
Genetic Analysis of Torque Teno Canis Virus Identified in Republic of Korea
 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Viral DNA Extraction
2.3. TTCaV Detection
2.4. PCR Amplification of Entire TTCaV Genome
2.5. Phylogenetic Tree and Sequence Identity Analysis
2.6. Similarity Plot and Recombination Analysis
3. Results
3.1. TTCaV Incidence in Dogs
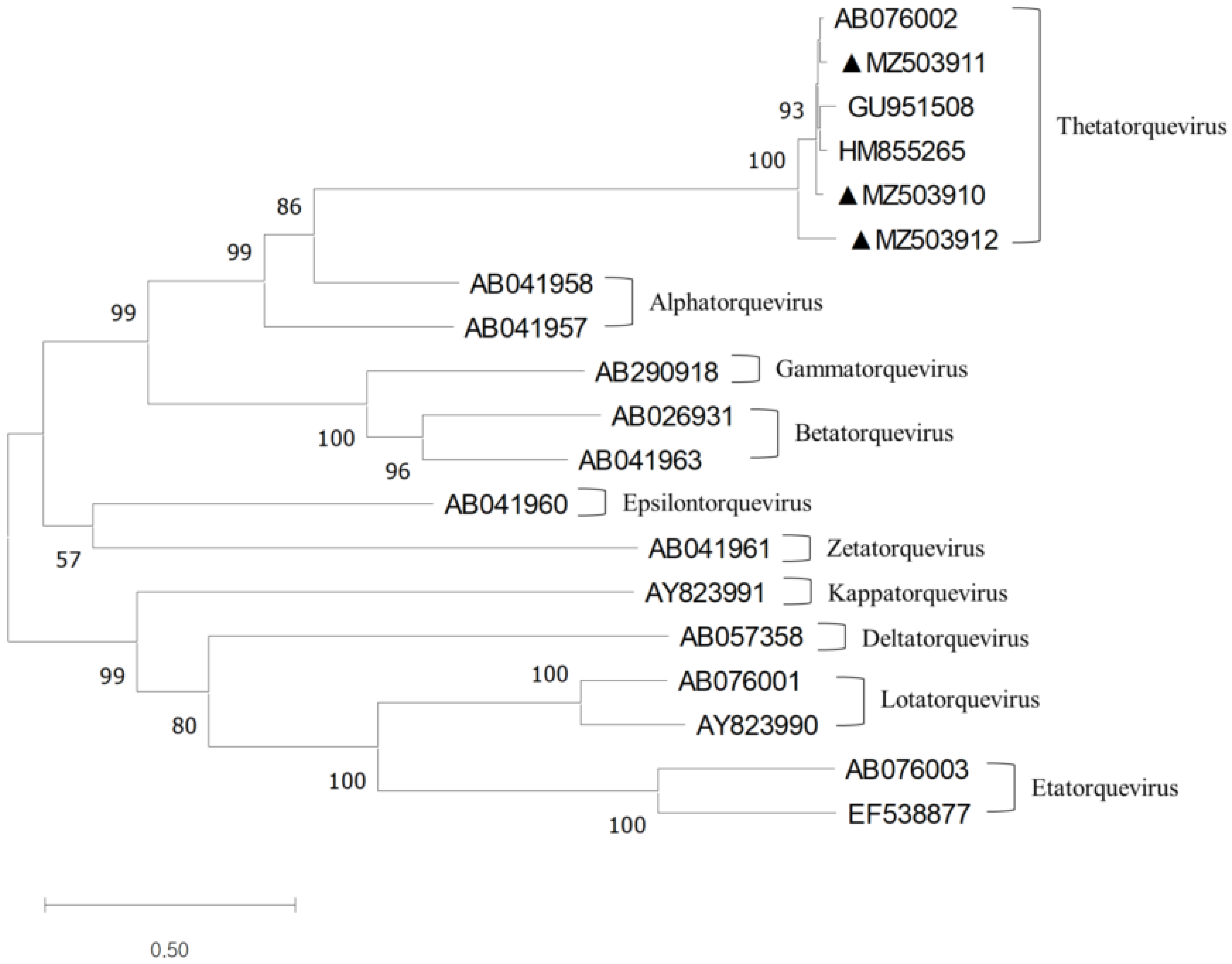
3.2. Complete Genomic Analysis of TTCaV
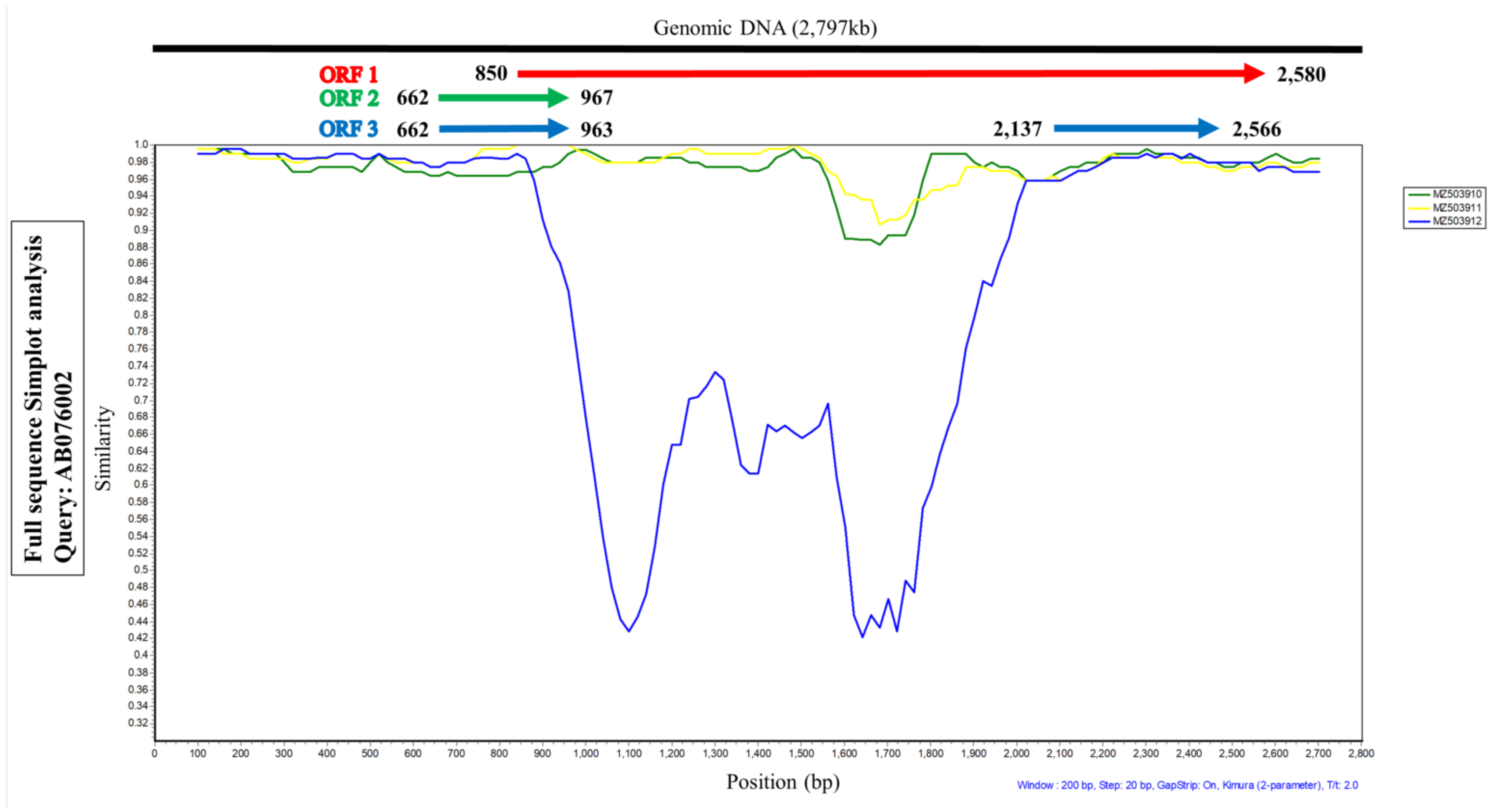
3.3. SimPlot and Recombination Analysis of TTCaV
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reshetnyak, V.I.; Maev, I.V.; Burmistrov, A.I.; Chekmazov, I.A.; Karlovich, T.I. Torque Teno Virus in Liver Diseases: On the Way towards Unity of View. World J. Gastroenterol. 2020, 26, 1691. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H. TT Viruses in Animals. Curr. Top Microbiol. Immunol. 2009, 331, 35–52. [Google Scholar] [CrossRef] [PubMed]
- Varsani, A.; Opriessnig, T.; Celer, V.; Maggi, F.; Okamoto, H.; Blomström, A.-L.; Cadar, D.; Harrach, B.; Biagini, P.; Kraberger, S. Taxonomic Update for Mammalian Anelloviruses (Family Anelloviridae). Arch. Virol. 2021, 166, 2943–2953. [Google Scholar] [CrossRef] [PubMed]
- Saback, F.L.; Gomes, S.A.; de Paula, V.S.; da Silva, R.R.S.; Lewis-Ximenez, L.L.; Niel, C. Age-Specific Prevalence and Transmission of TT Virus. J. Med. Virol. 1999, 59, 318–322. [Google Scholar] [CrossRef]
- Peng, Y.H.; Nishizawa, T.; Takahashi, M.; Ishikawa, T.; Yoshikawa, A.; Okamoto, H. Analysis of the Entire Genomes of Thirteen TT Virus Variants Classifiable into the Fourth and Fifth Genetic Groups, Isolated from Viremic Infants. Arch. Virol. 2002, 147, 21–41. [Google Scholar] [CrossRef] [PubMed]
- Muljono, D.H.; Nishizawa, T.; Tsuda, F.; Takahashi, M.; Okamoto, H. Molecular Epidemiology of TT Virus (TTV) and Characterization of Two Novel TTV Genotypes in Indonesia. Arch. Virol. 2001, 146, 1249–1266. [Google Scholar] [CrossRef]
- Tanaka, Y.; Mizokami, M.; Orito, E.; Nakano, T.; Kato, T.; Ding, X.; Ohno, T.; Ueda, R.; Sonoda, S.; Tajima, K.; et al. A New Genotype of TT Virus (TTV) Infection Among Colombian Native Indians. J. Med. Virol. 1999, 57, 264–268. [Google Scholar] [CrossRef]
- Singh Mankotia, D.; Irshad, M. Cloning and Expression of N22 Region of Torque Teno Virus (TTV) Genome and Use of Peptide in Developing Immunoassay for TTV Antibodies. Virol. J. 2014, 11, 96. [Google Scholar] [CrossRef]
- Okamoto, H. Molecular Cloning and Characterization of a Novel DNA Virus (TTV) Associated with Posttransfusion Hepatitis of Unknown Etiology. Hepatol. Res. 1998, 10, 1–16. [Google Scholar] [CrossRef]
- Rezahosseini, O.; Drabe, C.H.; Sørensen, S.S.; Rasmussen, A.; Perch, M.; Ostrowski, S.R.; Nielsen, S.D. Torque-Teno Virus Viral Load as a Potential Endogenous Marker of Immune Function in Solid Organ Transplantation. Transpl. Rev. 2019, 33, 137–144. [Google Scholar] [CrossRef]
- Bendinelli, M.; Pistello, M.; Maggi, F.; Fornai, C.; Freer, G.; Vatteroni, M.L. Molecular Properties, Biology, and Clinical Implications of TT Virus, a Recently Identified Widespread Infectious Agent of Humans. Clin. Microbiol. Rev. 2001, 14, 98–113. [Google Scholar] [CrossRef] [PubMed]
- Spandole, S.; Cimponeriu, D.; Berca, L.M.; Mihăescu, G. Human Anelloviruses: An Update of Molecular, Epidemiological and Clinical Aspects. Arch. Virol. 2015, 160, 893–908. [Google Scholar] [CrossRef]
- Deng, X.; Terunuma, H.; Handema, R.; Sakamoto, M.; Kitamura, T.; Ito, M.; Akahane, Y. Higher Prevalence and Viral Load of TT Virus in Saliva Than in the Corresponding Serum: Another Possible Transmission Route and Replication Site of TT Virus. J. Med. Virol. 2000, 62, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H.; Nishizawa, T.; Tawara, A.; Peng, Y.; Takahashi, M.; Kishimoto, J.; Tanaka, T.; Miyakawa, Y.; Mayumi, M. Species-Specific TT Viruses in Humans and Nonhuman Primates and Their Phylogenetic Relatedness. Virology 2000, 277, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H.; Nishizawa, T.; Takahashi, M.; Tawara, A.; Peng, Y.; Kishimoto, J.; Wang, Y. Genomic and Evolutionary Characterization of TT Virus (TTV) in Tupaias and Comparison with Species-Specific TTVs in Humans and Non-Human Primates. J. Gen. Virol. 2001, 82, 2041–2050. [Google Scholar] [CrossRef]
- Okamoto, H.; Takahashi, M.; Nishizawa, T.; Tawara, A.; Fukai, K.; Muramatsu, U.; Naito, Y.; Yoshikawa, A. Genomic Characterization of TT Viruses (TTVs) in Pigs, Cats and Dogs and Their Relatedness with Species-Specific TTVs in Primates and Tupaias. J. Gen. Virol. 2002, 83, 1291–1297. [Google Scholar] [CrossRef]
- Brassard, J.; Gagné, M.J.; Lamoureux, L.; Inglis, G.D.; Leblanc, D.; Houde, A. Molecular Detection of Bovine and Porcine Torque Teno Virus in Plasma and Feces. Vet. Microbiol. 2008, 126, 271–276. [Google Scholar] [CrossRef]
- Sun, W.; Xie, C.; Liang, C.; Zheng, M.; Zhao, G.; Zhang, P.; Han, J.; Jing, J.; Wen, S.; Xiao, P.; et al. Molecular Detection and Genomic Characterization of Torque Teno Canis Virus in Domestic Dogs in Guangxi Province, China. J. Biotechnol. 2017, 252, 50–54. [Google Scholar] [CrossRef]
- Hino, S.; Miyata, H. Torque Teno Virus (TTV): Current Status. Rev. Med. Virol. 2007, 17, 45–57. [Google Scholar] [CrossRef]
- Manzin, A.; Mallus, F.; Macera, L.; Maggi, F.; Blois, S. Global Impact of Torque Teno Virus Infection in Wild and Domesticated Animals. J. Infect. Dev. Ctries 2015, 9, 562–570. [Google Scholar] [CrossRef]
- Işidan, H.; Turan, T.; Atasoy, M.O. Molecular Detection and Genomic Characterisation of Torque Teno Canis Virus in Turkey Türkiye’nin Doğu ve Güneydoğu Anadolu Bölgelerinde Hayvan Kuduzunun Epidemiyolojisi View Project. Vet. Arhiv. 2020, 90, 467–475. [Google Scholar] [CrossRef]
- Lan, D.; Hua, X.; Cui, L.; Luo, X.; Liu, Z.; San, T.; Zhu, C.X.; Zhao, W.; Yang, Z. Sequence Analysis of a Torque Teno Canis Virus Isolated in China. Virus Res. 2011, 160, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Hillis, D.M.; Bull, J.J. An Empirical Test of Bootstrapping as a Method for Assessing Confidence in Phylogenetic Analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Worobey, M. Extensive Homologous Recombination among Widely Divergent TT Viruses. J. Virol. 2000, 74, 7666–7670. [Google Scholar] [CrossRef] [PubMed]
- Manni, F.; Rotola, A.; Caselli, E.; Bertorelle, G.; di Luca, D. Detecting Recombination in TT Virus: A Phylogenetic Approach. J. Mol. Evol. 2002, 55, 563–572. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Primer Sets | Primer Names | Sequences (5′→3′) | Positions * | Amplicon Length (bp) |
|---|---|---|---|---|
| I | TTCaV-F1 | CGCCATCTTGGATTGGAAATC | 169–189 993–1015 | 847 |
| TTCaV-R1 | TAGAAATGTATTGTCTTTTGGTG | |||
| II | ORF1-F | AAGCAGCACTGTAGCTGGAG | 770–789 2613–2634 | 1865 |
| ORF1-R | CTTACGTCACAAAACAAGATGG | |||
| III | TTCaV-F2 | ATGGTGGCCCATTACCAACCCCTAC | 1911–1935 247–273 | 1160 |
| TTCaV-R2 | TATTCCGATGTCCGATTTGCATAATCG |
| Region | Number of Fecal Samples | Number of TTCaV-Positive Fecal Samples | Incidence (%) |
|---|---|---|---|
| Yangpyeong | 74 | 4 | 5 |
| Anseong | 29 | 7 | 24 |
| Seoul | 32 | 2 | 6 |
| Total | 135 | 13 | 9.6 |
| Accession Number | Genome Length (nt) | ORF1 aa | ORF2 aa | ORF3 aa |
|---|---|---|---|---|
| AB076002 | 2797 | 576 | 101 | 243 |
| MZ503910 | 2793 | 576 | 101 | 243 |
| MZ503911 | 2795 | 577 | 101 | 243 |
| MZ503912 | 2791 | 575 | 101 | 243 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.-Y.; Ahn, H.-S.; Han, S.-H.; Go, H.-J.; Kim, D.-H.; Kim, J.-H.; Lee, J.-B.; Park, S.-Y.; Song, C.-S.; Lee, S.-W.; et al. Genetic Analysis of Torque Teno Canis Virus Identified in Republic of Korea. Vet. Sci. 2022, 9, 693. https://doi.org/10.3390/vetsci9120693
Kim D-Y, Ahn H-S, Han S-H, Go H-J, Kim D-H, Kim J-H, Lee J-B, Park S-Y, Song C-S, Lee S-W, et al. Genetic Analysis of Torque Teno Canis Virus Identified in Republic of Korea. Veterinary Sciences. 2022; 9(12):693. https://doi.org/10.3390/vetsci9120693
Chicago/Turabian StyleKim, Da-Yoon, Hee-Seop Ahn, Sang-Hoon Han, Hyeon-Jeong Go, Dong-Hwi Kim, Jae-Hyeong Kim, Joong-Bok Lee, Seung-Yong Park, Chang-Seon Song, Sang-Won Lee, and et al. 2022. "Genetic Analysis of Torque Teno Canis Virus Identified in Republic of Korea" Veterinary Sciences 9, no. 12: 693. https://doi.org/10.3390/vetsci9120693
APA StyleKim, D.-Y., Ahn, H.-S., Han, S.-H., Go, H.-J., Kim, D.-H., Kim, J.-H., Lee, J.-B., Park, S.-Y., Song, C.-S., Lee, S.-W., & Choi, I.-S. (2022). Genetic Analysis of Torque Teno Canis Virus Identified in Republic of Korea. Veterinary Sciences, 9(12), 693. https://doi.org/10.3390/vetsci9120693