Protective Effect of Hydroxytyrosol on LPS-Induced Inflammation and Oxidative Stress in Bovine Endometrial Epithelial Cell Line

 ,
,  ,
,  , ,
, ,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Treatment
2.3. Cell Viability Assay
2.4. ELISA
2.5. Reactive Oxygen Species (ROS) Evaluation
2.6. RNA Extraction-cDNA Synthesis
2.7. Real-Time PCR
2.8. Statistical Analysis
3. Results
3.1. HT Effect on BEND Cells Viability
3.2. HT Effect on LPS-Induced Inflammatory Response and Oxidative Stress in BEND Cells
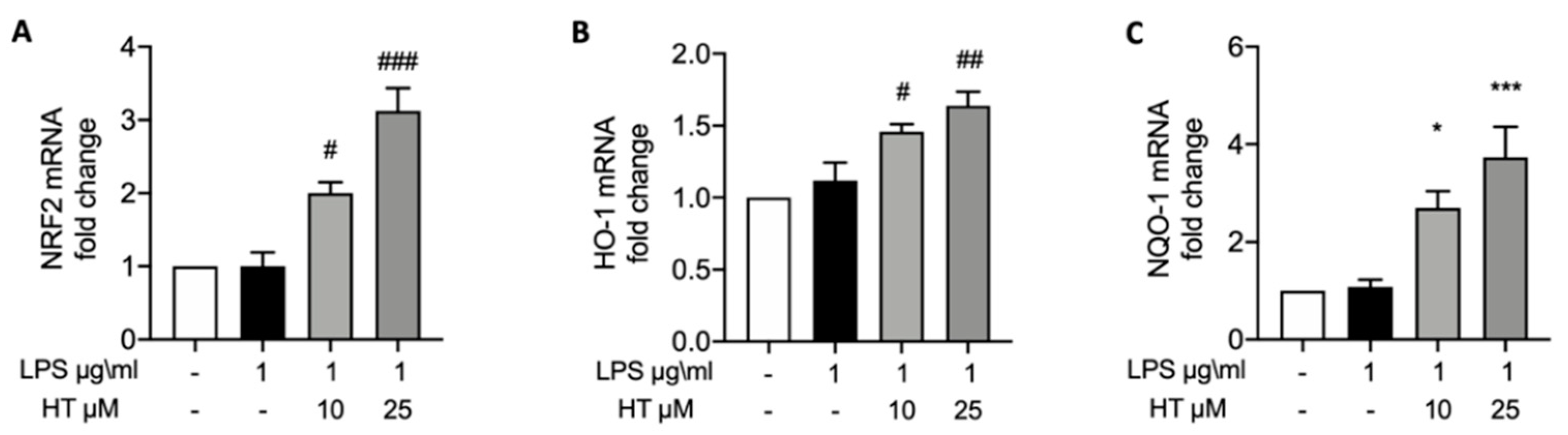
3.3. HT Effect on LPS-Induced Nrf2 Pathway
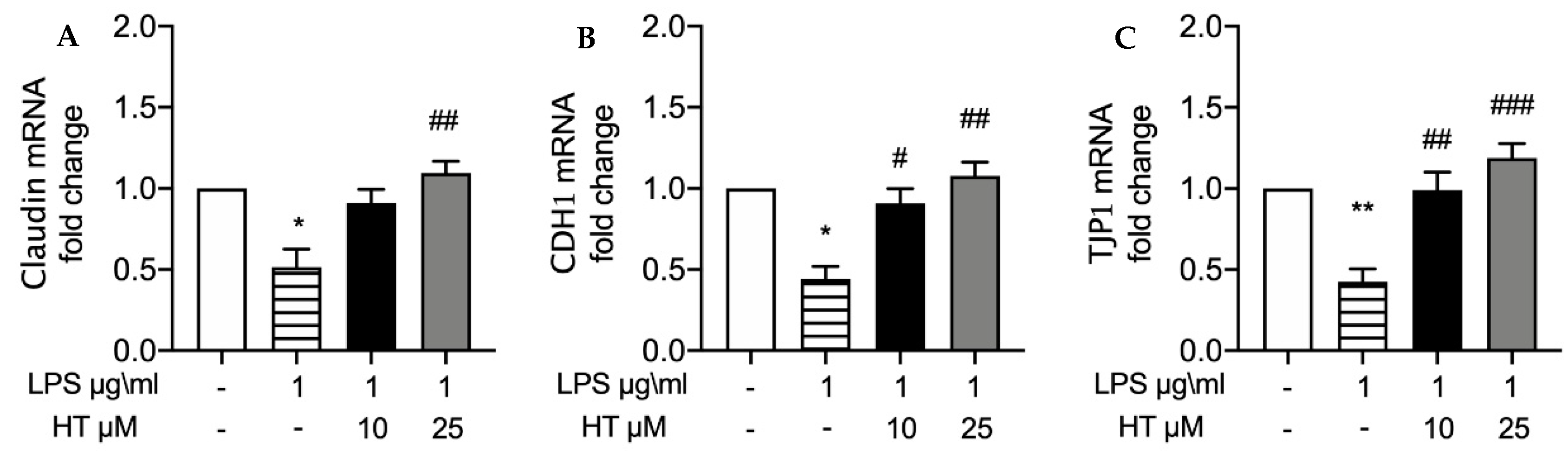
3.4. HT Effect on and Expressions of Tight Junction Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hansen, P.J.; Soto, P.; Natzke, R.P. Mastitis and fertility in cattle—Possible involvement of inflammation or immune activation in embryonic mortality. Am. J. Reprod. Immunol. 2004, 51, 294–301. [Google Scholar] [CrossRef]
- Siemieniuch, M.J.; Szostek, A.Z.; Gajos, K.; Kozdrowski, R.; Nowak, M.; Okuda, K. Type of Inflammation Differentially Affects Expression of Interleukin 1beta and 6, Tumor Necrosis Factor-alpha and Toll-Like Receptors in Subclinical Endometritis in Mares. PLoS ONE 2016, 11, e0154934. [Google Scholar] [CrossRef]
- Burdet, J.; Rubio, A.P.; Salazar, A.I.; Ribeiro, M.L.; Ibarra, C.; Franchi, A.M. Inflammation, infection and preterm birth. Curr. Pharm. Des. 2014, 20, 4741–4748. [Google Scholar] [CrossRef]
- LeBlanc, S.J.; Osawa, T.; Dubuc, J. Reproductive tract defense and disease in postpartum dairy cows. Theriogenology 2011, 76, 1610–1618. [Google Scholar] [CrossRef]
- Koyama, T.; Omori, R.; Koyama, K.; Matsui, Y.; Sugimoto, M. Optimization of diagnostic methods and criteria of endometritis for various postpartum days to evaluate infertility in dairy cows. Theriogenology 2018, 119, 225–232. [Google Scholar] [CrossRef]
- Roche, J.R.; Burke, C.R.; Crookenden, M.A.; Heiser, A.; Loor, J.L.; Meier, S.; Mitchell, M.D.; Phyn, C.V.C.; Turner, S.A. Fertility and the transition dairy cow. Reprod. Fertil. Dev. 2017, 30, 85–100. [Google Scholar] [CrossRef]
- Raheem, K.A. An insight into maternal recognition of pregnancy in mammalian species. J. Saudi Soc. Agric. Sci. 2017, 16, 1–6. [Google Scholar] [CrossRef]
- Herath, S.; Fischer, D.P.; Werling, D.; Williams, E.J.; Lilly, S.T.; Dobson, H.; Bryant, C.E.; Sheldon, I.M. Expression and function of Toll-like receptor 4 in the endometrial cells of the uterus. Endocrinology 2006, 147, 562–570. [Google Scholar] [CrossRef]
- Amjadi, F.; Salehi, E.; Mehdizadeh, M.; Aflatoonian, R. Role of the innate immunity in female reproductive tract. Adv. Biomed. Res. 2014, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, I.M.; Owens, S.E. Postpartum uterine infection and endometritis in dairy cattle. Anim. Reprod. 2018, 14, 622–629. [Google Scholar] [CrossRef]
- Dohmen, M.; Joop, K.; Sturk, A.; Bols, P.; Lohuis, J. Relationship between intra-uterine bacterial contamination, endotoxin levels and the development of endometritis in postpartum cows with dystocia or retained placenta. Theriogenology 2000, 54, 1019–1032. [Google Scholar] [CrossRef]
- Rosenfeld, Y.; Shai, Y. Lipopolysaccharide (Endotoxin)-host defense antibacterial peptides interactions: Role in bacterial resistance and prevention of sepsis. Biochim. Biophys. Acta 2006, 1758, 1513–1522. [Google Scholar] [CrossRef] [PubMed]
- Harju, K.; Ojaniemi, M.; Rounioja, S.; Glumoff, V.; Paananen, R.; Vuolteenaho, R.; Hallman, M. Expression of toll-like receptor 4 and endotoxin responsiveness in mice during perinatal period. Pediatr. Res. 2005, 57, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Healy, L.L.; Cronin, J.G.; Sheldon, I.M. Endometrial cells sense and react to tissue damage during infection of the bovine endometrium via interleukin 1. Sci. Rep. 2014, 4, 7060. [Google Scholar] [CrossRef]
- Splichal, I.; Trebichavsky, I. Cytokines and other important inflammatory mediators in gestation and bacterial intraamniotic infections. Folia Microbiol. 2001, 46, 345–351. [Google Scholar] [CrossRef]
- Willi, J.; Kupfer, P.; Evequoz, D.; Fernandez, G.; Katz, A.; Leumann, C.; Polacek, N. Oxidative stress damages rRNA inside the ribosome and differentially affects the catalytic center. Nucleic Acids Res. 2018, 46, 1945–1957. [Google Scholar] [CrossRef]
- Blekas, G.; Vassilakis, C.; Harizanis, C.; Tsimidou, M.; Boskou, D.G. Biophenols in table olives. J. Agric. Food Chem. 2002, 50, 3688–3692. [Google Scholar] [CrossRef]
- Vilaplana-Perez, C.; Aunon, D.; Garcia-Flores, L.A.; Gil-Izquierdo, A. Hydroxytyrosol and potential uses in cardiovascular diseases, cancer, and AIDS. Front. Nutr. 2014, 1, 18. [Google Scholar] [CrossRef]
- Parkinson, L.; Cicerale, S. The Health Benefiting Mechanisms of Virgin Olive Oil Phenolic Compounds. Molecules 2016, 21, 1734. [Google Scholar] [CrossRef]
- Fuccelli, R.; Fabiani, R.; Rosignoli, P. Hydroxytyrosol Exerts Anti-Inflammatory and Anti-Oxidant Activities in a Mouse Model of Systemic Inflammation. Molecules 2018, 23, 3212. [Google Scholar] [CrossRef]
- Larussa, T.; Imeneo, M.; Luzza, F. Olive Tree Biophenols in Inflammatory Bowel Disease: When Bitter is Better. Int. J. Mol. Sci. 2019, 20, 1390. [Google Scholar] [CrossRef]
- Amini, A.; Liu, M.; Ahmad, Z. Understanding the link between antimicrobial properties of dietary olive phenolics and bacterial ATP synthase. Int. J. Biol. Macromol. 2017, 101, 153–164. [Google Scholar] [CrossRef]
- Furneri, P.M.; Piperno, A.; Sajia, A.; Bisignano, G. Antimycoplasmal activity of hydroxytyrosol. Antimicrob. Agents Chemother. 2004, 48, 4892–4894. [Google Scholar] [CrossRef] [PubMed]
- Belaqziz, M.; Tan, S.P.; El-Abbassi, A.; Kiai, H.; Hafidi, A.; O’Donovan, O.; McLoughlin, P. Assessment of the antioxidant and antibacterial activities of different olive processing wastewaters. PLoS ONE 2017, 12, e0182622. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Huelamo, M.; Rodriguez-Morato, J.; Boronat, A.; de la Torre, R. Modulation of Nrf2 by Olive Oil and Wine Polyphenols and Neuroprotection. Antioxidants 2017, 6, 73. [Google Scholar] [CrossRef]
- Fukai, T.; Ushio-Fukai, M. Superoxide dismutases: Role in redox signaling, vascular function, and diseases. Antioxid. Redox Signal. 2011, 15, 1583–1606. [Google Scholar] [CrossRef] [PubMed]
- Swanson, A.M.; David, A.L. Animal models of fetal growth restriction: Considerations for translational medicine. Placenta 2015, 36, 623–630. [Google Scholar] [CrossRef]
- Bahr, A.; Wolf, E. Domestic animal models for biomedical research. Reprod. Domest. Anim. 2012, 47 (Suppl. 4), 59–71. [Google Scholar] [CrossRef]
- Cibelli, J.; Emborg, M.E.; Prockop, D.J.; Roberts, M.; Schatten, G.; Rao, M.; Harding, J.; Mirochnitchenko, O. Strategies for improving animal models for regenerative medicine. Cell Stem Cell 2013, 12, 271–274. [Google Scholar] [CrossRef]
- Bonney, E.A. Demystifying animal models of adverse pregnancy outcomes: Touching bench and bedside. Am. J. Reprod. Immunol. 2013, 69, 567–584. [Google Scholar] [CrossRef]
- Sharifi, T.; Ghayeb, Y. A computational study to identify the key residues of peroxisome proliferator-activated receptor gamma in the interactions with its antagonists. J. Biomol. Struct. Dyn. 2018, 36, 1822–1833. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.-C.; Zhang, C.; Jin, Q.; Wei, C.; Zhao, H.-B.; Zhang, X.-L.; You, W.; Liu, X.-M.; Liu, G.-F.; Liu, Y.-F. Protective effects of astaxanthin on lipopolysaccharide-induced inflammation in bovine endometrial epithelial cells. Biol. Reprod. 2020, 102, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.I.; Kim, T.G.; Lim, D.H.; Kim, S.B.; Park, S.M.; Hur, T.Y.; Ki, K.S.; Kwon, E.G.; Vijayakumar, M.; Kim, Y.J. Preparation of Nanoemulsions of Vitamin A and C by Microfluidization: Efficacy on the Expression Pattern of Milk-Specific Proteins in MAC-T Cells. Molecules 2019, 24, 2566. [Google Scholar] [CrossRef] [PubMed]
- Pallio, G.; Bitto, A.; Pizzino, G.; Galfo, F.; Irrera, N.; Squadrito, F.; Squadrito, G.; Pallio, S.; Anastasi, G.P.; Cutroneo, G.; et al. Adenosine Receptor Stimulation by Polydeoxyribonucleotide Improves Tissue Repair and Symptomology in Experimental Colitis. Front. Pharmacol. 2016, 7, 273. [Google Scholar] [CrossRef] [PubMed]
- Squadrito, F.; Micali, A.; Rinaldi, M.; Irrera, N.; Marini, H.; Puzzolo, D.; Pisani, A.; Lorenzini, C.; Valenti, A.; Laura, R.; et al. Polydeoxyribonucleotide, an Adenosine-A2A Receptor Agonist, Preserves Blood Testis Barrier from Cadmium-Induced Injury. Front Pharmacol 2016, 7, 537. [Google Scholar] [CrossRef]
- Kim, Y.C.; Kim, B.G.; Lee, J.H. Thymosin beta10 expression driven by the human TERT promoter induces ovarian cancer-specific apoptosis through ROS production. PLoS ONE 2012, 7, e35399. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Bitto, A.; Pallio, G.; Mannino, F.; Arcoraci, V.; Aliquo, F.; Minutoli, L.; De Ponte, C.; D’Andrea, P.; et al. Cadmium-Induced Oxidative Stress Impairs Glycemic Control in Adolescents. Oxid. Med. Cell. Longev. 2017, 2017, 6341671. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Cronin, J.G.; Bromfield, J.J. Tolerance and Innate Immunity Shape the Development of Postpartum Uterine Disease and the Impact of Endometritis in Dairy Cattle. Annu. Rev. Anim. Biosci. 2019, 7, 361–384. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Cronin, J.; Goetze, L.; Donofrio, G.; Schuberth, H.J. Defining postpartum uterine disease and the mechanisms of infection and immunity in the female reproductive tract in cattle. Biol. Reprod. 2009, 81, 1025–1032. [Google Scholar] [CrossRef]
- Swangchan-Uthai, T.; Lavender, C.R.; Cheng, Z.; Fouladi-Nashta, A.A.; Wathes, D.C. Time course of defense mechanisms in bovine endometrium in response to lipopolysaccharide. Biol. Reprod. 2012, 87, 135. [Google Scholar] [CrossRef]
- Castillo, C.; Hernandez, J.; Bravo, A.; Lopez-Alonso, M.; Pereira, V.; Benedito, J.L. Oxidative status during late pregnancy and early lactation in dairy cows. Vet. J. 2005, 169, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, L.; Zhang, C.; Cheng, X.; Zhang, Y.; Guo, Y.; Long, M.; Yang, S.; He, J. Palmitic Acid and beta-Hydroxybutyrate Induce Inflammatory Responses in Bovine Endometrial Cells by Activating Oxidative Stress-Mediated NF-kappaB Signaling. Molecules 2019, 24, 2421. [Google Scholar] [CrossRef]
- Mandhwani, R.; Bhardwaz, A.; Kumar, S.; Shivhare, M.; Aich, R. Insights into bovine endometritis with special reference to phytotherapy. Vet. World 2017, 10, 1529–1532. [Google Scholar] [CrossRef][Green Version]
- Sharma, P.; Srivastava, S.; Kumar, R.; Singh, V. Phytotherapy: An Alternative Low Cost Therapeutic Management of Endometritis in Dairy Animals: A Review. Int. J. Curr. Microbiol. App. Sci 2018, 7, 4581–4591. [Google Scholar]
- Bhardwaz, A.; Nema, S.; Mahour, S.; Chabbra, D.; Rajput, N.; Sudarshan, K. Effect of Garlic (Allium sativum) Extract on Recovery and Conception Rate in Infectious Repeat Breeder Crossbred Cows. Indian J. Vet. Sci. Biotechnol. 2018, 14, 60–63. [Google Scholar] [CrossRef]
- Rathaur, P.; Raja, W.; Ramteke, P.; John, S.A. Turmeric: The golden spice of life. Int. J. Pharm. Sci. Res. 2012, 3, 1987. [Google Scholar]
- Karkovic Markovic, A.; Toric, J.; Barbaric, M.; Jakobusic Brala, C. Hydroxytyrosol, Tyrosol and Derivatives and Their Potential Effects on Human Health. Molecules 2019, 24, 2001. [Google Scholar] [CrossRef]
- Fusco, R.; Cordaro, M.; Siracusa, R.; Peritore, A.F.; D’Amico, R.; Licata, P.; Crupi, R.; Gugliandolo, E. Effects of Hydroxytyrosol against Lipopolysaccharide-Induced Inflammation and Oxidative Stress in Bovine Mammary Epithelial Cells: A Natural Therapeutic Tool for Bovine Mastitis. Antioxidants 2020, 9, 693. [Google Scholar] [CrossRef]
- Basirico, L.; Morera, P.; Dipasquale, D.; Bernini, R.; Santi, L.; Romani, A.; Lacetera, N.; Bernabucci, U. (-)-Epigallocatechin-3-gallate and hydroxytyrosol improved antioxidative and anti-inflammatory responses in bovine mammary epithelial cells. Animal 2019, 13, 2847–2856. [Google Scholar] [CrossRef]
- Goldsmith, C.D.; Bond, D.R.; Jankowski, H.; Weidenhofer, J.; Stathopoulos, C.E.; Roach, P.D.; Scarlett, C.J. The Olive Biophenols Oleuropein and Hydroxytyrosol Selectively Reduce Proliferation, Influence the Cell Cycle, and Induce Apoptosis in Pancreatic Cancer Cells. Int. J. Mol. Sci. 2018, 19, 1937. [Google Scholar] [CrossRef]
- Chase, C.; Kaushik, R.S. Mucosal Immune System of Cattle: All Immune Responses Begin Here. Vet Clin. N. Am. Food Anim. Pract. 2019, 35, 431–451. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| LPS -/HT - | |
|---|---|
| Ctrl | 100 ± 0 |
| HT 10 μM | 98.6 ± 0.50 |
| HT 25 μM | 98.4± 0.67 |
| HT 50 μM | 97.8 ± 0.86 |
| HT 100 μM | 94.6 ± 1.20# |
| HT 250 μM | 93.6 ± 1.56## |
| LPS -/HT - | LPS +/HT- | LPS +/HT 10 μM | LPS +/HT 25 μM | |
|---|---|---|---|---|
| ROS | 16.6 ± 0.02 | 33.8 ± 1.68 *** | 22.4 ± 1.20 ## | 15.6 ± 1.07 ### |
| TNF-α | 252.8 ± 10.8 | 304.8 ± 7.41 ** | 241.8 ± 5.2 # | 237 ± 7.36 ### |
| IL-6 | 1.70 ± 0.1 | 4.30 ± 120.8 ** | 1.50 ± 0.11 ## | 1.32 ± 0.16 ### |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gugliandolo, E.; Fusco, R.; Licata, P.; Peritore, A.F.; D’amico, R.; Cordaro, M.; Siracusa, R.; Cuzzocrea, S.; Crupi, R. Protective Effect of Hydroxytyrosol on LPS-Induced Inflammation and Oxidative Stress in Bovine Endometrial Epithelial Cell Line. Vet. Sci. 2020, 7, 161. https://doi.org/10.3390/vetsci7040161
Gugliandolo E, Fusco R, Licata P, Peritore AF, D’amico R, Cordaro M, Siracusa R, Cuzzocrea S, Crupi R. Protective Effect of Hydroxytyrosol on LPS-Induced Inflammation and Oxidative Stress in Bovine Endometrial Epithelial Cell Line. Veterinary Sciences. 2020; 7(4):161. https://doi.org/10.3390/vetsci7040161
Chicago/Turabian StyleGugliandolo, Enrico, Roberta Fusco, Patrizia Licata, Alessio Filippo Peritore, Ramona D’amico, Marika Cordaro, Rosalba Siracusa, Salvatore Cuzzocrea, and Rosalia Crupi. 2020. "Protective Effect of Hydroxytyrosol on LPS-Induced Inflammation and Oxidative Stress in Bovine Endometrial Epithelial Cell Line" Veterinary Sciences 7, no. 4: 161. https://doi.org/10.3390/vetsci7040161
APA StyleGugliandolo, E., Fusco, R., Licata, P., Peritore, A. F., D’amico, R., Cordaro, M., Siracusa, R., Cuzzocrea, S., & Crupi, R. (2020). Protective Effect of Hydroxytyrosol on LPS-Induced Inflammation and Oxidative Stress in Bovine Endometrial Epithelial Cell Line. Veterinary Sciences, 7(4), 161. https://doi.org/10.3390/vetsci7040161