
Molecular Characterization of CF33 Canine Cell Line and Evaluation of Its Ability to Respond against Infective Stressors in Sight of Anticancer Approaches

 , , , , , and
, , , , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Selection of Reference Genes
2.3. Basal Gene Expression Profiles
2.4. Response to the Infective Stressor
2.4.1. Bacterial Invasion Assessment
2.4.2. Modulation of Innate Immune Response
2.5. ELISA Assay for IL6
2.6. Sequencing of CXCR4
2.7. Statistical Analyses
3. Results
3.1. Selection of Reference Genes
3.2. Basal Gene Expression Profile
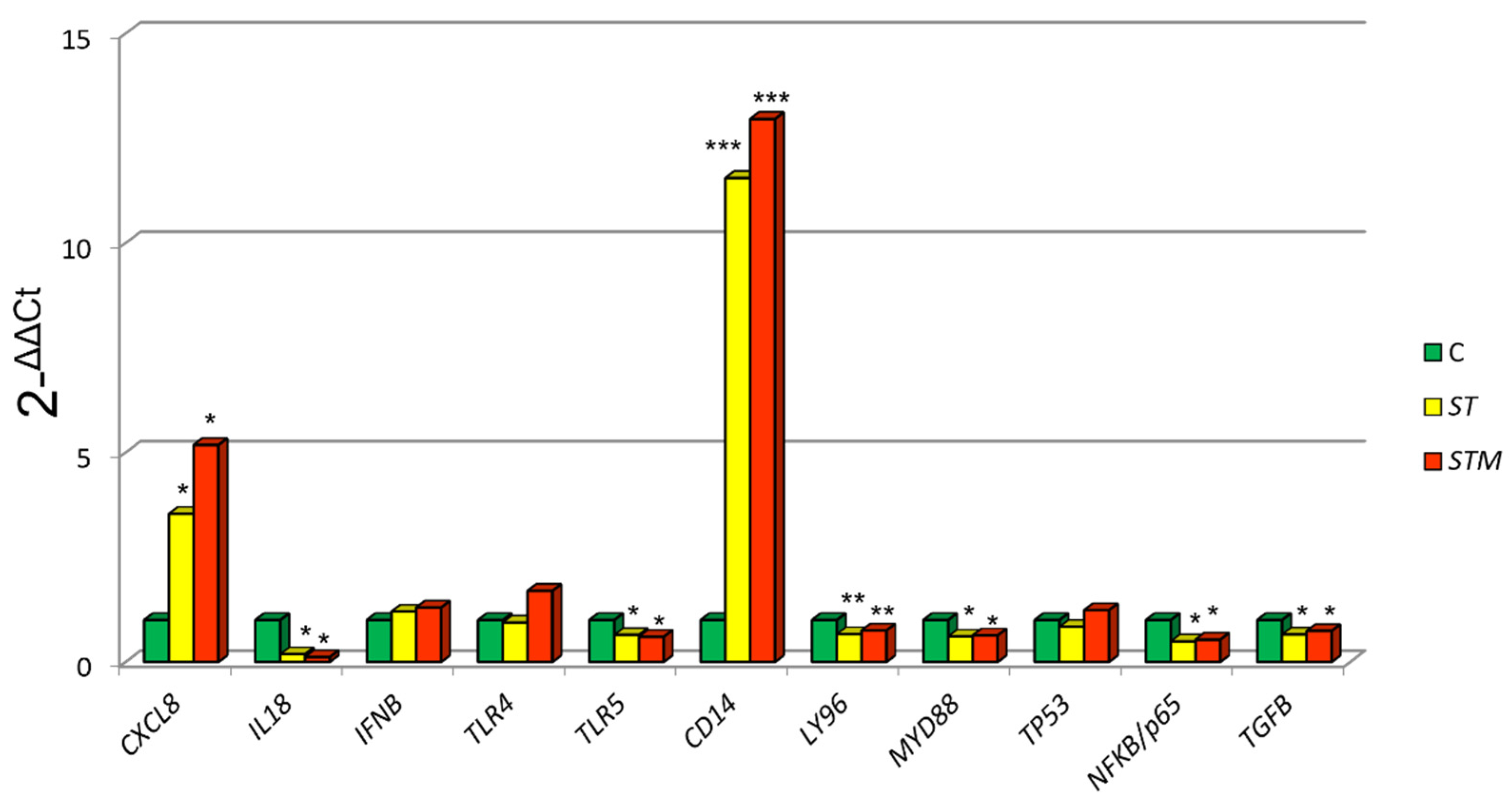
3.3. Response to Infective Stressor
3.3.1. Modulation of Innate Immune Response
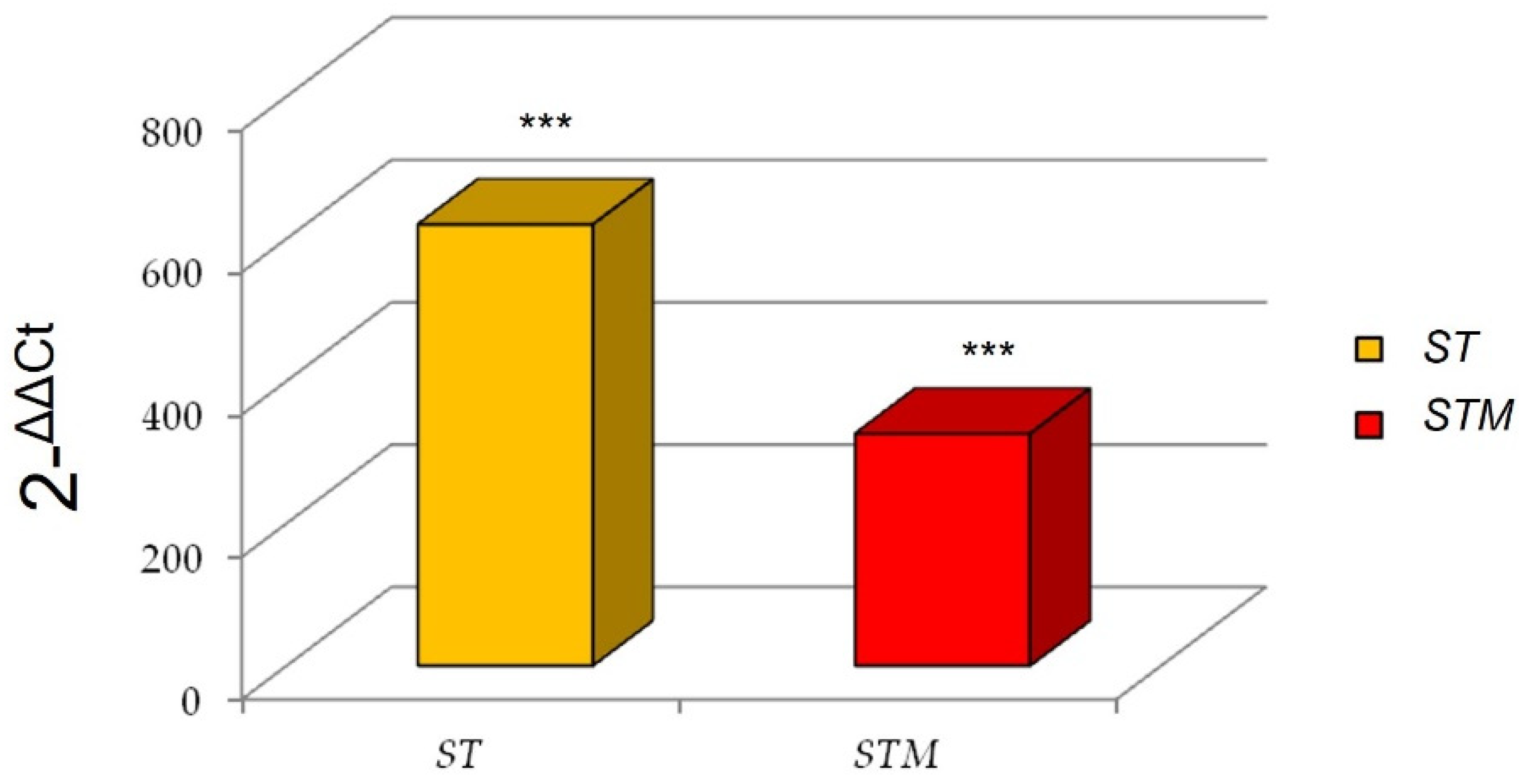
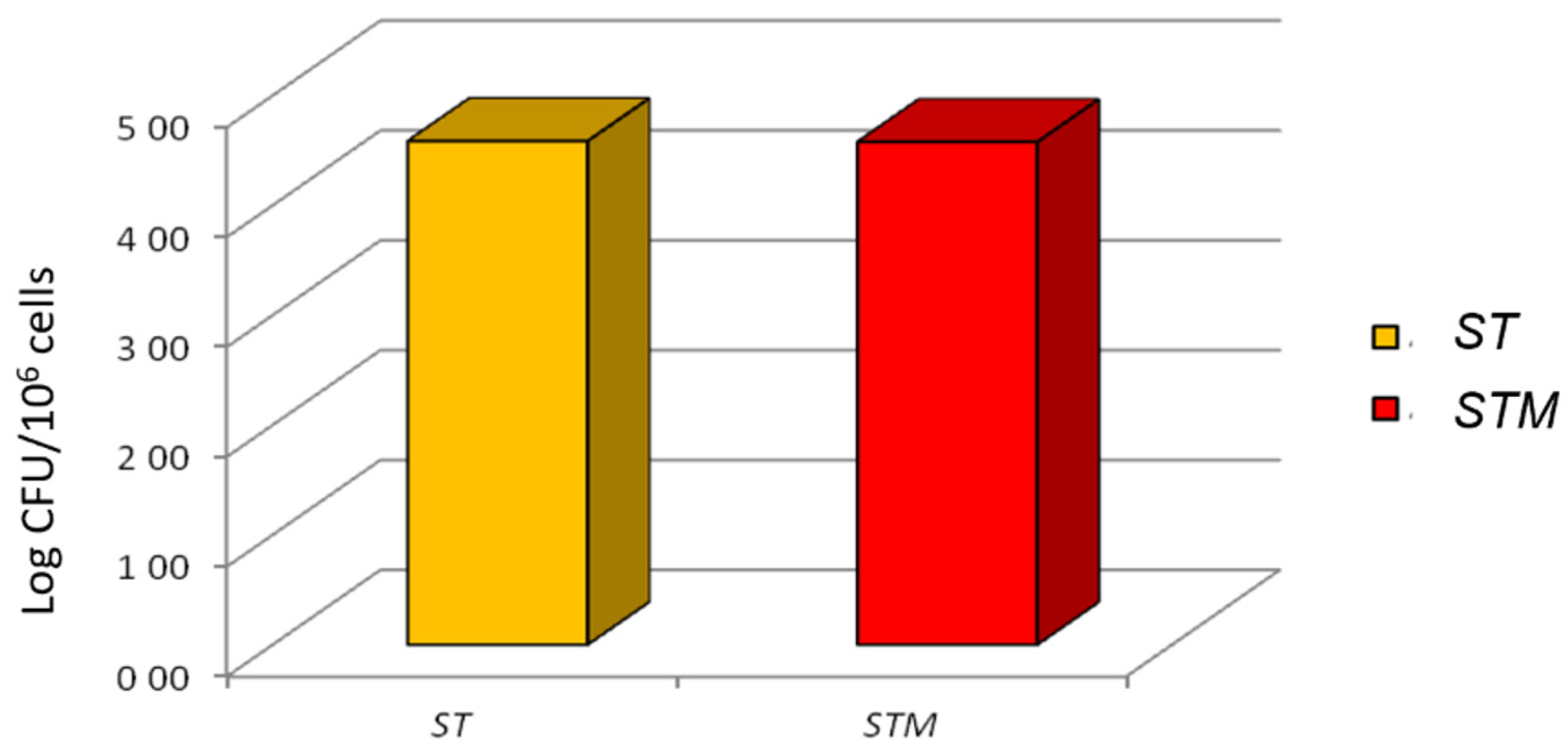
3.3.2. Bacterial Invasion Assessment
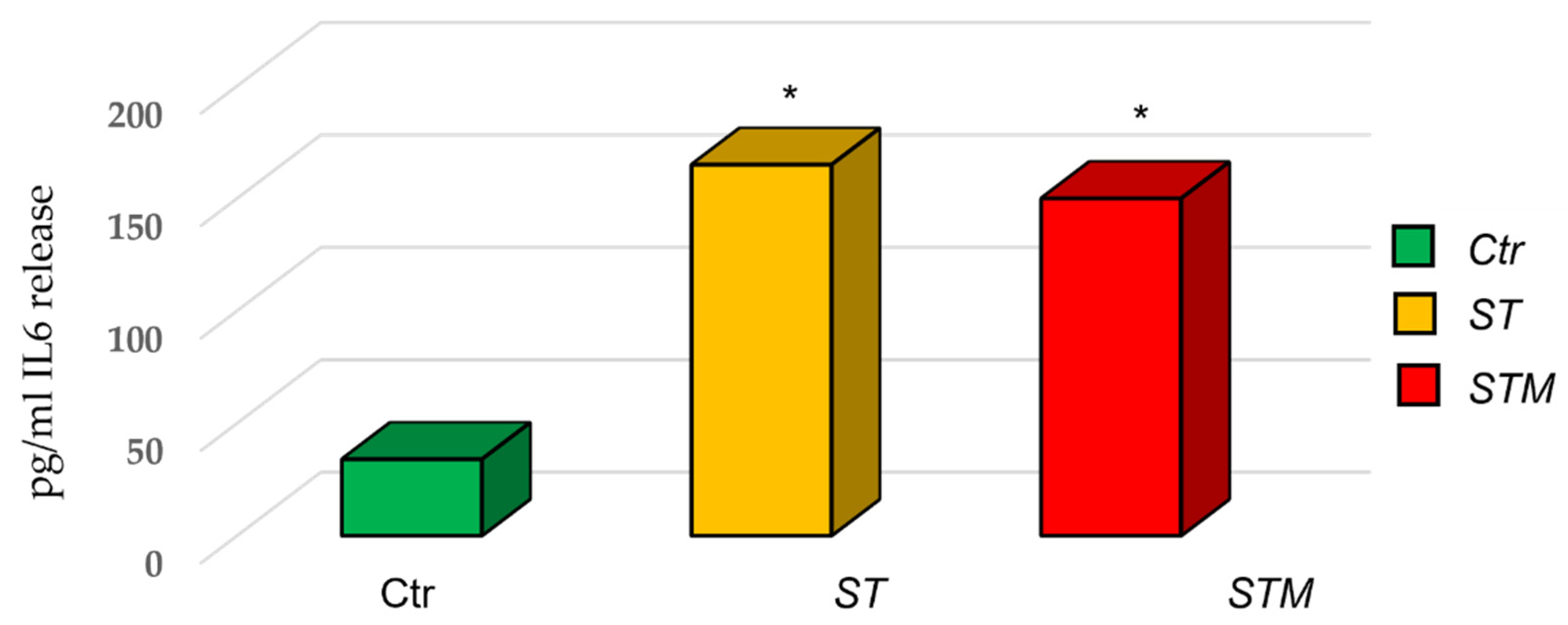
3.4. IL6 ELISA Assay
3.5. CXCR4 Sequencing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nguyen, F.; Peña, L.; Ibisch, C.; Loussouarn, D.; Gama, A.; Rieder, N.; Belousov, A.; Campone, M.; Abadie, J. Canine Invasive Mammary Carcinomas as Models of Human Breast Cancer. Part 1: Natural History and Prognostic Factors. Breast Cancer Res. Treat. 2018, 167, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Frontiers|From Conventional to Precision Therapy in Canine Mammary Cancer: A Comprehensive Review|Veterinary Science. Available online: https://www.frontiersin.org/articles/10.3389/fvets.2021.623800/full (accessed on 25 March 2022).
- Abdelmegeed, S.M.; Mohammed, S. Canine Mammary Tumors as a Model for Human Disease. Oncol. Lett. 2018, 15, 8195–8205. [Google Scholar] [CrossRef] [PubMed]
- Kishima, M.O.; de Oliveira, K.B.; Ariza, C.B.; Oliveira, C.E.C.; Guembarovski, R.L.; Hirata, B.K.B.; de Almeida, F.C.; Vitiello, G.A.F.; Trugilo, K.P.; Guembarovski, A.F.M.L.; et al. Genetic Polymorphism and Expression of CXCR4 in Breast Cancer. Anal. Cell. Pathol. 2015, 2015, 289510. [Google Scholar] [CrossRef]
- Chu, Q.D.; Panu, L.; Holm, N.T.; Li, B.D.L.; Johnson, L.W.; Zhang, S. High Chemokine Receptor CXCR4 Level in Triple Negative Breast Cancer Specimens Predicts Poor Clinical Outcome. J. Surg. Res. 2010, 159, 689–695. [Google Scholar] [CrossRef]
- Scala, S.; Ottaiano, A.; Ascierto, P.A.; Cavalli, M.; Simeone, E.; Giuliano, P.; Napolitano, M.; Franco, R.; Botti, G.; Castello, G. Expression of CXCR4 Predicts Poor Prognosis in Patients with Malignant Melanoma. Clin. Cancer Res. 2005, 11, 1835–1841. [Google Scholar] [CrossRef]
- Abadie, J.; Nguyen, F.; Loussouarn, D.; Peña, L.; Gama, A.; Rieder, N.; Belousov, A.; Bemelmans, I.; Jaillardon, L.; Ibisch, C.; et al. Canine Invasive Mammary Carcinomas as Models of Human Breast Cancer. Part 2: Immunophenotypes and Prognostic Significance. Breast Cancer Res. Treat. 2018, 167, 459–468. [Google Scholar] [CrossRef]
- Al-Mansour, M.A.; Kubba, M.A.G.; Al-Azreg, S.A.; Dribika, S.A. Comparative Histopathology and Immunohistochemistry of Human and Canine Mammary Tumors. Open Vet. J. 2018, 8, 243–249. [Google Scholar] [CrossRef]
- Queiroga, F.L.; Raposo, T.; Carvalho, M.I.; Prada, J.; Pires, I. Canine Mammary Tumors as a Model to Study Human Breast Cancer: Most Recent Findings. Vivo Athens Greece 2011, 25, 455–465. [Google Scholar]
- Kim, T.-M.; Yang, I.S.; Seung, B.-J.; Lee, S.; Kim, D.; Ha, Y.-J.; Seo, M.-K.; Kim, K.-K.; Kim, H.S.; Cheong, J.-H.; et al. Cross-Species Oncogenic Signatures of Breast Cancer in Canine Mammary Tumors. Nat. Commun. 2020, 11, 3616. [Google Scholar] [CrossRef]
- Beetch, M.; Harandi-Zadeh, S.; Yang, T.; Boycott, C.; Chen, Y.; Stefanska, B.; Mohammed, S.I. DNA Methylation Landscape of Triple-Negative Ductal Carcinoma in Situ (DCIS) Progressing to the Invasive Stage in Canine Breast Cancer. Sci. Rep. 2020, 10, 2415. [Google Scholar] [CrossRef]
- Amirkhani Namagerdi, A.; d’Angelo, D.; Ciani, F.; Iannuzzi, C.A.; Napolitano, F.; Avallone, L.; De Laurentiis, M.; Giordano, A. Triple-Negative Breast Cancer Comparison With Canine Mammary Tumors From Light Microscopy to Molecular Pathology. Front. Oncol. 2020, 10, 563779. [Google Scholar] [CrossRef] [PubMed]
- Chirullo, B.; Ammendola, S.; Leonardi, L.; Falcini, R.; Petrucci, P.; Pistoia, C.; Vendetti, S.; Battistoni, A.; Pasquali, P. Attenuated Mutant Strain of Salmonella Typhimurium Lacking the ZnuABC Transporter Contrasts Tumor Growth Promoting Anti-Cancer Immune Response. Oncotarget 2015, 6, 17648–17660. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jahangir, A.; Chandra, D.; Quispe-Tintaya, W.; Singh, M.; Selvanesan, B.C.; Gravekamp, C. Immunotherapy with Listeria Reduces Metastatic Breast Cancer in Young and Old Mice through Different Mechanisms. Oncoimmunology 2017, 6, e1342025. [Google Scholar] [CrossRef] [PubMed]
- Urban-Wojciuk, Z.; Khan, M.M.; Oyler, B.L.; Fåhraeus, R.; Marek-Trzonkowska, N.; Nita-Lazar, A.; Hupp, T.R.; Goodlett, D.R. The Role of TLRs in Anti-Cancer Immunity and Tumor Rejection. Front. Immunol. 2019, 10, 2388. [Google Scholar] [CrossRef]
- Sedighi, M.; Zahedi Bialvaei, A.; Hamblin, M.R.; Ohadi, E.; Asadi, A.; Halajzadeh, M.; Lohrasbi, V.; Mohammadzadeh, N.; Amiriani, T.; Krutova, M.; et al. Therapeutic Bacteria to Combat Cancer; Current Advances, Challenges, and Opportunities. Cancer Med. 2019, 8, 3167–3181. [Google Scholar] [CrossRef] [PubMed]
- Badie, F.; Ghandali, M.; Tabatabaei, S.A.; Safari, M.; Khorshidi, A.; Shayestehpour, M.; Mahjoubin-Tehran, M.; Morshedi, K.; Jalili, A.; Tajiknia, V.; et al. Use of Salmonella Bacteria in Cancer Therapy: Direct, Drug Delivery and Combination Approaches. Front. Oncol. 2021, 11, 624759. [Google Scholar] [CrossRef] [PubMed]
- Silvertown, J.D.; Geddes, B.J.; Summerlee, A.J.S. Adenovirus-Mediated Expression of Human Prorelaxin Promotes the Invasive Potential of Canine Mammary Cancer Cells. Endocrinology 2003, 144, 3683–3691. [Google Scholar] [CrossRef]
- Saito, T.; Tamura, D.; Nakamura, T.; Makita, Y.; Ariyama, H.; Komiyama, K.; Yoshihara, T.; Asano, R. 4-Methylumbelliferone Leads to Growth Arrest and Apoptosis in Canine Mammary Tumor Cells. Oncol. Rep. 2013, 29, 335–342. [Google Scholar] [CrossRef]
- Modesto, P.; Fernandez, J.L.C.; Martini, I.; Zoccola, R.; Pugliano, M.C.; De Ciucis, C.G.; Goria, M.; Ferrari, A.; Razzuoli, E. Characterization of D-17 Canine Osteosarcoma Cell Line and Evaluation of Its Ability to Response to Infective Stressor Used as Alternative Anticancer Therapy. Animals 2020, 10, 1981. [Google Scholar] [CrossRef]
- Qiu, H.; Lin, D. Roles of DNA Mutation in the Coding Region and DNA Methylation in the 5′ Flanking Region of BRCA1 in Canine Mammary Tumors. J. Vet. Med. Sci. 2016, 78, 943–949. [Google Scholar] [CrossRef]
- Razzuoli, E.; Amadori, M.; Lazzara, F.; Bilato, D.; Ferraris, M.; Vito, G.; Ferrari, A. Salmonella Serovar-Specific Interaction with Jejunal Epithelial Cells. Vet. Microbiol. 2017, 207, 219–225. [Google Scholar] [CrossRef]
- Capellini, F.M.; Vencia, W.; Amadori, M.; Mignone, G.; Parisi, E.; Masiello, L.; Vivaldi, B.; Ferrari, A.; Razzuoli, E. Characterization of MDCK Cells and Evaluation of Their Ability to Respond to Infectious and Non-Infectious Stressors. Cytotechnology 2020, 72, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Razzuoli, E.; Villa, R.; Ferrari, A.; Amadori, M. A Pig Tonsil Cell Culture Model for Evaluating Oral, Low-Dose IFN-α Treatments. Vet. Immunol. Immunopathol. 2014, 160, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Razzuoli, E.; Villa, R.; Amadori, M. IPEC-J2 Cells as Reporter System of the Anti-Inflammatory Control Actions of Interferon-Alpha. J. Interferon Cytokine Res. 2013, 33, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Ghoncheh, M.; Pournamdar, Z.; Salehiniya, H. Incidence and Mortality and Epidemiology of Breast Cancer in the World. Asian Pac. J. Cancer Prev. APJCP 2016, 17, 43–46. [Google Scholar] [CrossRef]
- Global Cancer Rates Could Increase by 50% to 15 Million by 2020. Available online: https://www.who.int/news/item/03-04-2003-global-cancer-rates-could-increase-by-50-to-15-million-by-2020 (accessed on 25 March 2022).
- Pinho, S.S.; Carvalho, S.; Cabral, J.; Reis, C.A.; Gärtner, F. Canine Tumors: A Spontaneous Animal Model of Human Carcinogenesis. Transl. Res. J. Lab. Clin. Med. 2012, 159, 165–172. [Google Scholar] [CrossRef]
- Amadori, D.; Bertoni, L.; Flamigni, A.; Savini, S.; De Giovanni, C.; Casanova, S.; De Paola, F.; Amadori, A.; Giulotto, E.; Zoli, W. Establishment and Characterization of a New Cell Line from Primary Human Breast Carcinoma. Breast Cancer Res. Treat. 1993, 28, 251–260. [Google Scholar] [CrossRef]
- Oladejo, M.; Paterson, Y.; Wood, L.M. Clinical Experience and Recent Advances in the Development of Listeria-Based Tumor Immunotherapies. Front. Immunol. 2021, 12, 642316. [Google Scholar] [CrossRef]
- de Andrés, P.J.; Illera, J.C.; Cáceres, S.; Díez, L.; Pérez-Alenza, M.D.; Peña, L. Increased Levels of Interleukins 8 and 10 as Findings of Canine Inflammatory Mammary Cancer. Vet. Immunol. Immunopathol. 2013, 152, 245–251. [Google Scholar] [CrossRef]
- Khademalhosseini, M.; Arababadi, M.K. Toll-like Receptor 4 and Breast Cancer: An Updated Systematic Review. Breast Cancer 2019, 26, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Canadas, A.; Santos, M.; Nogueira, A.; Assis, J.; Gomes, M.; Lemos, C.; Medeiros, R.; Dias-Pereira, P. Canine Mammary Tumor Risk Is Associated with Polymorphisms in RAD51 and STK11 Genes. J. Vet. Diagn. Investig. 2018, 30, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.-Z.; Jones, R.F.; Juhasz, C.; Gibson, H.; Veenstra, J. Evolution of Animal Models in Cancer Vaccine Development. Vaccine 2015, 33, 7401–7407. [Google Scholar] [CrossRef] [PubMed]
- Nance, R.L.; Sajib, A.M.; Smith, B.F. Canine Models of Human Cancer: Bridging the Gap to Improve Precision Medicine. Prog. Mol. Biol. Transl. Sci. 2022, 189, 67–99. [Google Scholar] [CrossRef]
- Ariyarathna, H.; Thomson, N.; Aberdein, D.; Munday, J.S. Chemokine Gene Expression Influences Metastasis and Survival Time of Female Dogs with Mammary Carcinoma. Vet. Immunol. Immunopathol. 2020, 227, 110075. [Google Scholar] [CrossRef]
- Klopfleisch, R.; Gruber, A.D. Increased Expression of BRCA2 and RAD51 in Lymph Node Metastases of Canine Mammary Adenocarcinomas. 2009. Available online: https://journals.sagepub.com/doi/10.1354/vp.08-VP-0212-K-FL?url_ver=Z39.88-2003&rfr_id=ori:rid:crossref.org&rfr_dat=cr_pub%20%200pubmed (accessed on 25 March 2022).
- Oliveira, T.F.; Maués, T.; Ramundo, M.S.; Figueiredo, A.M.S.; De Mello, M.F.V.; El-Jaick, K.B.; Ferreira, M.D.L.G.; Ferreira, A.M.R. TP53 Gene Expression Levels and Tumor Aggressiveness in Canine Mammary Carcinomas. J. Veter-Diagn. Investig. 2017, 29, 865–868. [Google Scholar] [CrossRef]
- Maués, T.; de Oliveira, T.F.; El-Jaick, K.B.; Figueiredo, A.M.S.; Ferreira, M.D.L.G.; Ferreira, A.M.R. PGAM1 and TP53 MRNA Levels in Canine Mammary Carcinomas—Short Communication. Acta Veter-Hung. 2021, 69, 50–54. [Google Scholar] [CrossRef]
- Kaszak, I.; Ruszczak, A.; Kanafa, S.; Kacprzak, K.; Król, M.; Jurka, P. Current Biomarkers of Canine Mammary Tumors. Acta Vet. Scand. 2018, 60, 66. [Google Scholar] [CrossRef]
- Cocco, S.; Piezzo, M.; Calabrese, A.; Cianniello, D.; Caputo, R.; Di Lauro, V.; Fusco, G.; di Gioia, G.; Licenziato, M.; de Laurentiis, M. Biomarkers in Triple-Negative Breast Cancer: State-of-the-Art and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 4579. [Google Scholar] [CrossRef]
- Kim, S.-H.; Seung, B.-J.; Cho, S.-H.; Lim, H.-Y.; Bae, M.-K.; Sur, J.-H. Dysregulation of PI3K/Akt/PTEN Pathway in Canine Mammary Tumor. Animals 2021, 11, 2079. [Google Scholar] [CrossRef]
- Asproni, P.; Millanta, F.; Ressel, L.; Podestà, F.; Parisi, F.; Vannozzi, I.; Poli, A. An Immunohistochemical Study of the PTEN/AKT Pathway Involvement in Canine and Feline Mammary Tumors. Animals 2021, 11, 365. [Google Scholar] [CrossRef] [PubMed]
- Gray, C.W.; Coster, A.C.F. From Insulin to Akt: Time Delays and Dominant Processes. J. Theor. Biol. 2020, 507, 110454. [Google Scholar] [CrossRef]
- Wee, P.; Wang, Z. Epidermal Growth Factor Receptor Cell Proliferation Signaling Pathways. Cancers 2017, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Singer, J.; Weichselbaumer, M.; Stockner, T.; Mechtcheriakova, D.; Sobanov, Y.; Bajna, E.; Wrba, F.; Horvat, R.; Thalhammer, J.G.; Willmann, M.; et al. Comparative Oncology: ErbB-1 and ErbB-2 Homologues in Canine Cancer Are Susceptible to Cetuximab and Trastuzumab Targeting. Mol. Immunol. 2012, 50, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Worzfeld, T.; Swiercz, J.M.; Looso, M.; Straub, B.K.; Sivaraj, K.K.; Offermanns, S. ErbB-2 Signals through Plexin-B1 to Promote Breast Cancer Metastasis. J. Clin. Investig. 2012, 122, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, T.; Mizoshita, T.; Tatematsu, M. Animal Models of Stomach Carcinogenesis. Toxicol. Pathol. 2007, 35, 636–648. [Google Scholar] [CrossRef] [PubMed]
- Grano, F.G.; Silva, J.E.D.S.; Melo, G.D.; de Souza, M.S.; Lima, V.M.; Machado, G.F. Toll-like Receptors and Cytokines in the Brain and in Spleen of Dogs with Visceral Leishmaniosis. Vet. Parasitol. 2018, 253, 30–38. [Google Scholar] [CrossRef]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, Inflammation, and Cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef]
- Zuccari, D.; Castro, R.; Gelaleti, G.; Mancini, U. Interleukin-8 Expression Associated with Canine Mammary Tumors. Genet. Mol. Res. GMR 2011, 10, 1522–1532. [Google Scholar] [CrossRef]
- Gelaleti, G.B.; Jardim, B.V.; Leonel, C.; Moschetta, M.G.; Zuccari, D.A.P.D.C. Interleukin-8 as a Prognostic Serum Marker in Canine Mammary Gland Neoplasias. Vet. Immunol. Immunopathol. 2012, 146, 106–112. [Google Scholar] [CrossRef]
- Beck, T.C.; Gomes, A.C.; Cyster, J.G.; Pereira, J.P. CXCR4 and a Cell-Extrinsic Mechanism Control Immature B Lymphocyte Egress from Bone Marrow. J. Exp. Med. 2014, 211, 2567–2581. [Google Scholar] [CrossRef] [PubMed]
- Pozzobon, T.; Goldoni, G.; Viola, A.; Molon, B. CXCR4 Signaling in Health and Disease. Immunol. Lett. 2016, 177, 6–15. [Google Scholar] [CrossRef]
- Peng, S.-B.; Zhang, X.; Paul, D.; Kays, L.M.; Gough, W.; Stewart, J.; Uhlik, M.T.; Chen, Q.; Hui, Y.-H.; Zamek-Gliszczynski, M.J.; et al. Identification of LY2510924, a Novel Cyclic Peptide CXCR4 Antagonist That Exhibits Antitumor Activities in Solid Tumor and Breast Cancer Metastatic Models. Mol. Cancer Ther. 2015, 14, 480–490. [Google Scholar] [CrossRef] [PubMed]
- Furusato, B.; Mohamed, A.; Uhlén, M.; Rhim, J.S. CXCR4 and Cancer. Pathol. Int. 2010, 60, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, U.; Siranosian, B.; Ha, G.; Tang, H.; Oren, Y.; Hinohara, K.; Strathdee, C.A.; Dempster, J.; Lyons, N.J.; Burns, R.; et al. Genetic and Transcriptional Evolution Alters Cancer Cell Line Drug Response. Nature 2018, 560, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Rogez, B.; Pascal, Q.; Bobillier, A.; Machuron, F.; Lagadec, C.; Tierny, D.; Le Bourhis, X.; Chopin, V. CD44 and CD24 Expression and Prognostic Significance in Canine Mammary Tumors. Vet. Pathol. 2019, 56, 377–388. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Product Length (bp) | Accession Number |
|---|---|---|---|
| RPS5 | F: TCACTGGTGAGAACCCCCT R: CCTGATTCACACGGCGTAG | 141 | XM_533568 |
| B2M | F: TCCTCATCCTCCTCGCT R: TTCTCTGCTGGGTGTCG | 84 | NM_001284479.1 |
| GUSB | F: AGACGTTCCAAGTACCCC R: AGGTGTGGTGTAGAGGAGCAC | 102 | NM_001003191 |
| GAPDH | F: CTGGGGCTCACTTGAAAGG R: GGAGGCATTGCTGACAATC | 131 | NM_001003142.2 |
| HPRT1 | F: CTGAAGAGCTACTGTAATGACCAGTC R: CTTTTCACCAGCAAGCTTGCAACC | 197 | NM_001003357.2 |
| HRNPH1 | F: CTCACTATGATCCACCACG R: TAGCCTCCATAACCTCCAC | 150 | XM_025446760.3 |
| BACT | F: ACGGAGCGTGGCTACAGC′ R: TCCTTGATGTCACGCACGA | 61 | NM_001195845.3 |
| SRPR | F: GCTTCAGGATCTGGACTGC R: GTTCCCTTGGTAGCACTGG | 80 | XM_03866445.1 |
| TBP | F: TCCACAGCCTATCCAGAACA R: CTGCTGCTGTTGTCTCTGCT | 66 | XM_005627735.4 |
| RPL13A | F: GGGGCAGGTCCTGGTGCTCG R: CCAGGTACTTCAACTTGTTTCTGTAG | 158 | NM_001313766.1 |
| RPS19 | F: CCTTCCTCAAAAAGTCTGGG R: GCTGTGGAAGCAGCTCGC | 124 | XM_005616513.4 |
| SDHA | F: GGTGGCACTTCTACGACACC R: CCATAATTCTCCAGCTCTACC | 112 | XM_014110317.3 |
| Gene | Primers | Product Length (bp) | Accession Number |
|---|---|---|---|
| IL1B | F: TGCAAAACAGATGCGGATAA R: GTAACTTGCAGTCCACCGATT | 64 | NM_001037971.1 |
| IL2 | F: CCTCAACTCCTGCCACAATGT R: TGCGACAAGTACAAGCGTCAGT | 71 | NM_001003305.2 |
| IL4 | F: TGCAGAGCTGCTACTGTACTGCGGC R: CATGCTGCTGAGGTTCCTGT | 90 | NM_001003159.1 |
| IL5 | F: GCCTATGTTTCTGCCTTTGC R: GGTTCCCATCGCCTATCA | 106 | NM_001006950.1 |
| IL6 | F: TCCAGAACAACTATGAGGGTGA R: TCCTGATTCTTTACCTTGCTCTT | 100 | NM_001003301.1 |
| CXCL8 | F: TGATTGACAGTGGCCCACATTGTG R: GTCCAGGCACACCTCATTTC | 77 | NM_001003200.1 |
| IL10 | F: CGACCCAGACATCAAGAACC R: CACAGGGAAGAAATCGGTGA | 101 | NM_001003077.1 |
| IL12B | F: TGGAGGTCAGCTGGGAATACC R: TGCAAAATGTCAGGGAGAAGTA | 69 | NM_001003292.1 |
| IL15 | F: ACTTCCATCCAGTGCTACTT R: CGAGCGAGATAACACCTAAC | 271 | NM_001197188.1 |
| IL16 | F: CCAGTCCAAGGGGATTACAG R: TGAGAATGAGCGGTTGTG | 100 | XM_005618407.4 |
| IL17A | F: ACTCCAGAAGGCCCTCAGATTA R: GATTCCAAGGTGAGGTAGATCG | 51 | NM_001165878.1 |
| IL18 | F: CTCTCCTGTAAGAACAAAACTATTTCCTT R: GAACACTTCTCTGAAAGAATATGATGTCA | 100 | NM_001003169.1 |
| IL23A | F: ACAGAACGGACAGCATCAGG R: CGCTGCCTGCTTCTCAAATC | 101 | XM_538231.7 |
| IL27 | F: TTACTGCTCTCCCTGCTCCT R: TTGAACTCCCTCCGCAACTC | 101 | XM_038668726.1 |
| IFNG | F: CCAGATCATTCAAAGGAGCA R: CGTTCACAGGAATTTGAATCAG | 116 | NM_001003174.1 |
| TNFA | F: CGTCCATTCTTGCCCAAAC R: AGCCCTGAGCCCTTAATTC | 94 | NM_001003244.4 |
| TLR4 | F: GCTGGATGGTAAACCGTGGA R: AGCACAGTGGCAGGTACATC | 158 | NM_001002950.3 |
| TLR5 | F: CCAGGACCAGACGTTCAGAT R: GCCCAGGAAGATGGTGTCTA | 109 | NM_001197176.1 |
| BRCA1 | F: CAGAGAGATACCATGCAAGATAAC R: CTCTTTCTGATGCGTTTTGTTCCG | 172 | NM_001013416.1 |
| CD14 | F: GCCGGGCCTCAAGGTACT R: TCGTGCGCAGGAAAAAGC | 61 | XM_843653.6 |
| CD44 | F: CAAGGCTTTCAACAGCACCC R: TACGTGTCGTACTGGGAGGT | 192 | NM_001197022.2 |
| CXCR4 | F: GCGTCTGGATACCTGCTCTC R: GATACCCGGCAGGATAAGGC | 163 | DQ182699.1 |
| ERBB2 | F: CTGAGGGCCGATATACCTTC R: TCACCTCTTGGTTGTTCAGG | 114 | NM_001003217.3 |
| LY96 | F: GGGAATACGATTTTCTAAGGGACAA R: CGGTAAAATTCAAACAAAAGAGCTT | 92 | XM_848045.5 |
| MYD88 | F: GAGGAGATGGGCTTCGAGTA R: GTTCCACCAACACGTCGTC | 160 | XM_534223.7 |
| TP53 | F: CGTTTGGGGTTCCTGCATTC R: CACTACTGTCAGAGCAGCGT | 232 | NM_001389218.1 |
| NF-KB/p65 | F: TGTAAAGAAGCGGGACCTGG R: AGAGTTTCGGTTCACTCGGC | 250 | XM_038424975.1 |
| PTEN | F: GTGAAGCTGTACTTCACAA R: CTGGGTCAGAGTCAGTGGTG | 136 | NM_001003192.1 |
| RAD51 | F: GGAGAAGGAAAGGCCATGTA R: GGGTCTGGTGGTCTGTGTT | 148 | NM_001003043.1 |
| TGFB | F: CAAGTAGACATTAACGGGTTCAGTTC R: GGTCGGTTCATGCCATGAAT | 70 | XM_038656896.1 |
| Primer | Position | Product Length (bp) | Accession Number | |
|---|---|---|---|---|
| CXCR4 F | TCT GTG GCA GAC CTC CTC TT | F 266–285 R 611–630 | 364 | NM_001048026.1 |
| CXCR4 R | TGA AAC TGG AAC ACC ACC AA | |||
| CXCR4 F7 | TGA CTC CAT GAA GGA ACC CTG | F 88–108 R 971–990 | 902 | NM_001048026.1 |
| CXCR4 R2 | CTG CTC ACA GAG GTG AGT GC | |||
| CXCR4 Fow 3a | GTC ATC CTG TCC TGC TAC TG | F 665–684 R 296–314 | Sequencing 902 bp | NM_001048026.1 |
| CXCR4 Rev 3b | CAA CTG CCC AGA AGG GAA G | |||
| Gene | Stability (p) |
|---|---|
| B2M | 0.031 |
| BACT | 0.027 |
| RPS5 | 0.014 |
| RPS19 | 0.041 |
| GAPDH | 0.045 |
| HPRT1 | 0.051 |
| RPL13A | 0.04 |
| HRNPH1 | 0.051 |
| SRPR | 0.042 |
| TBP | 0.048 |
| SDHA | 0.039 |
| GUSB | 0.044 |
| Gene | Expression | Cq ± SD |
|---|---|---|
| IL1B | − | ND |
| IL2 | − | ND |
| IL4 | ± | 38.3 ± 0.1 |
| IL5 | + | 37.7 ± 0.8 |
| IL6 | ± | 38.0 ± 0.9 |
| CXCL8 | + | 35.7 ± 0.8 |
| IL10 | − | ND |
| IL12B | ± | 38.7 ± 0.1 |
| IL15 | − | ND |
| IL16 | + | 38.7 ± 0.5 |
| IL17A | − | ND |
| IL18 | + | 37.7 ± 0.6 |
| IL23A | ± | 37.3 ± 0.6 |
| IL27 | − | ND |
| IFNG | − | ND |
| TNFA | ± | 37.1 ± 1.1 |
| TLR4 | ± | 38.2 ± 0.6 |
| TLR5 | ± | 38.1 ± 1.0 |
| BRCA1 | + | 32.5 ± 0.8 |
| CD14 | ± | 37.8 ± 0.8 |
| CD44 | + | 37.1 ± 1.1 |
| CXCR4 | ± | 36.8 ± 0.3 |
| ERB-B2 | ± | 38.1 ± 0.6 |
| LY96 | + | 27.1 ± 0.5 |
| MYD88 | + | 37.2 ± 0.7 |
| TP53 | ± | 35.8 ± 0.5 |
| NF-KB/(p65) | ± | 38.8 ± 0.7 |
| PTEN | ± | 38.3 ± 1.2 |
| RAD51 | + | 32.5 ± 1.3 |
| TGFB1 | ± | 37.1 ± 3.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Razzuoli, E.; De Ciucis, C.G.; Chirullo, B.; Varello, K.; Zoccola, R.; Guardone, L.; Petrucci, P.; Rubini, D.; Bozzetta, E.; Goria, M.; et al. Molecular Characterization of CF33 Canine Cell Line and Evaluation of Its Ability to Respond against Infective Stressors in Sight of Anticancer Approaches. Vet. Sci. 2022, 9, 543. https://doi.org/10.3390/vetsci9100543
Razzuoli E, De Ciucis CG, Chirullo B, Varello K, Zoccola R, Guardone L, Petrucci P, Rubini D, Bozzetta E, Goria M, et al. Molecular Characterization of CF33 Canine Cell Line and Evaluation of Its Ability to Respond against Infective Stressors in Sight of Anticancer Approaches. Veterinary Sciences. 2022; 9(10):543. https://doi.org/10.3390/vetsci9100543
Chicago/Turabian StyleRazzuoli, Elisabetta, Chiara Grazia De Ciucis, Barbara Chirullo, Katia Varello, Roberto Zoccola, Lisa Guardone, Paola Petrucci, Danja Rubini, Elena Bozzetta, Maria Goria, and et al. 2022. "Molecular Characterization of CF33 Canine Cell Line and Evaluation of Its Ability to Respond against Infective Stressors in Sight of Anticancer Approaches" Veterinary Sciences 9, no. 10: 543. https://doi.org/10.3390/vetsci9100543
APA StyleRazzuoli, E., De Ciucis, C. G., Chirullo, B., Varello, K., Zoccola, R., Guardone, L., Petrucci, P., Rubini, D., Bozzetta, E., Goria, M., Fruscione, F., & Modesto, P. (2022). Molecular Characterization of CF33 Canine Cell Line and Evaluation of Its Ability to Respond against Infective Stressors in Sight of Anticancer Approaches. Veterinary Sciences, 9(10), 543. https://doi.org/10.3390/vetsci9100543