
Effect of Breeding Season on Haemosporidian Infections in Domestic Chickens

 and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
Statistical Analysis
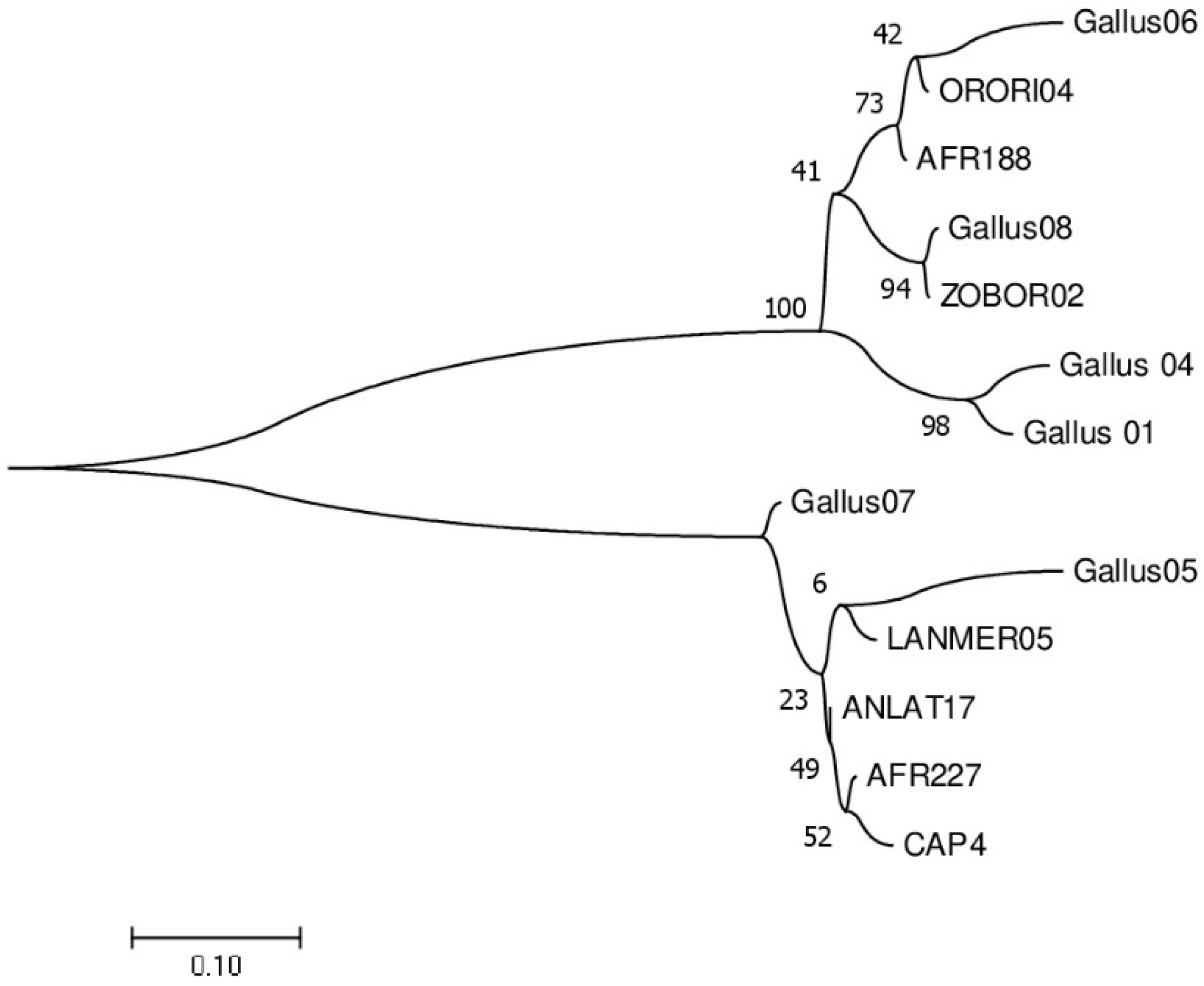
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shaw, D.J.; Grenfell, B.T.; Dobson, A.P. Patterns of macroparasite aggregation in wildlife host populations. Parasitology 1998, 117, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Huijben, S.; Chan, B.H.K.; Nelson, W.A.; Read, A.F. The impact of within-host ecology on the fitness of a drug-resistant parasite. Evol. Med. Public Health 2018, 6, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Loiseau, C.; Harrigan, R.J.; Cornel, A.J.; Guers, S.L.; Dodge, M.; Marzec, T.; Carlson, J.S.; Seppi, B.; Sehgal, R.N.M. First evidence and predictions of plasmodium transmission in Alaskan bird populations. PLoS ONE 2012, 7, e44729. [Google Scholar] [CrossRef]
- Ricklefs, R.E.; Swanson, B.L.; Fallon, S.M. Community relationships of avian malaria parasites in southern Missouri. Ecol Monogr. 2005, 75, 543–559. [Google Scholar] [CrossRef]
- Wegge, P.; Rolstad, J. Climate change and bird reproduction: Warmer springs benefit breeding success in boreal forest grouse. Proc. R. Soc. B Bio Sci. 2017, 284, 20171528. [Google Scholar] [CrossRef] [PubMed]
- Waller, E.K.; Crimmins, T.M.; Walker, J.J.; Posthumus, E.E.; Weltzin, J.F. Differential changes in the onset of spring across US national wildlife refuges and north American migratory bird flyways. PLoS ONE 2018, 13, e0202495. [Google Scholar] [CrossRef]
- Segner, H.; Verburg-van, K.; Lidy, B.M.; Magdalena, C. The immunomodulatory role of the hypothalamus-pituitary-gonad axis: Proximate mechanism for reproduction-immune tradeoffs? Dev. Comp. Immunol. 2017, 66, 43–60. [Google Scholar] [CrossRef]
- Casto, J.M.; Nolan, V., Jr.; Ketterson, E.D. Steroid hormones and immune function: Experimental studies in wild and captive dark-eyed juncos (Junco hyemalis). Am. Nat. 2001, 157, 408–420. [Google Scholar] [CrossRef]
- Singh, S.S.; Haldar, C. Melatonin prevents testosterone-induced suppression of immune parameters and splenocyte proliferation in Indian tropical jungle bush quail, Perdicula asiatica. Gen. Comp. Endocrinol. 2005, 141, 226–232. [Google Scholar] [CrossRef]
- Deviche, P.; Parris, J.; Hobson, K.A. Testosterone treatment to free ranging male dark-eyed juncos (Junco hyemalis) exacerbates hemoparasitic infection. Auk 2006, 123, 548–562. [Google Scholar] [CrossRef]
- Romero, L.M. Seasonal changes in plasma glucocorticoid concentrations in free-living vertebrates. Gen. Comp. Endocrinol. 2002, 128, 1–24. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Wingfield, J.C. The concept of allostasis in biology and biomedicine. Horm. Behav. 2003, 43, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Landys, M.M.; Ramenofsky, M.; Wingfield, J.C. Actions of glucocorticoids at a seasonal baseline as compared to stress-related levels in the regulation of periodic life processes. Gen. Comp. Endocrinol. 2006, 148, 132–149. [Google Scholar] [CrossRef]
- Atkinson, C.; van Riper, C. Pathogenicity and epizootiology of avian haematozoa: Plasmodium, Leucocytozoon, and Haemoproteus. In Bird-Parasite Interactions; Loye, J., Zuk, M., Eds.; Oxford University Press: London, UK, 1991; pp. 19–48. [Google Scholar]
- Norris, K.; Anwar, M.; Read, A.F. Reproductive effort influences the prevalence of haematozoan parasites in great tits. J. Anim. Ecol. 1994, 63, 601–610. [Google Scholar] [CrossRef]
- Allander, K. Reproductive investment and parasite susceptibility in the great tit. Funct. Ecol. 1997, 11, 358–364. [Google Scholar] [CrossRef]
- Stearns, S.C. The Evolution of Life Histories; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Christe, P.; Arlettaz, R.; Vogel, P. Variation in intensity of a parasitic mite (Spinturnix myoti) in relation to the reproductive cycle and immunocompetence of its bat host (Myotis myotis). Ecol. Lett. 2000, 3, 207–212. [Google Scholar] [CrossRef]
- Neuhaus, P. Parasite removal and its impact on litter size and body condition in Columbian ground squirrels (Spermophilus columbianus). Proc. R. Soc. B Biol. Sci. 2003, 270, 213–215. [Google Scholar] [CrossRef]
- Hillegass, M.A.; Waterman, J.M.; Roth, J.D. Parasite removal increases reproductive success in a social African ground squirrel. Behav. Ecol. 2010, 21, 696–700. [Google Scholar] [CrossRef]
- Schwanz, L.E. Chronic parasitic infection alters reproductive output in deer mice. Behav. Ecol. Soc. 2008, 62, 1351–1358. [Google Scholar] [CrossRef]
- Mougeot, F.; Pérez-Rodríguez, L.; Martínez-Padilla, J.; Leckie, F.; Redpath, S.M. Parasites, testosterone and honest carotenoid-based signalling of health. Funct. Ecol. 2004, 21, 886–898. [Google Scholar] [CrossRef]
- Pelletier, F.; Page, K.A.; Ostiguy, T.; Festa-Bianchet, M. Fecal counts of lungworm larvae and reproductive effort in bighorn sheep, Ovis canadensis. Oikos 2005, 110, 473–480. [Google Scholar] [CrossRef]
- Viljoen, H.; Bennett, N.C.; Ueckermann, E.A.; Lutermann, H. The role of host traits, season and group size on parasite burdens in a cooperative mamma. PLoS ONE 2011, 6, e27003. [Google Scholar] [CrossRef]
- Cattadori, I.M.; Boag, B.; Bjornstad, O.N.; Cornell, S.J.; Hudson, P.J. Peak shift and epidemiology in a seasonal host-nematode system. Proc. R. Soc. B Biol. Sci. 2005, 272, 1163–1169. [Google Scholar] [CrossRef] [PubMed]
- Cornell, S.J.; Bjornstad, O.N.; Cattadori, I.M.; Boag, B.; Hudson, P.J. Seasonality, cohort-dependence and the development of immunity in a natural host-nematode system. Proc. R.Soc. B Biol. Sci. 2008, 275, 511–518. [Google Scholar] [CrossRef]
- Lourenco, S.I.; Palmeirin, J.M. Which factors regulate the reproduction of ectoparasites of temperate-zone cave-dwelling bats? Para. Res. 2008, 104, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Brockmann, J.H.; St Mary, C.M.; Ponciano, J.M. Discovering structural complexity and its causes: Breeding aggregations in horseshoe crabs. Anim. Behav. 2018, 143, 177–191. [Google Scholar] [CrossRef]
- Campbell, L.A.D.; Tkaczynski, P.J.; Lehmann, J.; Mouna, M.; Majolo, B. Social thermoregulation as a potential mechanism linking sociality and fitness: Barbary macaques with more social partners form larger huddles. Sci. Rep. 2018, 8, 6074. [Google Scholar] [CrossRef]
- Pulgarín-R, P.C.; Cadena, C.D.; Gómez, J.P.; Robinson, S.; Ricklefs, R.E. Host species, and not environment, predicts variation in blood parasite prevalence, distribution, and diversity along a humidity gradient in northern South America. Ecol. Evol. 2018, 8, 3800–3814. [Google Scholar] [CrossRef]
- Garamszegi, L.Z. Climate change increases the risk of malaria in birds. Glob. Chang. Biol. 2010, 17, 1751–1759. [Google Scholar] [CrossRef]
- Schmid-Hempel, P. Parasites and their social hosts. Trends Parasitol. 2017, 33, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.A.; McBrayer, L.D.; Herrel, A. Bite force in vertebrates: Opportunities and caveats for use of a nonpareil whole-animal performance measure. Biol. J. Linn. Soc. Lond. 2008, 93, 709–720. [Google Scholar] [CrossRef]
- Hellgren, O.; Waldenström, J.; Bensch, S. A new PCR assay for simultaneous studies of Leucocytozoon, Plasmodium, and Haemoproteus from avian blood. J. Parasitol. 2004, 90, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Thrusfield, M. Veterinary Epidemiology, 2nd ed.; Blackwell Science: Oxford, UK, 1995; pp. 182–183. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Bensch, S.; Hellgren, O.; Pérez-Tris, J. MalAvi: A public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Mol. Ecol. Res. 2009, 9, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Lutz, H.L.; Hochachka, W.M.; Engel, J.I.; Bell, J.A.; Tkach, V.V.; Bates, J.M.; Hackett, S.J.; Weckstein, J.D. Parasite prevalence corresponds to host life history in a diverse assemblage of Afrotropical birds and haemosporidian parasites. PLoS ONE 2015, 10, e0121254. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Whittingham, M.J.; Stephens, P.A.; Bradbury, R.B.; Freckleton, R.P. Why do we still use stepwise modelling in ecology and behaviour? J. Anim. Ecol. 2006, 75, 1182–1189. [Google Scholar] [CrossRef]
- Murtaugh, P.A. Performance of several variable-selection methods applied to real ecological data. Ecol. Lett. 2009, 12, 1061–1068. [Google Scholar] [CrossRef]
- Deviche, P.; Greiner, E.C.; Manteca, X. Interspecific variability of prevalence in blood parasites of adult passerine birds during the breeding season in Alaska. J. Wildl. Dis. 2001, 37, 28–35. [Google Scholar] [CrossRef]
- Szöllősi, E.; Garamszegi, L.Z.; Hegyi, G.; Laczi, M.; Rosivall, B.; Török, J. Haemoproteus infection status of collared flycatcher males’ changes within a breeding season. Parasitol. Res. 2016, 115, 4663–4672. [Google Scholar] [CrossRef]
- Ferraguti, M.; Martínez-de la Puente, J.; Muñoz, J.; Roiz, D.; Ruiz, S.; Soriguer, R.; Figuerola, J. Avian Plasmodium in Culex and Ochlerotatus mosquitoes from southern Spain: Effects of season and host-feeding source on parasite dynamics. PLoS ONE 2013, 8, e66237. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, C.L.; Wood, M.J.; Day, K.P.; Sheldon, B.C. Seasonal variation in Plasmodium prevalence in a population of blue tits Cyanistes caeruleus. J. Anim. Ecol. 2008, 77, 540–548. [Google Scholar] [CrossRef]
- Santiago-Alarcon, D.; Bloch, R.; Rolshausen, G.; Schaefer, H.M.; Segelbacher, G. Prevalence, diversity, and interaction patterns of avian haemosporidians in a four-year study of blackcaps in a migratory divide. Parasitology 2011, 138, 824–835. [Google Scholar] [CrossRef]
- Cornelius, J.M.; Zylberberg, M.; Breuner, C.C.; Gleiss, A.C.; Hahn, T.P. Assessing the role of reproduction and stress in the spring emergence of haematozoan parasites in birds. J. Exp. Biol. 2014, 217, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Martínez-de la Puente, J.; Merino, S.; Tomás, G.; Moreno, J.; Morales, J.; Lobato, E.; Martinez, J. The blood parasite Haemoproteus reduces survival in a wild bird: A medication experiment. Biol. Lett. 2011, 6, 663–665. [Google Scholar] [CrossRef] [PubMed]
- Dunn, J.C.; Goodman, S.J.; Benton, T.G.; Hamer, K.C. Avian blood parasite infection during the non-breeding season: An overlooked issue in declining populations? BMC Ecol. 2013, 13, 30. [Google Scholar] [CrossRef]
- Reinoso-Pérez, M.T.; Canales-Delgadillo, J.C.; Chapa-Vargas, L.; Riego-Ruiz, L. Haemosporidian parasite prevalence, parasitemia, and diversity in three resident bird species at a shrubland dominated landscape of the Mexican highland plateau. Parasites Vectors 2016, 9, 307. [Google Scholar] [CrossRef]


{kind=link}
| Genus | Lineage | Breeding Season | Non-Breeding Season | ||
|---|---|---|---|---|---|
| Number of Chickens (n) | Prevalence (%) | Number of Chickens (n) | Prevalence (%) | ||
| Plasmodium | Gallus 01 | 1 | 1.76 | 0 | 0 |
| Plasmodium | Gallus 04 | 0 | 0 | 2 | 5.13 |
| Leucocytozoon | Gallus 05 | 7 | 12.50 | 2 | 5.13 |
| Leucocytozoon | Gallus 06 | 6 | 10.71 | 1 | 0 |
| Leucocytozoon | Gallus 07 | 5 | 8.93 | 0 | 0 |
| Leucocytozoon | Gallus 08 | 9 | 16.07 | 25 | 64.10 |
| Leucocytozoon | AFR188 | 4 | 7.14 | 0 | 0 |
| Leucocytozoon | AFR227 | 2 | 3.57 | 0 | 0 |
| Leucocytozoon | ANLAT17 | 2 | 3.57 | 0 | 0 |
| Leucocytozoon | CAP4 | 3 | 5.36 | 0 | 0 |
| Leucocytozoon | LANMER05 | 0 | 0 | 1 | 2.56 |
| Leucocytozoon | ORORI04 | 0 | 0 | 2 | 5.13 |
| Leucocytozoon | ZOBOR02 | 17 | 30.36 | 6 | 15.38 |
| Total | 56 | 39 | |||
| Breeding Season | Non-Breeding Season | |||
|---|---|---|---|---|
| Number of Infected Chickens | Number of Uninfected Chickens | Number of Infected Chickens | Number of Uninfected Chickens | |
| Male chickens | 30 | 4 | 20 | 8 |
| Female chickens | 26 | 6 | 19 | 9 |
| Total | 56 | 10 | 39 | 17 |
| Body Parameter | Breeding Season | Non-Breeding Season | Body Parameter | Breeding Season | Non-Breeding Season |
|---|---|---|---|---|---|
| BWt/g | 960.23 ± 15.65 a | 834.43 ± 18.72 b | ED/mm | 12.52 ± 0.12 a | 10.53 ± 0.92 b |
| BLt/mm | 461.88 ± 8.44 a | 479.11 ± 6.93 a | ESD/mm | 10.02 ± 0.11 a | 8.59 ± 0.10 b |
| WL/mm | 215.48 ± 1.95 a | 206.58 ± 1.22 b | BL/mm | 18.38 ± 0.20 a | 17.61 ± 0.17 b |
| TL/mm | 195.97 ± 7.46 a | 200.63 ± 4.71 a | BW/mm | 19.22 ± 0.20 a | 18.34 ± 0.15 b |
| UGL/mm | 7.63 ± 0.16 a | 7.00 ± 0.10 b | BH/mm | 11.33 ± 0.16 a | 12.28 ± 0.12 b |
| UGW/mm | 11.36 ± 0.26 a | 10.52 ± 0.15 a | WS/mm | 7.73 ± 0.17 a | 6.96 ± 0.16 b |
| UGH/mm | 7.73 ± 0.17 a | 6.96 ± 0.16 b | BF1/N | 18.90 ± 0.76 a | 28.09 ± 1.11 b |
| HL/mm | 70.36 ± 0.68 a | 71.11 ± 0.68 a | BF2/N | 18.61 ± 0.76 a | 30.08 ± 1.00 b |
| HB/mm | 28.01 ± 0.22 a | 29.47 ± 0.19 b | BF3/N | 18.65 ± 0.70 a | 29.64 ± 0.98 b |
| HH/mm | 30.45 ± 0.27 a | 33.91 ± 0.34 b | ABF/N | 18.72 ± 0.66 a | 29.27 ± 0.88 b |
| LSt/mm | 31.46 ± 0.30 a | 29.62 ± 0.30 b | MBF/N | 21.14 ± 0.69 a | 33.31 ± 0.97 b |
| TT/mm | 69.66 ± 0.84 a | 73.88 ± 1.00 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Che-Ajuyo, N.M.; Rao, X.; Liu, B.; Deng, Z.; Dong, L.; Liang, W. Effect of Breeding Season on Haemosporidian Infections in Domestic Chickens. Vet. Sci. 2022, 9, 681. https://doi.org/10.3390/vetsci9120681
Che-Ajuyo NM, Rao X, Liu B, Deng Z, Dong L, Liang W. Effect of Breeding Season on Haemosporidian Infections in Domestic Chickens. Veterinary Sciences. 2022; 9(12):681. https://doi.org/10.3390/vetsci9120681
Chicago/Turabian StyleChe-Ajuyo, Nuela Manka’a, Xiaodong Rao, Boye Liu, Zhuqing Deng, Lu Dong, and Wei Liang. 2022. "Effect of Breeding Season on Haemosporidian Infections in Domestic Chickens" Veterinary Sciences 9, no. 12: 681. https://doi.org/10.3390/vetsci9120681
APA StyleChe-Ajuyo, N. M., Rao, X., Liu, B., Deng, Z., Dong, L., & Liang, W. (2022). Effect of Breeding Season on Haemosporidian Infections in Domestic Chickens. Veterinary Sciences, 9(12), 681. https://doi.org/10.3390/vetsci9120681