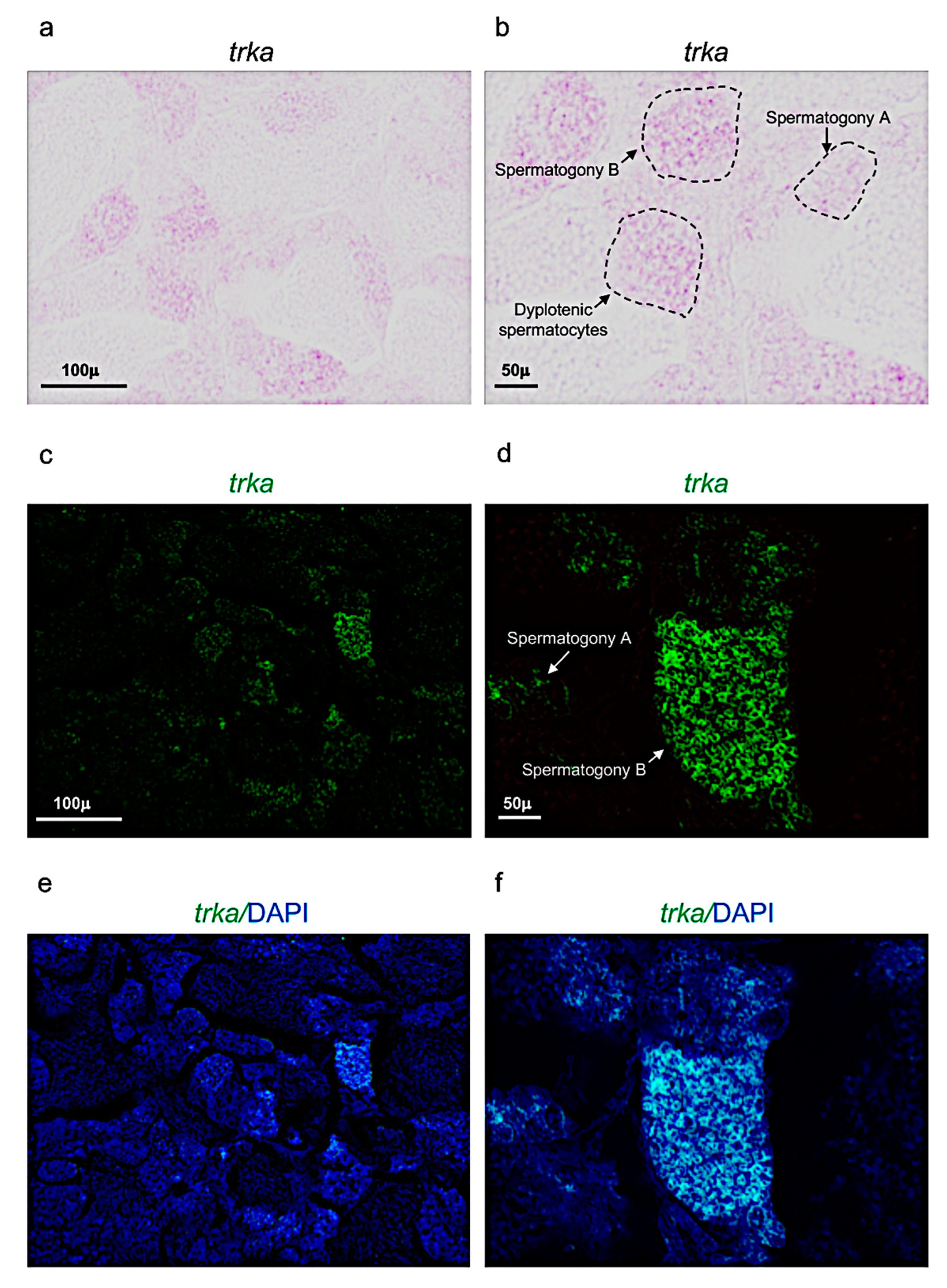
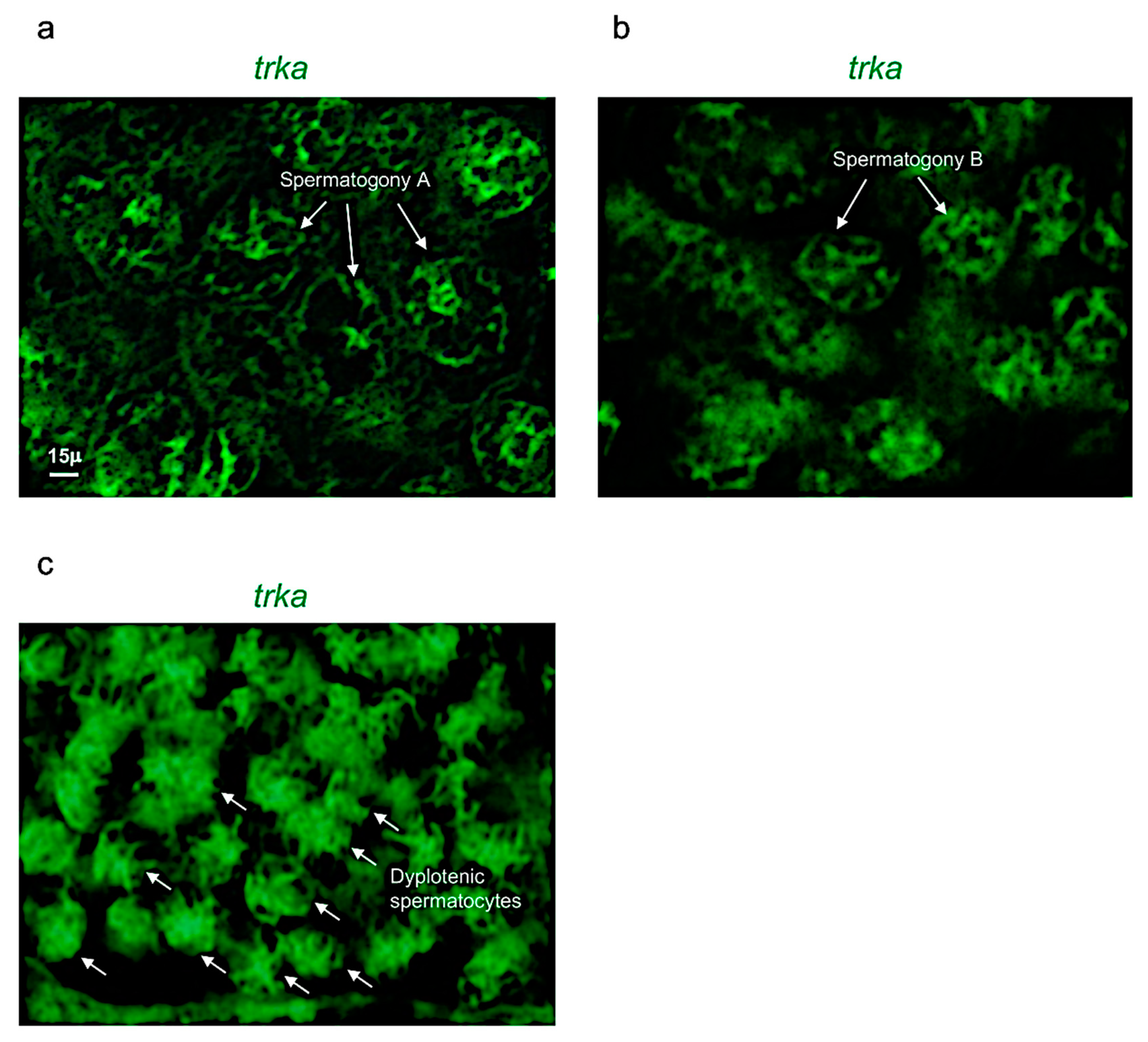
3.2. Ngf and Its Receptor trka Are Expressed in Adult Zebrafish Testis
3.3. Morphological Analysis of Zebrafish Ovary
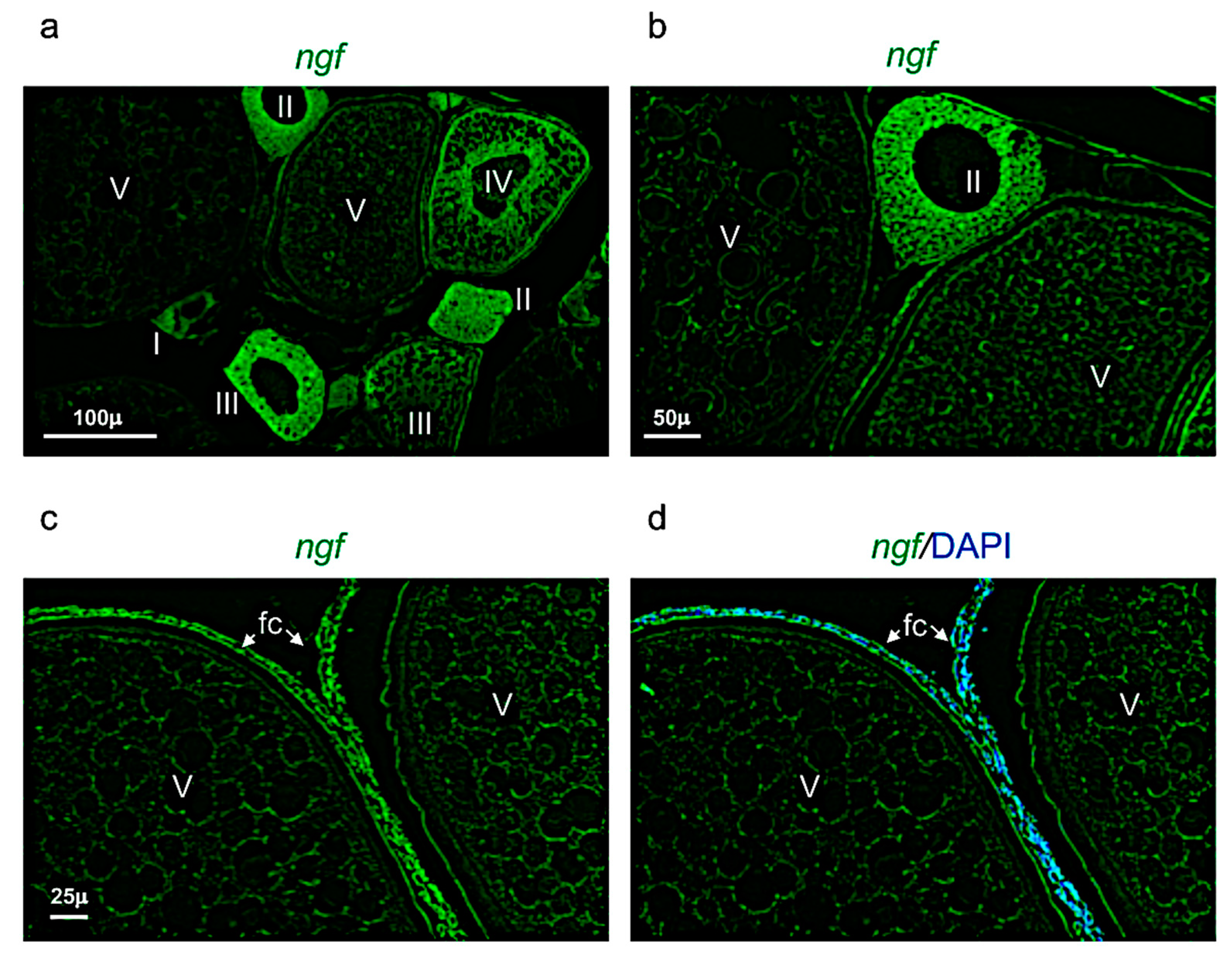
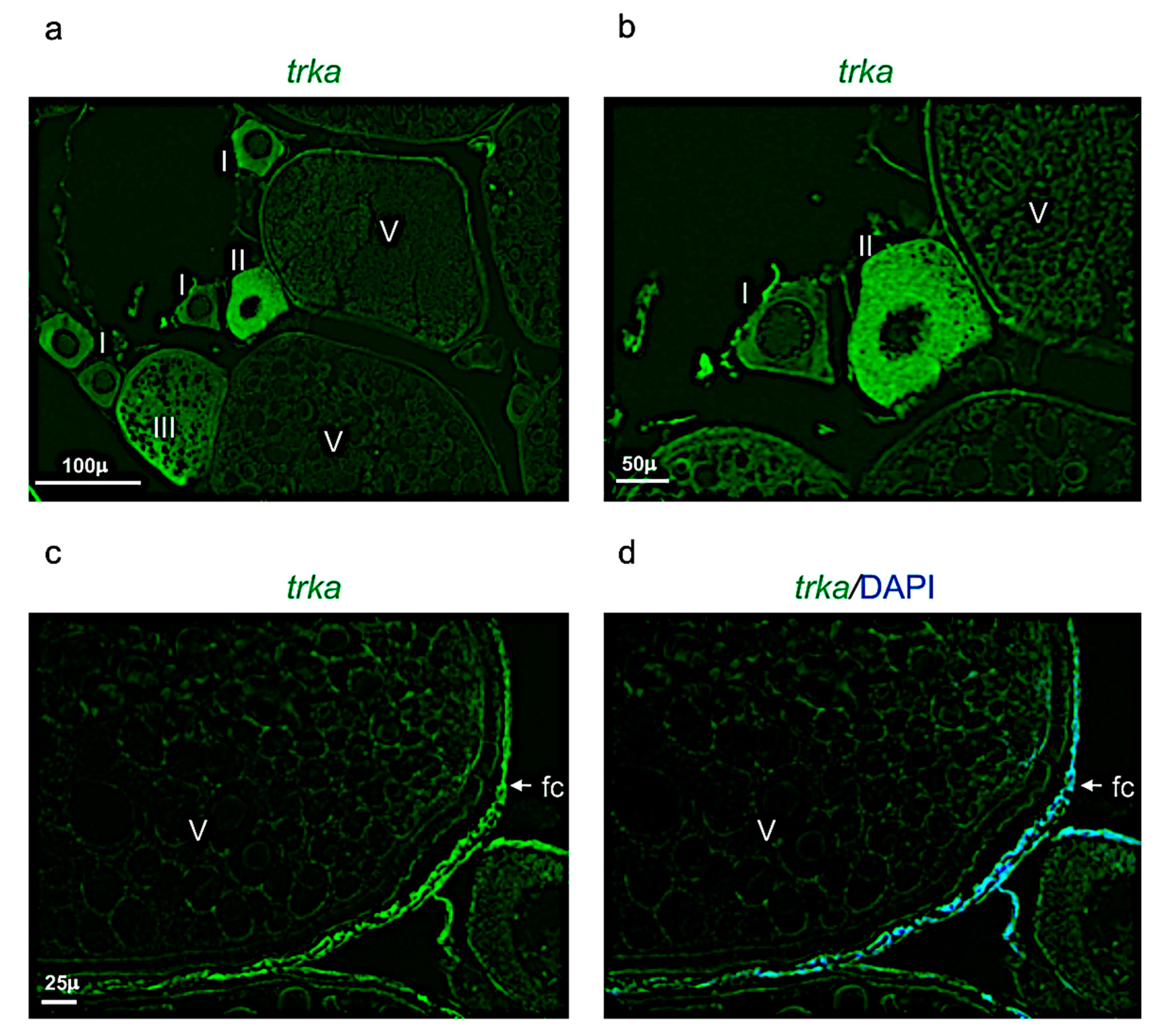
3.4. Ngf and Its Receptor Trka Are Expressed in Adult Zebrafish Ovary
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Montalcini, R.L. Laboratory of Cellular Biology. Department of Neurobiology, Rome. Ric. Sci. 1970, 40, 296–298. [Google Scholar] [PubMed]
- Aloe, L. Rita Levi-Montalcini and the discovery of NGF, the first nerve cell growth factor. Arch. Ital. Biol. 2011, 149, 175–181. [Google Scholar] [PubMed]
- Park, H.; Poo, M.M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2013, 14, 7–23. [Google Scholar] [CrossRef]
- McPhee, G.M.; Downey, L.A.; Stough, C. Neurotrophins as a reliable biomarker for brain function, structure and cognition: A systematic review and meta-analysis. Neurobiol. Learn. Mem 2020, 175, 107298. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in neuronal development and function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [PubMed]
- Chao, M.V. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299–309. [Google Scholar] [CrossRef]
- Chao, M.V. Dependence receptors: What is the mechanism? Sci. STKE 2003, 2003, PE38. [Google Scholar] [CrossRef]
- Chao, M.V. Retrograde transport redux. Neuron 2003, 39, 1–2. [Google Scholar] [CrossRef]
- Hallbook, F.; Wilson, K.; Thorndyke, M.; Olinski, R.P. Formation and evolution of the chordate neurotrophin and Trk receptor genes. Brain Behav. Evol. 2006, 68, 133–144. [Google Scholar] [CrossRef]
- Hallbook, F. Evolution of the vertebrate neurotrophin and Trk receptor gene families. Curr. Opin. Neurobiol. 1999, 9, 616–621. [Google Scholar] [CrossRef]
- Cacialli, P.; Gatta, C.; D’Angelo, L.; Leggieri, A.; Palladino, A.; de Girolamo, P.; Pellegrini, E.; Lucini, C. Nerve growth factor is expressed and stored in central neurons of adult zebrafish. J. Anat. 2019, 235, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Cacialli, P.; Lucini, C. Adult neurogenesis and regeneration in zebrafish brain: Are the neurotrophins involved in? Neural Regen. Res. 2019, 14, 2067–2068. [Google Scholar] [PubMed]
- Cacialli, P. Neurotrophins Time Point Intervention after Traumatic Brain Injury: From Zebrafish to Human. Int. J. Mol. Sci. 2021, 22, 1585. [Google Scholar] [CrossRef] [PubMed]
- Hartman, D.S.; McCormack, M.; Schubenel, R.; Hertel, C. Multiple trkA proteins in PC12 cells bind NGF with a slow association rate. J. Biol. Chem. 1992, 267, 24516–24522. [Google Scholar] [CrossRef]
- Jung, K.M.; Tan, S.; Landman, N.; Petrova, K.; Murray, S.; Lewis, R.; Kim, P.K.; Kim, D.S.; Ryu, S.H.; Chao, M.V.; et al. Regulated intramembrane proteolysis of the p75 neurotrophin receptor modulates its association with the TrkA receptor. J. Biol. Chem. 2003, 278, 42161–42169. [Google Scholar] [CrossRef]
- Allsopp, T.E.; Robinson, M.; Wyatt, S.; Davies, A.M. TrkA mediates an NGF survival response in NGF-independent sensory neurons but not in parasympathetic neurons. Gene Ther. 1994, 1 (Suppl. S1), S59. [Google Scholar] [CrossRef]
- Arcamone, N.; Lucini, C.; Borzacchiello, G.; Castaldo, L.; Gargiulo, G.; De Girolamo, P. Distribution of NGF and NT-3-like protein immunoreactivity in the teleost kidney. Microsc. Res. Tech. 2005, 66, 17–24. [Google Scholar] [CrossRef]
- Russo, M.A.; Giustizieri, M.L.; Farini, D.; Siracusa, G. Expression of neurotrophin receptors in the developing and adult testis. Ital. J. Anat. Embryol. 1995, 100 (Suppl. S1), 543–551. [Google Scholar]
- Ji, A.; Shu, S.; Li, M.; Bao, X.; Zou, H.; Zhang, Z. Expression of recombinant rat Neurotrophin-3 in Chinese hamster ovary cells. Sci. China C Life Sci. 1999, 42, 655–662. [Google Scholar] [CrossRef]
- Iwane, M.; Watanabe, T.; Shintani, A.; Kaisho, Y.; Matsumoto, S.; Sasada, R.; Igarashi, K. Purification and characterization of biologically active recombinant human neurotrophin-3 produced by expression of a chimera gene in Chinese hamster ovary cells. Appl. Microbiol. Biotechnol. 1994, 41, 225–232. [Google Scholar] [CrossRef]
- Russo, M.A.; Giustizieri, M.L.; Farini, D.; Campagnolo, L.; De Felici, M.; Siracusa, G. Expression of the p75 neurotrophin receptor in the developing and adult testis of the rat. Int. J. Dev. Biol. 2001, 40 (Suppl. S1), 227S–228S. [Google Scholar]
- D’Angelo, L.; de Girolamo, P.; Cellerino, A.; Tozzini, E.T.; Castaldo, L.; Lucini, C. Neurotrophin Trk receptors in the brain of a teleost fish, Nothobranchius furzeri. Microsc. Res. Tech. 2012, 75, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Persson, H. NGF remove grower factors and neurotransmitters in the testis--new discoveries in reproduction biology. Lakartidningen 1990, 87, 4193–4196. [Google Scholar] [PubMed]
- Carrasco, R.; Singh, J.; Adams, G.P. The dynamics of trkA expression in the bovine ovary are associated with a luteotrophic effect of ovulation-inducing factor/nerve growth factor (OIF/NGF). Reprod. Biol. Endocrinol. 2016, 14, 47. [Google Scholar] [CrossRef] [PubMed]
- Julio-Pieper, M.; Lara, H.E.; Bravo, J.A.; Romero, C. Effects of nerve growth factor (NGF) on blood vessels area and expression of the angiogenic factors VEGF and TGFbeta1 in the rat ovary. Reprod. Biol. Endocrinol. 2006, 4, 57. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bao, L.; Li, Q.; Liu, Y.; Li, B.; Sheng, X.; Han, Y.; Weng, Q. Immunolocalization of NGF and its receptors in ovarian surface epithelium of the wild ground squirrel during the breeding and nonbreeding seasons. Eur. J. Histochem. 2014, 58, 2363. [Google Scholar] [CrossRef]
- Abir, R.; Fisch, B.; Jin, S.; Barnnet, M.; Ben-Haroush, A.; Felz, C.; Kessler-Icekson, G.; Feldberg, D.; Nitke, S.; Ao, A. Presence of NGF and its receptors in ovaries from human fetuses and adults. Mol. Hum. Reprod. 2005, 11, 229–236. [Google Scholar] [CrossRef]
- Maruccio, L.; Lucini, C.; de Girolamo, P.; Avallone, L.; Solcan, C.; Nechita, L.E.; Castaldo, L. Neurotrophins and Trk receptors in the developing and adult ovary of Coturnix coturnix japonica. Ann. Anat. 2018, 219, 35–43. [Google Scholar] [CrossRef]
- Schultz, R.; Metsis, M.; Hokfelt, T.; Parvinen, M.; Pelto-Huikko, M. Expression of neurotrophin receptors in rat testis. Upregulation of TrkA mRNA with hCG treatment. Mol. Cell Endocrinol. 2001, 182, 121–127. [Google Scholar] [CrossRef]
- Dees, W.L.; Hiney, J.K.; Schultea, T.D.; Mayerhofer, A.; Danilchik, M.; Dissen, G.A.; Ojeda, S.R. The primate ovary contains a population of catecholaminergic neuron-like cells expressing nerve growth factor receptors. Endocrinology 1995, 136, 5760–5768. [Google Scholar] [CrossRef]
- Zhu, G.; Fang, C.; Mo, C.; Wang, Y.; Huang, Y.; Li, J. Transcriptomic analysis of granulosa cell populations proximal and distal to the germinal disc of chicken preovulatory follicles. Sci. Rep. 2021, 11, 4683. [Google Scholar] [CrossRef] [PubMed]
- Dorfman, M.; Arancibia, S.; Fiedler, J.L.; Lara, H.E. Chronic intermittent cold stress activates ovarian sympathetic nerves and modifies ovarian follicular development in the rat. Biol. Reprod. 2003, 68, 2038–2043. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y. Effect of nerve growth factor (NGF) on the development of preimplantation rabbit embryos in vitro. Vet. Res. Commun. 2010, 34, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Nebreda, A.R.; Martin-Zanca, D.; Kaplan, D.R.; Parada, L.F.; Santos, E. Induction by NGF of meiotic maturation of Xenopus oocytes expressing the trk proto-oncogene product. Science 1991, 252, 558–561. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Y.; Zhang, J.; Wang, L.; Li, Q.; Sheng, X.; Han, Y.; Yuan, Z.; Weng, Q. Testicular expression of NGF, TrkA and p75 during seasonal spermatogenesis of the wild ground squirrel (Citellus dauricus Brandt). Eur. J. Histochem. 2015, 59, 2522. [Google Scholar] [CrossRef]
- Schachter, M.; Peret, M.W.; Moriwaki, C.; Wheeler, G.D.; Matthews, R.W.; Mehta, J.G.; Labedz, T. The varied localization and functional significance of kallikrein-like enzymes in salivary glands, pancreas, colon, sex glands and spermatozoa, including evidence for the presence of nerve growth factor (NGF) in bull sperm acrosome. Adv. Exp. Med. Biol. 1986, 198 Pt A, 1–10. [Google Scholar]
- Li, C.; Sun, Y.; Yi, K.; Ma, Y.; Sun, Y.; Zhang, W.; Zhou, X. Detection of nerve growth factor (NGF) and its specific receptor (TrkA) in ejaculated bovine sperm, and the effects of NGF on sperm function. Theriogenology 2010, 74, 1615–1622. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, A.; Arias-Alvarez, M.; Timon, P.; Bautista, J.M.; Rebollar, P.G.; Lorenzo, P.L.; Garcia-Garcia, R.M. Characterization of beta-Nerve Growth Factor-TrkA system in male reproductive tract of rabbit and the relationship between beta-NGF and testosterone levels with seminal quality during sexual maturation. Theriogenology 2019, 126, 206–213. [Google Scholar] [CrossRef]
- Castellini, C.; Mattioli, S.; Dal Bosco, A.; Mancinelli, A.C.; Rende, M.; Stabile, A.M.; Pistilli, A. Role of NGF on sperm traits: A review. Theriogenology 2020, 150, 210–214. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, H.; Yang, Y.; Liu, H.; Zhang, Q.; Xiang, Q.; Ge, R.; Su, Z.; Huang, Y. NGF induces adult stem Leydig cells to proliferate and differentiate during Leydig cell regeneration. Biochem. Biophys. Res. Commun. 2013, 436, 300–305. [Google Scholar] [CrossRef]
- Castellini, C.; Mattioli, S.; Cotozzolo, E.; Pistilli, A.; Rende, M.; Bartolini, D.; Di Sante, G.; Menchetti, L.; Dal Bosco, A.; Stabile, A.M. The Effect of Interaction NGF/p75(NTR) in Sperm Cells: A Rabbit Model. Cells 2022, 11, 1035. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Yang, Y.; Ji, X.; He, W.; Fan, J.; Huang, Y.; Wang, Y. NGF Rescues Spermatogenesis in Azoospermic Mice. Reprod. Sci. 2021, 28, 2780–2788. [Google Scholar] [CrossRef] [PubMed]
- Confino, S.; Dor, T.; Tovin, A.; Wexler, Y.; Ben-Moshe Livne, Z.; Kolker, M.; Pisanty, O.; Park, S.K.; Geyer, N.; Reiter, J.; et al. A Zebrafish Model for a Rare Genetic Disease Reveals a Conserved Role for FBXL3 in the Circadian Clock System. Int. J. Mol. Sci. 2022, 23, 2373. [Google Scholar] [CrossRef] [PubMed]
- Bowley, G.; Kugler, E.; Wilkinson, R.; Lawrie, A.; van Eeden, F.; Chico, T.J.A.; Evans, P.C.; Noel, E.S.; Serbanovic-Canic, J. Zebrafish as a tractable model of human cardiovascular disease. Br. J. Pharmacol. 2022, 179, 900–917. [Google Scholar] [CrossRef] [PubMed]
- Di Franco, G.; Usai, A.; Funel, N.; Palmeri, M.; Montesanti, I.E.R.; Bianchini, M.; Gianardi, D.; Furbetta, N.; Guadagni, S.; Vasile, E.; et al. Use of zebrafish embryos as avatar of patients with pancreatic cancer: A new xenotransplantation model towards personalized medicine. World J. Gastroenterol. 2020, 26, 2792–2809. [Google Scholar] [CrossRef] [PubMed]
- Cacialli, P.; D’Angelo, L.; Kah, O.; Coumailleau, P.; Gueguen, M.M.; Pellegrini, E.; Lucini, C. Neuronal expression of brain derived neurotrophic factor in the injured telencephalon of adult zebrafish. J. Comp. Neurol. 2018, 526, 569–582. [Google Scholar] [CrossRef]
- Menke, A.L.; Spitsbergen, J.M.; Wolterbeek, A.P.; Woutersen, R.A. Normal anatomy and histology of the adult zebrafish. Toxicol. Pathol. 2011, 39, 759–775. [Google Scholar] [CrossRef]
- Cacialli, P.; D’Angelo, L.; de Girolamo, P.; Avallone, L.; Lucini, C.; Pellegrini, E.; Castaldo, L. Morpho-Functional Features of the Gonads of Danio rerio: The Role of Brain-Derived Neurotrophic Factor. Anat Rec. 2018, 301, 140–147. [Google Scholar] [CrossRef]
- Cacialli, P.; Mahony, C.B.; Petzold, T.; Bordignon, P.; Rougemont, A.L.; Bertrand, J.Y. A connexin/ifi30 pathway bridges HSCs with their niche to dampen oxidative stress. Nat. Commun. 2021, 12, 4484. [Google Scholar] [CrossRef]
- Mahony, C.B.; Cacialli, P.; Pasche, C.; Monteiro, R.; Savvides, S.N.; Bertrand, J.Y. Hapln1b, a central organizer of the ECM, modulates kit signaling to control developmental hematopoiesis in zebrafish. Blood Adv. 2021, 5, 4935–4948. [Google Scholar] [CrossRef]
- Russo, B.; Borowczyk, J.; Cacialli, P.; Moguelet, P.; Truchetet, M.E.; Modarressi, A.; Brembilla, N.C.; Bertrand, J.; Boehncke, W.H.; Chizzolini, C. IL-25 participates in keratinocyte-driven dermal matrix turnover and is reduced in Systemic Sclerosis epidermis. Rheumatology 2022, keac044. [Google Scholar] [CrossRef] [PubMed]
- Perrard, M.H.; Chassaing, E.; Montillet, G.; Sabido, O.; Durand, P. Cytostatic factor proteins are present in male meiotic cells and beta-nerve growth factor increases mos levels in rat late spermatocytes. PLoS ONE 2009, 4, e7237. [Google Scholar] [CrossRef] [PubMed]
- Sari, L.M.; Zampini, R.; Arganaraz, M.E.; Carretero, M.I.; Fumuso, F.G.; Barraza, D.E.; Ratto, M.; Apichela, S.A. Expression of beta-NGF and high-affinity NGF receptor (TrKA) in llama (Lama glama) male reproductive tract and spermatozoa. Mol. Reprod. Dev. 2018, 85, 934–944. [Google Scholar] [CrossRef] [PubMed]
- Perrard, M.H.; Durand, P. Redundancy of the effect of TGFbeta1 and beta-NGF on the second meiotic division of rat spermatocytes. Microsc. Res. Tech. 2009, 72, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Li, C.J.; Jiang, Y.W.; Chen, S.X.; Li, H.J.; Chen, L.; Liu, Y.T.; Gao, S.; Zhao, Y.; Zhu, X.L.; Wang, H.T.; et al. 4-Methylcatechol inhibits cell growth and testosterone production in TM3 Leydig cells by reducing mitochondrial activity. Andrologia 2017, 49, e12581. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.J.; Liu, Y.H.; Huang, S.Y.; Zang, Z.J. Insights into the Regulation on Proliferation and Differentiation of Stem Leydig Cells. Stem Cell Rev. Rep. 2021, 17, 1521–1533. [Google Scholar] [CrossRef]
- Silva, M.; Paiva, L.; Ratto, M.H. Ovulation mechanism in South American Camelids: The active role of beta-NGF as the chemical signal eliciting ovulation in llamas and alpacas. Theriogenology 2020, 150, 280–287. [Google Scholar] [CrossRef]
- Saeednia, S.; Bahadoran, H.; Amidi, F.; Asadi, M.H.; Naji, M.; Fallahi, P.; Nejad, N.A. Nerve growth factor in human semen: Effect of nerve growth factor on the normozoospermic men during cryopreservation process. Iran J. Basic Med. Sci. 2015, 18, 292–299. [Google Scholar]
- Saeednia, S.; Shabani Nashtaei, M.; Bahadoran, H.; Aleyasin, A.; Amidi, F. Effect of nerve growth factor on sperm quality in asthenozoosprmic men during cryopreservation. Reprod. Biol. Endocrinol. 2016, 14, 29. [Google Scholar] [CrossRef]
- Li, C.; Zheng, L.; Wang, C.; Zhou, X. Absence of nerve growth factor and comparison of tyrosine kinase receptor A levels in mature spermatozoa from oligoasthenozoospermic, asthenozoospermic and fertile men. Clin. Chim. Acta 2010, 411, 1482–1486. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, A.; Abad, P.; Arias-Alvarez, M.; Rebollar, P.G.; Bautista, J.M.; Lorenzo, P.L.; Garcia-Garcia, R.M. Recombinant rabbit beta nerve growth factor production and its biological effects on sperm and ovulation in rabbits. PLoS ONE 2019, 14, e0219780. [Google Scholar]
- Garrido, M.P.; Vallejos, C.; Girardi, S.; Gabler, F.; Selman, A.; Lopez, F.; Vega, M.; Romero, C. NGF/TRKA Promotes ADAM17-Dependent Cleavage of P75 in Ovarian Cells: Elucidating a Pro-Tumoral Mechanism. Int. J. Mol. Sci. 2022, 23, 2124. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, W.; Du, J.; Yu, Y.; Liang, N.; Liang, M.; Yao, G.; Cui, S.; Huang, H.; Sun, F. NGF promotes mouse granulosa cell proliferation by inhibiting ESR2 mediated down-regulation of CDKN1A. Mol. Cell Endocrinol. 2015, 406, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Sheng, X.; Bao, L.; Huang, S.; Li, Q.; Liu, Y.; Han, Y.; Watanabe, G.; Taya, K.; Weng, Q. Seasonal changes in expression of nerve growth factor and its receptors TrkA and p75 in the ovary of wild ground squirrel (Citellus dauricus Brandt). J. Ovarian Res. 2014, 7, 3. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Salas, C.; Julio-Pieper, M.; Valladares, M.; Pommer, R.; Vega, M.; Mastronardi, C.; Kerr, B.; Ojeda, S.R.; Lara, H.E.; Romero, C. Nerve growth factor-dependent activation of trkA receptors in the human ovary results in synthesis of follicle-stimulating hormone receptors and estrogen secretion. J. Clin. Endocrinol. Metab. 2006, 91, 2396–2403. [Google Scholar] [CrossRef]
- Valderrama, X.; Ulloa-Leal, C.; Silva, M.E.; Goicochea, J.; Apichela, S.; Arganaraz, M.; Sari, L.; Paiva, L.; Ratto, V.F.; Ratto, M.H. beta-NGF Stimulates Steroidogenic Enzyme and VEGFA Gene Expression, and Progesterone Secretion via ERK 1/2 Pathway in Primary Culture of Llama Granulosa Cells. Front. Vet. Sci. 2020, 7, 586265. [Google Scholar] [CrossRef]
- Levanti, M.B.; Germana, A.; Abbate, F.; Montalbano, G.; Vega, J.A.; Germana, G. TrkA and p75NTR in the ovary of adult cow and pig. J. Anat. 2005, 207, 93–96. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cacialli, P. Expression of Nerve Growth Factor and Its Receptor TrkA in the Reproductive System of Adult Zebrafish. Vet. Sci. 2022, 9, 225. https://doi.org/10.3390/vetsci9050225
Cacialli P. Expression of Nerve Growth Factor and Its Receptor TrkA in the Reproductive System of Adult Zebrafish. Veterinary Sciences. 2022; 9(5):225. https://doi.org/10.3390/vetsci9050225
Chicago/Turabian StyleCacialli, Pietro. 2022. "Expression of Nerve Growth Factor and Its Receptor TrkA in the Reproductive System of Adult Zebrafish" Veterinary Sciences 9, no. 5: 225. https://doi.org/10.3390/vetsci9050225
APA StyleCacialli, P. (2022). Expression of Nerve Growth Factor and Its Receptor TrkA in the Reproductive System of Adult Zebrafish. Veterinary Sciences, 9(5), 225. https://doi.org/10.3390/vetsci9050225