Involvement of Various Enzymes in the Physiology and Pathogenesis of Streptococcus suis


Abstract
1. Introduction
2. ATPases
3. Immunoglobulin-Degrading Enzymes
4. Eukaryote-Like Serine/Threonine Kinase and Phosphatase
5. Subtilisin-Like Serine Proteases
6. Superoxide Dismutase and NADH Oxidase
7. Nucleases
8. Enolase
9. S-ribosylhomocysteinase (LuxS)
10. Peptidyl Isomerase PrsA
11. (p)ppGpp Synthetases
12. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Goyette-Desjardins, G.; Auger, J.P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing. Emerg. Microbes Infect. 2014, 3, e45. [Google Scholar] [CrossRef] [PubMed]
- Lun, Z.R.; Wang, Q.P.; Chen, X.G.; Li, A.X.; Zhu, X.Q. Streptococcus suis: An emerging zoonotic pathogen. Lancet Infect. Dis. 2007, 7, 201–209. [Google Scholar] [CrossRef]
- Zhang, B.; Ku, X.; Yu, X.; Sun, Q.; Wu, H.; Chen, F.; Zhang, X.; Guo, L.; Tang, X.; He, Q. Prevalence and antimicrobial susceptibilities of bacterial pathogens in Chinese pig farms from 2013 to 2017. Sci. Rep. 2019, 9, 9908. [Google Scholar] [CrossRef] [PubMed]
- Wertheim, H.F.; Nghia, H.D.; Taylor, W.; Schultsz, C. Streptococcus suis: An emerging human pathogen. Clin. Infect. Dis. 2009, 48, 617–625. [Google Scholar] [CrossRef]
- Tang, J.; Wang, C.; Feng, Y.; Yang, W.; Song, H.; Chen, Z.; Yu, H.; Pan, X.; Zhou, X.; Wang, H.; et al. Streptococcal toxic shock syndrome caused by Streptococcus suis serotype 2. PLoS Med. 2006, 3, e151. [Google Scholar]
- Yu, H.; Jing, H.; Chen, Z.; Zheng, H.; Zhu, X.; Wang, H.; Wang, S.; Liu, L.; Zu, R.; Luo, L.; et al. Human Streptococcus suis outbreak, Sichuan, China. Emerg. Infect. Dis. 2006, 12, 914–920. [Google Scholar] [CrossRef]
- Jiang, F.; Guo, J.; Cheng, C.; Gu, B. Human infection caused by Streptococcus suis serotype 2 in China: Report of two cases and epidemic distribution based on sequence type. BMC Infect. Dis. 2020, 20, 223. [Google Scholar] [CrossRef]
- Liu, S.S.; Wang, Y.; Xue, L.; Ma, C.; Li, C.H. Hemophagocytic lymphohistiocytosis due to Streptococcus suis in a 12-year-old girl A case report. Medicine 2019, 98, e15136. [Google Scholar] [CrossRef]
- Rayanakorn, A.; Katip, W.; Lee, L.H.; Oberdorfer, P. Endophthalmitis with bilateral deafness from disseminated Streptococcus suis infection. BMJ Case Rep. 2019, 12, e228501. [Google Scholar] [CrossRef]
- Nemeth, A.; Knausz, M.; Schmidt, P. Special case of purulent meningitis caused by Streptococcus suis. Case report. Orv. Hetil. 2019, 160, 30–34. [Google Scholar]
- Yanase, T.; Morii, D.; Kamio, S.; Nishimura, A.; Fukao, E.; Inose, Y.; Honma, Y.; Kitahara, N.; Yokozawa, T.; Chang, B.; et al. The first report of human meningitis and pyogenic ventriculitis caused by Streptococcus suis: A case report. J. Infect. Chemother. 2018, 24, 669–673. [Google Scholar] [CrossRef]
- Segura, M.; Fittipaldi, N.; Calzas, C.; Gottschalk, M. Critical Streptococcus suis virulence factors: Are they all really critical? Trends Microbiol. 2017, 25, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Higgins, R.; Gottschalk, M.; Boudreau, M.; Lebrun, A.; Henrichsen, J. Description of six new capsular types (29-34) of Streptococcus suis. J. Vet. Diagn. Investig. 1995, 7, 405–406. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.E.; Gottschalk, M.; Brousseau, R.; Harel, J.; Hemmingsen, S.M.; Goh, S.H. Biochemical analysis, cpn60 and 16S rDNA sequence data indicate that Streptococcus suis serotypes 32 and 34, isolated from pigs, are Streptococcus orisratti. Vet. Microbiol. 2005, 107, 63–69. [Google Scholar] [CrossRef]
- Tien le, H.T.; Nishibori, T.; Nishitani, Y.; Nomoto, R.; Osawa, R. Reappraisal of the taxonomy of Streptococcus suis serotypes 20, 22, 26, and 33 based on DNA-DNA homology and sodA and recN phylogenies. Vet. Microbiol. 2013, 162, 842–849. [Google Scholar] [CrossRef]
- Nomoto, R.; Maruyama, F.; Ishida, S.; Tohya, M.; Sekizaki, T.; Osawa, R. Reappraisal of the taxonomy of Streptococcus suis serotypes 20, 22 and 26: Streptococcus parasuis sp. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Tohya, M.; Arai, S.; Tomida, J.; Watanabe, T.; Kawamura, Y.; Katsumi, M.; Ushimizu, M.; Ishida-Kuroki, K.; Yoshizumi, M.; Uzawa, Y.; et al. Defining the taxonomic status of Streptococcus suis serotype 33: The proposal for Streptococcus ruminantium sp. nov. Int. J. Syst. Evol. Microbiol. 2017, 67, 3660–3665. [Google Scholar] [CrossRef]
- Segura, M.; Calzas, C.; Grenier, D.; Gottschalk, M. Initial steps of the pathogenesis of the infection caused by Streptococcus suis: Fighting against nonspecific defenses. FEBS Lett. 2016, 590, 3772–3799. [Google Scholar]
- Fittipaldi, N.; Segura, M.; Grenier, D.; Gottschalk, M. Virulence factors involved in the pathogenesis of the infection caused by the swine pathogen and zoonotic agent Streptococcus suis. Future Microbiol 2012, 7, 259–279. [Google Scholar]
- Zheng, C.; Jia, M.; Lu, T.; Gao, M.; Li, L. CopA Protects Streptococcus suis against copper toxicity. Int. J. Mol. Sci. 2019, 20, 2969. [Google Scholar] [CrossRef]
- Zheng, C.; Jia, M.; Gao, M.; Lu, T.; Li, L.; Zhou, P. PmtA functions as a ferrous iron and cobalt efflux pump in Streptococcus suis. Emerg. Microbes Infect. 2019, 8, 1254–1264. [Google Scholar] [CrossRef]
- Tan, M.F.; Gao, T.; Liu, W.Q.; Zhang, C.Y.; Yang, X.; Zhu, J.W.; Teng, M.Y.; Li, L.; Zhou, R. MsmK, an ATPase, contributes to utilization of multiple carbohydrates and host colonization of Streptococcus suis. PLoS ONE 2015, 10, e0130792. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.F.; Liu, W.Q.; Zhang, C.Y.; Gao, T.; Zheng, L.L.; Qiu, D.X.; Li, L.; Zhou, R. The involvement of MsmK in pathogenesis of the Streptococcus suis serotype 2. Microbiologyopen 2017, 6, e00433. [Google Scholar] [CrossRef] [PubMed]
- Seele, J.; Singpiel, A.; Spoerry, C.; von Pawel-Rammingen, U.; Valentin-Weigand, P.; Baums, C.G. Identification of a novel host-specific IgM protease in Streptococcus suis. J. Bacteriol. 2013, 195, 930–940. [Google Scholar] [CrossRef] [PubMed]
- Seele, J.; Beineke, A.; Hillermann, L.M.; Jaschok-Kentner, B.; von Pawel-Rammingen, U.; Valentin-Weigand, P.; Baums, C.G. The immunoglobulin M-degrading enzyme of Streptococcus suis, IdeSsuis, is involved in complement evasion. Vet. Res. 2015, 46, 45. [Google Scholar] [CrossRef] [PubMed]
- Rungelrath, V.; Weisse, C.; Schutze, N.; Muller, U.; Meurer, M.; Rohde, M.; Seele, J.; Valentin-Weigand, P.; Kirschfink, M.; Beineke, A.; et al. IgM cleavage by Streptococcus suis reduces IgM bound to the bacterial surface and is a novel complement evasion mechanism. Virulence 2018, 9, 1314–1337. [Google Scholar] [CrossRef]
- Spoerry, C.; Seele, J.; Valentin-Weigand, P.; Baums, C.G.; von Pawel-Rammingen, U. Identification and characterization of IgdE, a novel IgG-degrading protease of Streptococcus suis with unique specificity for porcine IgG. J. Biol. Chem. 2016, 291, 7915–7925. [Google Scholar] [CrossRef]
- Zhang, A.; Mu, X.; Chen, B.; Liu, C.; Han, L.; Chen, H.; Jin, M. Identification and characterization of IgA1 protease from Streptococcus suis. Vet. Microbiol. 2010, 140, 171–175. [Google Scholar] [CrossRef]
- Zhang, A.D.; Mu, X.F.; Chen, B.; Han, L.; Chen, H.C.; Jin, M.L. IgA1 protease contributes to the virulence of Streptococcus suis. Vet. Microbiol. 2011, 148, 436–439. [Google Scholar] [CrossRef]
- Bek-Thomsen, M.; Poulsen, K.; Kilian, M. Occurrence and evolution of the paralogous zinc metalloproteases IgA1 protease, ZmpB, ZmpC, and ZmpD in Streptococcus pneumoniae and related commensal species. mBio 2012, 3, e00303-12. [Google Scholar] [CrossRef]
- Dumesnil, A.; Auger, J.P.; Roy, D.; Votsch, D.; Willenborg, M.; Valentin-Weigand, P.; Park, P.W.; Grenier, D.; Fittipaldi, N.; Harel, J.; et al. Characterization of the zinc metalloprotease of Streptococcus suis serotype 2. Vet. Res. 2018, 49, 109. [Google Scholar]
- Zhu, H.; Zhou, J.; Ni, Y.; Yu, Z.; Mao, A.; Hu, Y.; Wang, W.; Zhang, X.; Wen, L.; Li, B.; et al. Contribution of eukaryotic-type serine/threonine kinase to stress response and virulence of Streptococcus suis. PLoS ONE 2014, 9, e91971. [Google Scholar]
- Zhang, C.; Sun, W.; Tan, M.; Dong, M.; Liu, W.; Gao, T.; Li, L.; Xu, Z.; Zhou, R. The eukaryote-like serine/threonine kinase STK regulates the growth and metabolism of zoonotic Streptococcus suis. Front. Cell Infect. Microbiol. 2017, 7, 66. [Google Scholar]
- Rui, L.; Weiyi, L.; Yu, M.; Hong, Z.; Jiao, Y.; Zhe, M.; Hongjie, F. The serine/threonine protein kinase of Streptococcus suis serotype 2 affects the ability of the pathogen to penetrate the blood-brain barrier. Cell Microbiol. 2018, 20, e12862. [Google Scholar] [PubMed]
- Zhu, H.; Huang, D.; Zhang, W.; Wu, Z.; Lu, Y.; Jia, H.; Wang, M.; Lu, C. The novel virulence-related gene stp of Streptococcus suis serotype 9 strain contributes to a significant reduction in mouse mortality. Microb. Pathog. 2011, 51, 442–453. [Google Scholar] [PubMed]
- Fang, L.; Zhou, J.; Fan, P.; Yang, Y.; Shen, H.; Fang, W. A serine/threonine phosphatase 1 of Streptococcus suis type 2 is an important virulence factor. J. Vet. Sci. 2017, 18, 439–447. [Google Scholar] [PubMed]
- Hu, Q.; Liu, P.; Yu, Z.; Zhao, G.; Li, J.; Teng, L.; Zhou, M.; Bei, W.; Chen, H.; Jin, M. Identification of a cell wall-associated subtilisin-like serine protease involved in the pathogenesis of Streptococcus suis serotype 2. Microb. Pathog. 2010, 48, 103–109. [Google Scholar]
- Yin, S.; Li, M.; Rao, X.; Yao, X.; Zhong, Q.; Wang, M.; Wang, J.; Peng, Y.; Tang, J.; Hu, F.; et al. Subtilisin-like protease-1 secreted through type IV secretion system contributes to high virulence of Streptococcus suis 2. Sci. Rep. 2016, 6, 27369. [Google Scholar]
- Bonifait, L.; de la Cruz Dominguez-Punaro, M.; Vaillancourt, K.; Bart, C.; Slater, J.; Frenette, M.; Gottschalk, M.; Grenier, D. The cell envelope subtilisin-like proteinase is a virulence determinant for Streptococcus suis. BMC Microbiol. 2010, 10, 42. [Google Scholar]
- Bonifait, L.; Vaillancourt, K.; Gottschalk, M.; Frenette, M.; Grenier, D. Purification and characterization of the subtilisin-like protease of Streptococcus suis that contributes to its virulence. Vet. Microbiol. 2011, 148, 333–340. [Google Scholar]
- Bonifait, L.; Grenier, D. The SspA subtilisin-like protease of Streptococcus suis triggers a pro-inflammatory response in macrophages through a non-proteolytic mechanism. BMC Microbiol. 2011, 11, 47. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.L.; Zhang, X.Y.; Wu, W.; Lu, Z.Y.; Fang, W.H. Inactivation of the sodA gene of Streptococcus suis type 2 encoding superoxide dismutase leads to reduced virulence to mice. Vet. Microbiol. 2012, 158, 360–366. [Google Scholar] [CrossRef]
- Fang, L.H.; Shen, H.X.; Tang, Y.L.; Fang, W.H. Superoxide dismutase of Streptococcus suis serotype 2 plays a role in anti-autophagic response by scavenging reactive oxygen species in infected macrophages. Vet. Microbiol. 2015, 176, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Ren, S.; Xu, J.; Zhao, X.; Shi, G.; Wu, J.; Li, J.; Chen, H.; Bei, W. Contribution of NADH oxidase to oxidative stress tolerance and virulence of Streptococcus suis serotype 2. Virulence 2017, 8, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Arenas, J.; Bossers-de Vries, R.; Harders-Westerveen, J.; Buys, H.; Ruuls-van Stalle, L.M.F.; Stockhofe-Zurwieden, N.; Zaccaria, E.; Tommassen, J.; Wells, J.M.; Smith, H.E.; et al. In vivo transcriptomes of Streptococcus suis reveal genes required for niche-specific adaptation and pathogenesis. Virulence 2019, 10, 334–351. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.; Bonifait, L.; Vaillancourt, K.; Charette, S.J.; Gottschalk, M.; Grenier, D. Characterization of DNase activity and gene in Streptococcus suis and evidence for a role as virulence factor. BMC Res. Notes 2014, 7, 424. [Google Scholar] [CrossRef]
- De Buhr, N.; Neumann, A.; Jerjomiceva, N.; von Kockritz-Blickwede, M.; Baums, C.G. Streptococcus suis DNase SsnA contributes to degradation of neutrophil extracellular traps (NETs) and evasion of NET-mediated antimicrobial activity. Microbiology 2014, 160, 385–395. [Google Scholar] [CrossRef]
- Li, M.; Cai, R.J.; Li, C.L.; Song, S.; Li, Y.; Jiang, Z.Y.; Yang, D.X. Deletion of ssnA attenuates the pathogenicity of Streptococcus suis and confers protection against serovar 2 strain challenge. PLoS ONE 2017, 12, e0169791. [Google Scholar] [CrossRef]
- De Buhr, N.; Stehr, M.; Neumann, A.; Naim, H.Y.; Valentin-Weigand, P.; von Kockritz-Blickwede, M.; Baums, C.G. Identification of a novel DNase of Streptococcus suis (EndAsuis) important for neutrophil extracellular trap degradation during exponential growth. Microbiology 2015, 161, 838–850. [Google Scholar] [CrossRef]
- Esgleas, M.; Li, Y.Y.; Hancock, M.A.; Harel, J.; Dubreuil, J.D.; Gottschalk, M. Isolation and characterization of alpha-enolase, a novel fibronectin-binding protein from Streptococcus suis. Microbiol-SGM 2008, 154, 2668–2679. [Google Scholar] [CrossRef]
- Lu, Q.; Lu, H.; Qi, J.X.; Lu, G.W.; Gao, G.F. An octamer of enolase from Streptococcus suis. Protein Cell 2012, 3, 769–780. [Google Scholar] [CrossRef] [PubMed]
- Pian, Y.Y.; Wang, P.P.; Liu, P.; Zheng, Y.L.; Zhu, L.; Wang, H.L.; Xu, B.; Yuan, Y.; Jiang, Y.Q. Proteomics identification of novel fibrinogen-binding proteins of Streptococcus suis contributing to antiphagocytosis. Front. Cell Infect. Microbiol. 2015, 5, 19. [Google Scholar] [CrossRef]
- Li, Q.; Liu, H.Z.; Du, D.C.; Yu, Y.F.; Ma, C.F.; Jiao, F.F.; Yao, H.C.; Lu, C.P.; Zhang, W. Identification of novel laminin- and fibronectin-binding proteins by far-western blot: Capturing the adhesins of Streptococcus suis type 2. Front. Cell Infect. Microbiol. 2015, 5, 82. [Google Scholar] [PubMed]
- Chen, B.; Zhang, A.; Xu, Z.; Li, R.; Chen, H.; Jin, M. Large-scale identification of bacteria-host crosstalk by affinity chromatography: Capturing the interactions of Streptococcus suis proteins with host cells. J. Proteome Res. 2011, 10, 5163–5174. [Google Scholar] [CrossRef]
- Zhang, A.; Chen, B.; Mu, X.; Li, R.; Zheng, P.; Zhao, Y.; Chen, H.; Jin, M. Identification and characterization of a novel protective antigen, enolase of Streptococcus suis serotype 2. Vaccine 2009, 27, 1348–1353. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.J.; Pan, X.Z.; Sun, W.; Wang, C.J.; Zhang, H.M.; Li, X.F.; Ma, Y.; Shao, Z.Q.; Ge, J.C.; Zheng, F.; et al. Streptococcus suis enolase functions as a protective antigen displayed on the bacterial cell surface. J. Infect. Dis. 2009, 200, 1583–1592. [Google Scholar] [CrossRef]
- Sun, Y.Y.; Li, N.; Zhang, J.; Liu, H.T.; Liu, J.F.; Xia, X.J.; Sun, C.J.; Feng, X.; Gu, J.M.; Du, C.T.; et al. Enolase of Streptococcus suis serotype 2 enhances blood-brain barrier permeability by inducing IL-8 release. Inflammation 2016, 39, 718–726. [Google Scholar] [CrossRef]
- Li, Q.; Fu, Y.; Guo, G.; Wang, Z.; Zhang, W. Identification of novel pig and human immunoglobulin G-binding proteins and characterization of the binding regions of enolase from Streptococcus suis serotype 2. AMB Express 2020, 10, 103. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, W.; Wu, Z.; Zhu, X.; Lu, C. Functional analysis of luxS in Streptococcus suis reveals a key role in biofilm formation and virulence. Vet. Microbiol. 2011, 152, 151–160. [Google Scholar] [CrossRef]
- Cao, M.; Feng, Y.; Wang, C.; Zheng, F.; Li, M.; Liao, H.; Mao, Y.; Pan, X.; Wang, J.; Hu, D.; et al. Functional definition of LuxS, an autoinducer-2 (AI-2) synthase and its role in full virulence of Streptococcus suis serotype 2. J. Microbiol. 2011, 49, 1000–1011. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, B.; Li, J.; Gong, S.; Dong, X.; Mao, C.; Yi, L. LuxS/AI-2 system is involved in fluoroquinolones susceptibility in Streptococcus suis through overexpression of efflux pump SatAB. Vet. Microbiol. 2019, 233, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Yang, Y.; Zhou, J.; Zhu, L.; Gu, Y.; Zhang, X.; Li, X.; Fang, W. Roles of the putative type IV-like secretion system key component VirD4 and PrsA in pathogenesis of Streptococcus suis type 2. Front. Cell Infect. Microbiol. 2016, 6, 172. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Yang, Y.; Zhou, J.; Liu, H.; Liao, X.; Luo, J.; Li, X.; Fang, W. Peptidyl isomerase PrsA is surface-associated on Streptococcus suis and offers cross-protection against serotype 9 strain. FEMS Microbiol. Lett. 2019, 366, fnz002. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fu, H.; Jiang, X.; Liao, X.; Yue, M.; Li, X.; Fang, W. PrsA contributes to Streptococcus suis serotype 2 pathogenicity by modulating secretion of selected virulence factors. Vet. Microbiol. 2019, 236, 108375. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhang, T.; Su, Z.; Li, L.; Wang, D.; Xiao, R.; Teng, M.; Tan, M.; Zhou, R. (p)ppGpp synthetases regulate the pathogenesis of zoonotic Streptococcus suis. Microbiol. Res. 2016, 191, 1–11. [Google Scholar] [CrossRef]
- Zhang, T.; Zhu, J.; Wei, S.; Luo, Q.; Li, L.; Li, S.; Tucker, A.; Shao, H.; Zhou, R. The roles of RelA/(p)ppGpp in glucose-starvation induced adaptive response in the zoonotic Streptococcus suis. Sci. Rep. 2016, 6, 27169. [Google Scholar] [CrossRef] [PubMed]
- Mitrakul, K.; Loo, C.Y.; Hughes, C.V.; Ganeshkumar, N. Role of a Streptococcus gordonii copper-transport operon, copYAZ, in biofilm detachment. Oral Microbiol. Immunol. 2004, 19, 395–402. [Google Scholar] [CrossRef]
- Solioz, M.; Abicht, H.K.; Mermod, M.; Mancini, S. Response of gram-positive bacteria to copper stress. J. Biol. Inorg. Chem. 2010, 15, 3–14. [Google Scholar] [CrossRef]
- Alquethamy, S.F.; Khorvash, M.; Pederick, V.G.; Whittall, J.J.; Paton, J.C.; Paulsen, I.T.; Hassan, K.A.; McDevitt, C.A.; Eijkelkamp, B.A. The Role of the CopA copper efflux system in Acinetobacter baumannii virulence. Int. J. Mol. Sci. 2019, 20, 575. [Google Scholar] [CrossRef]
- Shafeeq, S.; Yesilkaya, H.; Kloosterman, T.G.; Narayanan, G.; Wandel, M.; Andrew, P.W.; Kuipers, O.P.; Morrissey, J.A. The cop operon is required for copper homeostasis and contributes to virulence in Streptococcus pneumoniae. Mol. Microbiol. 2011, 81, 1255–1270. [Google Scholar] [CrossRef]
- Pi, H.L.; Patel, S.J.; Arguello, J.M.; Helmann, J.D. The Listeria monocytogenes Fur-regulated virulence protein FrvA is an Fe(II) efflux P1B4-type ATPase. Mol. Microbiol. 2016, 100, 1066–1079. [Google Scholar] [CrossRef] [PubMed]
- VanderWal, A.R.; Makthal, N.; Pinochet-Barros, A.; Helmann, J.D.; Olsen, R.J.; Kumaraswami, M. Iron efflux by PmtA is critical for oxidative stress resistance and contributes significantly to Group A Streptococcus virulence. Infect. Immun. 2017, 85, e00091-17. [Google Scholar] [CrossRef] [PubMed]
- Marion, C.; Aten, A.E.; Woodiga, S.A.; King, S.J. Identification of an ATPase, MsmK, which energizes multiple carbohydrate ABC transporters in Streptococcus pneumoniae. Infect. Immun. 2011, 79, 4193–4200. [Google Scholar] [CrossRef]
- Seele, J.; Hillermann, L.M.; Beineke, A.; Seitz, M.; von Pawel-Rammingen, U.; Valentin-Weigand, P.; Baums, C.G. The immunoglobulin M-degrading enzyme of Streptococcus suis, Ide(Ssuis), is a highly protective antigen against serotype 2. Vaccine 2015, 33, 2207–2212. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Zhao, J.Q.; Lin, L.; Zhang, Q.; Xu, Z.M.; Han, L.; Xie, C.Y.; Zhou, R.; Jin, M.L.; Zhang, A.D. Characterization of IgA1 protease as a surface protective antigen of Streptococcus suis serotype 2. Microbes Infect. 2016, 18, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Patenge, N.; Fiedler, T.; Kreikemeyer, B. Common regulators of virulence in streptococci. Curr. Top. Microbiol. Immunol. 2013, 368, 111–153. [Google Scholar]
- Ni, H.; Fan, W.W.; Li, C.L.; Wu, Q.Q.; Hongfen, H.F.; Hui, D.; Zheng, F.; Zhu, X.H.; Wang, C.J.; Cao, X.R.; et al. Streptococcus suis DivIVA protein is a substrate of Ser/Thr kinase STK and involved in cell division regulation. Front. Cell Infect. Microbiol. 2018, 8, 85. [Google Scholar] [CrossRef]
- Schatzman, S.S.; Culotta, V.C. Chemical warfare at the microorganismal level: A closer look at the superoxide dismutase enzymes of pathogens. ACS Infect. Dis. 2018, 4, 893–903. [Google Scholar] [CrossRef]
- Niven, D.F.; Ekins, A.; Al-Samaurai, A.A.W. Effects of iron and manganese availability on growth and production of superoxide dismutase by Streptococcus suis. Can. J. Microbiol. 1999, 45, 1027–1032. [Google Scholar] [CrossRef]
- Han, H.M.; Liu, C.H.; Wang, Q.H.; Xuan, C.L.; Zheng, B.W.; Tang, J.Q.; Yan, J.H.; Zhang, J.R.; Li, M.; Cheng, H.; et al. The two-component system Ihk/Irr contributes to the virulence of Streptococcus suis serotype 2 strain 05ZYH33 through alteration of the bacterial cell metabolism. Microbiol-SGM 2012, 158, 1852–1866. [Google Scholar] [CrossRef]
- Zheng, C.K.; Xu, J.L.; Li, J.Q.; Hu, L.H.; Xia, J.D.; Fan, J.Y.; Guo, W.N.; Chen, H.C.; Bei, W.C. Two Spx regulators modulate stress tolerance and virulence in Streptococcus suis serotype 2. PLoS ONE 2014, 9, e108197. [Google Scholar]
- Ge, X.C.; Yu, Y.; Zhang, M.; Chen, L.; Chen, W.H.; Elrami, F.; Kong, F.X.; Kitten, T.; Xu, P. Involvement of NADH oxidase in competition and endocarditis virulence in Streptococcus sanguinis. Infect. Immun. 2016, 84, 1470–1477. [Google Scholar] [PubMed]
- Higuchi, M.; Yamamoto, Y.; Poole, L.B.; Shimada, M.; Sato, Y.; Takahashi, N.; Kamio, Y. Functions of two types of NADH oxidases in energy metabolism and oxidative stress of Streptococcus mutans. J. Bacteriol. 1999, 181, 5940–5947. [Google Scholar] [PubMed]
- Zhao, G.; Zhang, H.; Chen, X.; Zhu, X.F.; Guo, Y.S.; He, C.F.; Khan, F.A.; Chen, Y.Y.; Hu, C.M.; Chen, H.C.; et al. Mycoplasma bovis NADH oxidase functions as both a NADH oxidizing and O2 reducing enzyme and an adhesin. Sci. Rep. 2017, 7, 44. [Google Scholar] [PubMed]
- Auzat, I.; Chapuy-Regaud, S.; Le Bras, G.; Dos Santos, D.; Ogunniyi, A.D.; Le Thomas, I.; Garel, J.R.; Paton, J.C.; Trombe, M.C. The NADH oxidase of Streptococcus pneumoniae: Its involvement in competence and virulence. Mol. Microbiol. 1999, 34, 1018–1028. [Google Scholar]
- Yamamoto, Y.; Pargade, V.; Lamberet, G.; Gaudu, P.; Thomas, F.; Texereau, J.; Gruss, A.; Trieu-Cuot, P.; Poyart, C. The Group B Streptococcus NADH oxidase Nox-2 is involved in fatty acid biosynthesis during aerobic growth and contributes to virulence. Mol. Microbiol. 2006, 62, 772–785. [Google Scholar]
- Fontaine, M.C.; Perez-Casal, J.; Willson, P.J. Investigation of a novel DNase of Streptococcus suis serotype 2. Infect. Immun. 2004, 72, 774–781. [Google Scholar]
- Wu, Z.; Zhang, W.; Lu, C. Comparative proteome analysis of secreted proteins of Streptococcus suis serotype 9 isolates from diseased and healthy pigs. Microbial. Pathog. 2008, 45, 159–166. [Google Scholar]
- Gomez-Gascon, L.; Cardoso-Toset, F.; Tarradas, C.; Gomez-Laguna, J.; Maldonado, A.; Nielsen, J.; Olaya-Abril, A.; Rodriguez-Ortega, M.J.; Luque, I. Characterization of the immune response and evaluation of the protective capacity of rSsnA against Streptococcus suis infection in pigs. Comp. Immunol. Microbiol. Infect. Dis. 2016, 47, 52–59. [Google Scholar]
- Gomez-Gascon, L.; Cardoso-Toset, F.; Amarilla, P.S.; Tarradas, C.; Carrasco, L.; Olaya-Abril, A.; Jimenez-Munguia, I.; Rodriguez-Ortega, M.J.; Luque, I. A new recombinant SsnA protein combined with aluminum hydroxide protects mouse against Streptococcus suis. Vaccine 2014, 32, 6992–6999. [Google Scholar]
- Esgleas, M.; Dominguez-Punaro, M.D.; Li, Y.Y.; Harel, J.; Dubreuil, J.D.; Gottschalk, M. Immunization with SsEno fails to protect mice against challenge with Streptococcus suis serotype 2. FEMS Microbiol. Lett. 2009, 294, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Dumesnil, A.; Martelet, L.; Grenier, D.; Auger, J.P.; Harel, J.; Nadeau, E.; Gottschalk, M. Enolase and dipeptidyl peptidase IV protein sub-unit vaccines are not protective against a lethal Streptococcus suis serotype 2 challenge in a mouse model of infection. BMC Vet. Res. 2019, 15, 448. [Google Scholar]
- Han, X.G.; Lu, C.P. Detection of autoinducer-2 and analysis of the profile of luxS and pfs transcription in Streptococcus suis serotype 2. Curr. Microbiol. 2009, 58, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yi, L.; Zhang, Z.; Fan, H.; Cheng, X.; Lu, C. Overexpression of luxS cannot increase autoinducer-2 production, only affect the growth and biofilm formation in Streptococcus suis. Sci. World J. 2013, 2013, 924276. [Google Scholar]
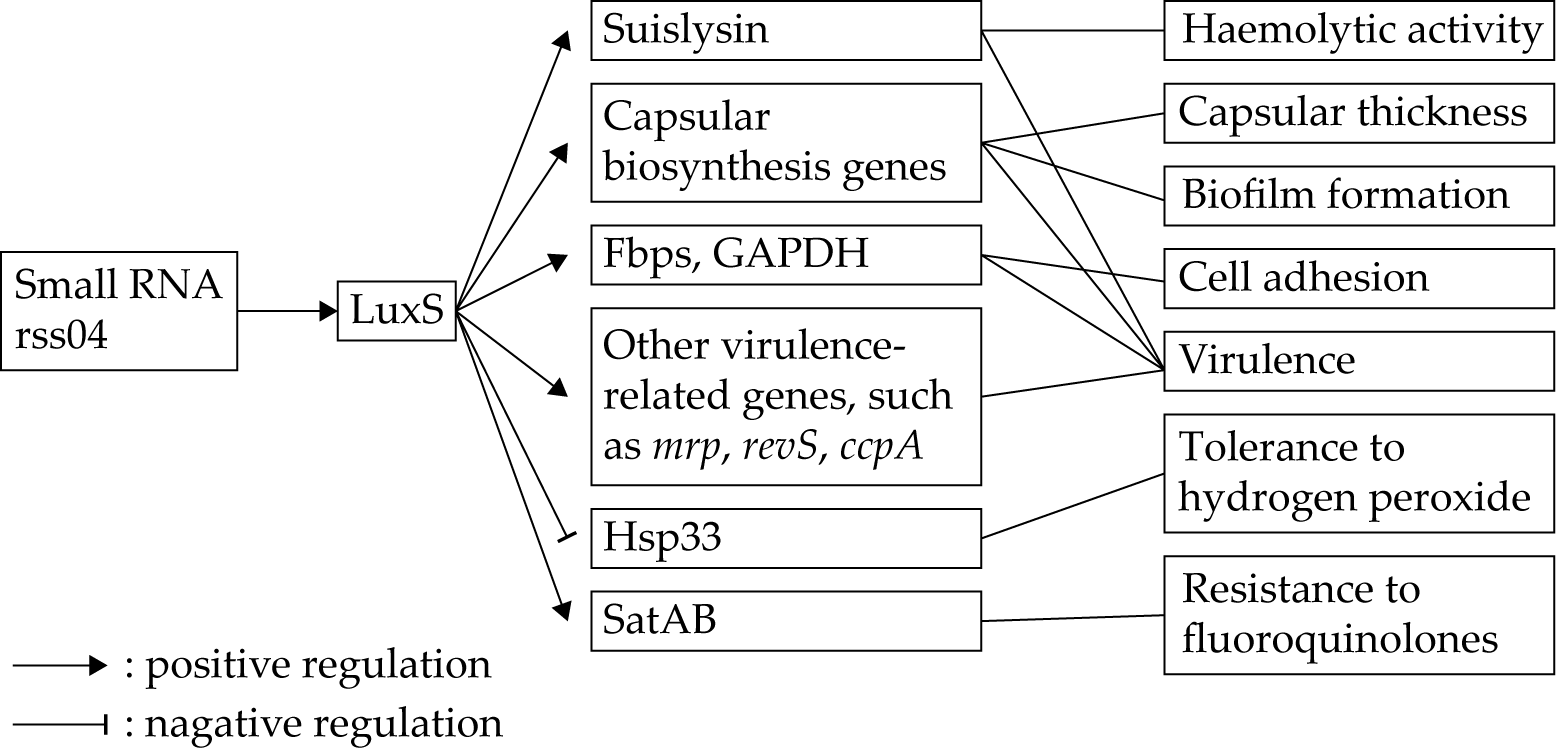
- Xiao, G.; Tang, H.; Zhang, S.; Ren, H.; Dai, J.; Lai, L.; Lu, C.; Yao, H.; Fan, H.; Wu, Z. Streptococcus suis small RNA rss04 contributes to the induction of meningitis by regulating capsule synthesis and by inducing biofilm formation in a mouse infection model. Vet. Microbiol. 2017, 199, 111–119. [Google Scholar] [CrossRef]
- Hauryliuk, V.; Atkinson, G.C.; Murakami, K.S.; Tenson, T.; Gerdes, K. Recent functional insights into the role of (p)ppGpp in bacterial physiology. Nat. Rev. Microbiol. 2015, 13, 298–309. [Google Scholar] [CrossRef]
- Zhu, J.; Zhang, T.; Su, Z.; Feng, L.; Liu, H.; Xu, Z.; Wu, Y.; Gao, T.; Shao, H.; Zhou, R. Co-regulation of CodY and (p)ppGpp synthetases on morphology and pathogenesis of Streptococcus suis. Microbiol. Res. 2019, 223–225, 88–98. [Google Scholar] [CrossRef]


{kind=link}
| Enzyme | Main Functions | References |
|---|---|---|
| CopA | Copper efflux | [20] |
| PmtA | Ferrous iron and cobalt efflux, tolerance to hydrogen peroxide-induced oxidative stress | [21] |
| MsmK | Utilization of multiple carbohydrates, pathogenesis | [22,23] |
| IdeSsuis | Degradation of porcine IgM, complement evasion | [24,25,26] |
| IgdE | Degradation of porcine IgG | [27] |
| IgA1 protease/ZmpC | Degradation of human IgA1, pathogenesis 1 | [28,29,30,31] |
| eSTK | Maintaining bacterial morphology, tolerance to stresses, pathogenesis | [32,33,34] |
| eSTP | Virulence, cell adhesion, and immune evasion 2 | [35,36] |
| SspA-1 | Virulence, trigger of proinflammatory cytokines | [37,38] |
| SspA-2 | Pathogenesis, proinflammatory response in macrophages | [39,40,41] |
| Superoxide dismutase | Oxidative stress resistance, virulence | [42,43] |
| NADH oxidase | Tolerance to oxidative stress, virulence | [44,45] |
| SsnA | Degradation of human and porcine neutrophil extracellular traps, pathogenesis | [46,47,48] |
| EndAsuis | Degradation of neutrophil extracellular traps | [49] |
| Enolase | Binding of extracellular matrix components, pathogenesis | [50,51,52,53,54,55,56,57,58] |
| LuxS | Growth, biofilm formation, capsule synthesis, hydrogen peroxide resistance, resistance to fluoroquinolones, pathogenesis | [59,60,61] |
| Peptidyl isomerase PrsA | Induction of proinflammatory cytokines, secretion of selected virulence factors, pathogenesis | [62,63,64] |
| (p)ppGpp synthetases (RelA, RelQ) | Pathogenesis, adaptation to glucose starvation (RelA) | [65,66] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, C.; Wei, M.; Jia, M.; Cao, M. Involvement of Various Enzymes in the Physiology and Pathogenesis of Streptococcus suis. Vet. Sci. 2020, 7, 143. https://doi.org/10.3390/vetsci7040143
Zheng C, Wei M, Jia M, Cao M. Involvement of Various Enzymes in the Physiology and Pathogenesis of Streptococcus suis. Veterinary Sciences. 2020; 7(4):143. https://doi.org/10.3390/vetsci7040143
Chicago/Turabian StyleZheng, Chengkun, Man Wei, Mengdie Jia, and ManMan Cao. 2020. "Involvement of Various Enzymes in the Physiology and Pathogenesis of Streptococcus suis" Veterinary Sciences 7, no. 4: 143. https://doi.org/10.3390/vetsci7040143
APA StyleZheng, C., Wei, M., Jia, M., & Cao, M. (2020). Involvement of Various Enzymes in the Physiology and Pathogenesis of Streptococcus suis. Veterinary Sciences, 7(4), 143. https://doi.org/10.3390/vetsci7040143