Avian Pattern Recognition Receptor Sensing and Signaling


Abstract
1. Introduction
2. Toll-Like Receptors
2.1. General Principles of Toll-Like Receptor Sensing and Activation
2.2. Avian TLRs
2.2.1. TLR1/2
2.2.2. TLR3
2.2.3. TLR4
2.2.4. TLR5
2.2.5. TLR7/8
2.2.6. TLR15
2.2.7. TLR21
3. NOD Leucine-Rich-Repeat Containing Protein Receptors (NLRs)
3.1. General Principles of NLR Activation and Inflammasome Assembly
3.2. Avian NLRs
3.2.1. NLRX1
3.2.2. NLRP3
3.2.3. NLRC3
3.2.4. NLRC5
4. RIG-I Like Receptors (RLRs)
4.1. RIG-I
4.2. MDA5
4.3. LGP-2
5. Additional RNA Sensors
5.1. OASL
5.2. PKR
5.3. DDX1, DDX21, and DHX36 Complex
5.4. DDX3
5.5. DDX23
5.6. DDX24
5.7. DDX60
5.8. Zinc Finger NFX1-Type Containing 1 (ZNFX1)
6. DNA and Cyclic Dinucleotide (CDN) Sensors
6.1. cGAS-STING DNA Sensing Pathway
6.1.1. cGAS
6.1.2. STING
6.1.3. ERADp
6.2. DDX41
6.3. DHX36
6.4. DHX9
6.5. hnRNPA2B1
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Keestra, A.M.; van Putten, J.P. Unique properties of the chicken TLR4/MD-2 complex: Selective lipopolysaccharide activation of the MyD88-dependent pathway. J. Immunol. 2008, 181, 4354–4362. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Cheng, A.; Wang, M. Innate sensing of viruses by pattern recognition receptors in birds. Vet. Res. 2013, 44, 82. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, R.; Chantrey, J.; Ganapathy, K. Immune Responses to Virulent and Vaccine Strains of Infectious Bronchitis Viruses in Chickens. Viral Immunol. 2015, 28, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Parvizi, P.; Abdul-Careem, M.F.; Haq, K.; Thanthrige-Don, N.; Schat, K.A.; Sharif, S. Immune responses against Marek’s disease virus. Anim. Health. Res. Rev. 2010, 11, 123–134. [Google Scholar] [CrossRef]
- Evseev, D.; Magor, K.E. Innate Immune Responses to Avian Influenza Viruses in Ducks and Chickens. Vet. Sci. 2019, 6, 5. [Google Scholar] [CrossRef]
- Coppo, M.J.C.; Hartley, C.A.; Devlin, J.M. Immune responses to infectious laryngotracheitis virus. Dev. Comp. Immunol. 2013, 41, 454–462. [Google Scholar] [CrossRef]
- Kapczynski, D.R.; Afonso, C.L.; Miller, P.J. Immune responses of poultry to Newcastle disease virus. Dev. Comp. Immunol. 2013, 41, 447–453. [Google Scholar] [CrossRef]
- Alkie, T.N.; Yitbarek, A.; Hodgins, D.C.; Kulkarni, R.R.; Taha-Abdelaziz, K.; Sharif, S. Development of innate immunity in chicken embryos and newly hatched chicks: A disease control perspective. Avian Pathol. 2019, 48, 288–310. [Google Scholar] [CrossRef]
- Kang, J.Y.; Lee, J.-O. Structural Biology of the Toll-Like Receptor Family. Annu. Rev. Biochem. 2011, 80, 917–941. [Google Scholar] [CrossRef]
- Carpenter, S.; O’Neill, L.A. Recent insights into the structure of Toll-like receptors and post-translational modifications of their associated signalling proteins. Biochem. J. 2009, 422, 1–10. [Google Scholar] [CrossRef]
- Botos, I.; Segal, D.M.; Davies, D.R. The structural biology of Toll-like receptors. Structure 2011, 19, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.C.; Avalos, A.M.; Ploegh, H.L. Accessory molecules for Toll-like receptors and their function. Nat. Rev. Immunol. 2012, 12, 168–179. [Google Scholar] [CrossRef]
- Jiang, Z.; Georgel, P.; Du, X.; Shamel, L.; Sovath, S.; Mudd, S.; Huber, M.; Kalis, C.; Keck, S.; Galanos, C.; et al. CD14 is required for MyD88-independent LPS signaling. Nat. Immunol. 2005, 6, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Baumann, C.L.; Aspalter, I.M.; Sharif, O.; Pichlmair, A.; Bluml, S.; Grebien, F.; Bruckner, M.; Pasierbek, P.; Aumayr, K.; Planyavsky, M.; et al. CD14 is a coreceptor of Toll-like receptors 7 and 9. J. Exp. Med. 2010, 207, 2689–2701. [Google Scholar] [CrossRef] [PubMed]
- Kirschning, C.J.; Schumann, R.R. TLR2: Cellular Sensor for Microbial and Endogenous Molecular Patterns. In Toll-Like Receptor Family Members and Their Ligands; Beutler, B., Wagner, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; Volume 8, pp. 121–144. [Google Scholar]
- Ewald, S.E.; Barton, G.M. Nucleic acid sensing Toll-like receptors in autoimmunity. Curr. Opin. Immunol. 2011, 23, 3–9. [Google Scholar] [CrossRef]
- Fukui, R.; Saitoh, S.; Matsumoto, F.; Kozuka-Hata, H.; Oyama, M.; Tabeta, K.; Beutler, B.; Miyake, K. Unc93B1 biases Toll-like receptor responses to nucleic acid in dendritic cells toward DNA-but against RNA-sensing. J. Exp. Med. 2009, 206, 1339–1350. [Google Scholar] [CrossRef]
- Takahashi, K.; Shibata, T.; Akashi-Takamura, S.; Kiyokawa, T.; Wakabayashi, Y.; Tanimura, N.; Kobayashi, T.; Matsumoto, F.; Fukui, R.; Kouro, T.; et al. A protein associated with Toll-like receptor (TLR) 4 (PRAT4A) is required for TLR-dependent immune responses. J. Exp. Med. 2007, 204, 2963–2976. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.; Dai, J.; Srivastava, P.K.; Zammit, D.J.; Lefrancois, L.; Li, Z. Heat shock protein gp96 is a master chaperone for toll-like receptors and is important in the innate function of macrophages. Immunity 2007, 26, 215–226. [Google Scholar] [CrossRef]
- Ewald, S.E.; Engel, A.; Lee, J.; Wang, M.; Bogyo, M.; Barton, G.M. Nucleic acid recognition by Toll-like receptors is coupled to stepwise processing by cathepsins and asparagine endopeptidase. J. Exp. Med. 2011, 208, 643–651. [Google Scholar] [CrossRef]
- Garcia-Cattaneo, A.; Gobert, F.X.; Muller, M.; Toscano, F.; Flores, M.; Lescure, A.; Del Nery, E.; Benaroch, P. Cleavage of Toll-like receptor 3 by cathepsins B and H is essential for signaling. Proc. Natl. Acad. Sci. USA 2012, 109, 9053–9058. [Google Scholar] [CrossRef]
- Park, B.; Brinkmann, M.M.; Spooner, E.; Lee, C.C.; Kim, Y.M.; Ploegh, H.L. Proteolytic cleavage in an endolysosomal compartment is required for activation of Toll-like receptor 9. Nat. Immunol. 2008, 9, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Ranoa, D.R.E.; Jiang, S.; Mutha, S.K.; Li, X.; Baudry, J.; Tapping, R.I. Human TLRs 10 and 1 Share Common Mechanisms of Innate Immune Sensing but Not Signaling. J. Immunol. 2010, 184, 5094–5103. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Mathur, R.; Oh, H.; Zhang, D.; Park, S.-G.; Seo, J.; Koblansky, A.; Hayden, M.S.; Ghosh, S. A Mouse Model of Salmonella Typhi Infection. Cell 2012, 151, 590–602. [Google Scholar] [CrossRef]
- Yarovinsky, F.; Zhang, D.; Andersen, J.F.; Bannenberg, G.L.; Serhan, C.N.; Hayden, M.S.; Hieny, S.; Sutterwala, F.S.; Flavell, R.A.; Ghosh, S.; et al. TLR11 Activation of Dendritic Cells by a Protozoan Profilin-Like Protein. Science 2005, 308, 1626–1629. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.K.; Askins, J.; Hall, P.R.; Davies, D.R.; Segal, D.M. The dsRNA binding site of human Toll-like receptor 3. Proc. Natl. Acad. Sci. USA 2006, 103, 8792–8797. [Google Scholar] [CrossRef]
- Lund, J.M.; Alexopoulou, L.; Sato, A.; Karow, M.; Adams, N.C.; Gale, N.W.; Iwasaki, A.; Flavell, R.A. Recognition of single-stranded RNA viruses by Toll-like receptor 7. Proc. Natl. Acad. Sci. 2004, 101, 5598–5603. [Google Scholar] [CrossRef]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-Specific Recognition of Single-Stranded RNA via Toll-like Receptor 7 and 8. Science 2004, 303, 1526–1529. [Google Scholar] [CrossRef]
- Bauer, S.; Kirschning, C.J.; Häcker, H.; Redecke, V.; Hausmann, S.; Akira, S.; Wagner, H.; Lipford, G.B. Human TLR9 confers responsiveness to bacterial DNA via species-specific CpG motif recognition. Proc. Natl. Acad. Sci. USA 2001, 98, 9237–9242. [Google Scholar] [CrossRef]
- Bonham, K.S.; Orzalli, M.H.; Hayashi, K.; Wolf, A.I.; Glanemann, C.; Weninger, W.; Iwasaki, A.; Knipe, D.M.; Kagan, J.C. A promiscuous lipid-binding protein diversifies the subcellular sites of toll-like receptor signal transduction. Cell 2014, 156, 705–716. [Google Scholar] [CrossRef]
- Kagan, J.C.; Medzhitov, R. Phosphoinositide-mediated adaptor recruitment controls Toll-like receptor signaling. Cell 2006, 125, 943–955. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Sato, S.; Hemmi, H.; Hoshino, K.; Kaisho, T.; Sanjo, H.; Takeuchi, O.; Sugiyama, M.; Okabe, M.; Takeda, K.; et al. Role of Adaptor TRIF in the MyD88-Independent Toll-Like Receptor Signaling Pathway. Science 2003, 301, 640–643. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.G.; Xu, S.; Kang, Z.H.; Huo, J.; Huang, M.; Liu, D.; Takeuchi, O.; Akira, S.; Lam, K.P. Bruton’s tyrosine kinase phosphorylates Toll-like receptor 3 to initiate antiviral response. Proc. Natl. Acad. Sci. USA 2012, 109, 5791–5796. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Chattopadhyay, S.; Fensterl, V.; Saikia, P.; Wetzel, J.L.; Sen, G.C. Epidermal Growth Factor Receptor Is Essential for Toll-Like Receptor 3 Signaling. Sci. Signal. 2012, 5, ra50-ra50. [Google Scholar] [CrossRef]
- Yamamoto, M.; Sato, S.; Hemmi, H.; Uematsu, S.; Hoshino, K.; Kaisho, T.; Takeuchi, O.; Takeda, K.; Akira, S. TRAM is specifically involved in the Toll-like receptor 4–mediated MyD88-independent signaling pathway. Nat. Immunol. 2003, 4, 1144–1150. [Google Scholar] [CrossRef]
- Keestra, A.M.; de Zoete, M.R.; van Aubel, R.A.M.H.; van Putten, J.P.M. The Central Leucine-Rich Repeat Region of Chicken TLR16 Dictates Unique Ligand Specificity and Species-Specific Interaction with TLR2. J. Immunol. 2007, 178, 7110–7119. [Google Scholar] [CrossRef]
- Keestra, A.M.; de Zoete, M.R.; Bouwman, L.I.; Vaezirad, M.M.; van Putten, J.P. Unique features of chicken Toll-like receptors. Dev. Comp. Immunol. 2013, 41, 316–323. [Google Scholar] [CrossRef]
- Ozinsky, A.; Underhill, D.M.; Fontenot, J.D.; Hajjar, A.M.; Smith, K.D.; Wilson, C.B.; Schroeder, L.; Aderem, A. The repertoire for pattern recognition of pathogens by the innate immune system is defined by cooperation between Toll-like receptors. Proc. Natl. Acad. Sci. USA 2000, 97, 13766–13771. [Google Scholar] [CrossRef]
- Gautam, J.K.; Ashish; Comeau, L.D.; Krueger, J.K.; Smith, M.F. Structural and Functional Evidence for the Role of the TLR2 DD Loop in TLR1/TLR2 Heterodimerization and Signaling. J. Biol. Chem. 2006, 281, 30132–30142. [Google Scholar] [CrossRef]
- Weber, A.N.R.; Moncrieffe, M.C.; Gangloff, M.; Imler, J.-L.; Gay, N.J. Ligand-Receptor and Receptor-Receptor Interactions Act in Concert to Activate Signaling in the Drosophila Toll Pathway. J. Biol. Chem. 2005, 280, 22793–22799. [Google Scholar] [CrossRef]
- Pichlmair, A.; Schulz, O.; Tan, C.P.; Naslund, T.I.; Liljestrom, P.; Weber, F.; Reis e Sousa, C. RIG-I-mediated antiviral responses to single-stranded RNA bearing 5’-phosphates. Science 2006, 314, 997–1001. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, M.; Matsuo, A.; Shingai, M.; Shida, K.; Ishii, A.; Funami, K.; Suzuki, Y.; Oshiumi, H.; Matsumoto, M.; Seya, T. Combinational recognition of bacterial lipoproteins and peptidoglycan by chicken Toll-like receptor 2 subfamily. Dev. Comp. Immunol. 2008, 32, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Ruan, W.K.; Wu, Y.H.; An, J.; Cui, D.F.; Li, H.R.; Zheng, S.J. Toll-like receptor 2 type 1 and type 2 polymorphisms in different chicken breeds. Poult. Sci. 2012, 91, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Temperley, N.D.; Ren, L.; Smith, J.; Li, N.; Burt, D.W. Molecular evolution of the vertebrate TLR1 gene family-a complex history of gene duplication, gene conversion, positive selection and co-evolution. BMC Evol. Biol. 2011, 11, 149. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Mak, T.W.; Sen, G.; Li, X. Toll-like receptor 3-mediated activation of NF-kappaB and IRF3 diverges at Toll-IL-1 receptor domain-containing adapter inducing IFN-beta. Proc. Natl. Acad. Sci. USA 2004, 101, 3533–3538. [Google Scholar] [CrossRef] [PubMed]
- Schoggins, J.W.; Wilson, S.J.; Panis, M.; Murphy, M.Y.; Jones, C.T.; Bieniasz, P.; Rice, C.M. A diverse range of gene products are effectors of the type I interferon antiviral response. Nature 2011, 472, 481–485. [Google Scholar] [CrossRef]
- Karpala, A.J.; Lowenthal, J.W.; Bean, A.G. Activation of the TLR3 pathway regulates IFNβ production in chickens. Dev. Comp. Immunol. 2008, 32, 435–444. [Google Scholar] [CrossRef]
- Schwarz, H.; Schneider, K.; Ohnemus, A.; Lavric, M.; Kothlow, S.; Bauer, S.; Kaspers, B.; Staeheli, P. Chicken toll-like receptor 3 recognizes its cognate ligand when ectopically expressed in human cells. J. Interferon Cytokine Res. 2007, 27, 97–102. [Google Scholar] [CrossRef]
- He, H.; Genovese, K.J.; Swaggerty, C.L.; MacKinnon, K.M.; Kogut, M.H. Co-stimulation with TLR3 and TLR21 ligands synergistically up-regulates Th1-cytokine IFN-γ and regulatory cytokine IL-10 expression in chicken monocytes. Dev. Comp. Immunol. 2012, 36, 756–760. [Google Scholar] [CrossRef]
- Gillespie, M.; Shamovsky, V.; D’Eustachio, P. Human and chicken TLR pathways: Manual curation and computer-based orthology analysis. Mamm. Genome 2011, 22, 130–138. [Google Scholar] [CrossRef]
- Cormican, P.; Lloyd, A.T.; Downing, T.; Connell, S.J.; Bradley, D.; O’Farrelly, C. The avian Toll-Like receptor pathway—Subtle differences amidst general conformity. Dev. Comp. Immunol. 2009, 33, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Ruan, W.; An, J.; Wu, Y. Polymorphisms of chicken TLR3 and 7 in different breeds. PLoS ONE 2015, 10, e0119967. [Google Scholar] [CrossRef] [PubMed]
- Jiao, P.R.; Wei, L.M.; Cheng, Y.Q.; Yuan, R.Y.; Han, F.; Liang, J.; Liu, W.L.; Ren, T.; Xin, C.A.; Liao, M. Molecular cloning, characterization, and expression analysis of the Muscovy duck Toll-like receptor 3 (MdTLR3) gene. Poult. Sci. 2012, 91, 2475–2481. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Song, K.; Li, C.; Chen, Z.; Ding, C.; Liu, G. Molecular cloning of Peking duck Toll-like receptor 3 (duTLR3) gene and its responses to reovirus infection. Virol. J. 2015, 12, 207. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yong, Y.-H.; Liu, S.-F.; Hua, G.-H.; Jia, R.-M.; Gooneratne, R.; Zhao, Y.-T.; Liao, M.; Ju, X.-H. Goose toll-like receptor 3 (TLR3) mediated IFN-γ and IL-6 in anti-H5N1 avian influenza virus response. Vet. Immunol. Immunopathol. 2018, 197, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Zanoni, I.; Ostuni, R.; Marek, L.R.; Barresi, S.; Barbalat, R.; Barton, G.M.; Granucci, F.; Kagan, J.C. CD14 controls the LPS-induced endocytosis of Toll-like receptor 4. Cell 2011, 147, 868–880. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, K.; Takeuchi, O.; Kawai, T.; Sanjo, H.; Ogawa, T.; Takeda, Y.; Takeda, K.; Akira, S. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: evidence for TLR4 as the Lps gene product. J. Immunol. 1999, 162, 3749–3752. [Google Scholar]
- Hajjar, A.M.; Ernst, R.K.; Tsai, J.H.; Wilson, C.B.; Miller, S.I. Human Toll-like receptor 4 recognizes host-specific LPS modifications. Nat. Immunol. 2002, 3, 354–359. [Google Scholar] [CrossRef]
- Muroi, M.; Tanamoto, K. Structural regions of MD-2 that determine the agonist-antagonist activity of lipid IVa. J. Biol. Chem. 2006, 281, 5484–5491. [Google Scholar] [CrossRef]
- Karnati, H.K.; Pasupuleti, S.R.; Kandi, R.; Undi, R.B.; Sahu, I.; Kannaki, T.R.; Subbiah, M.; Gutti, R.K. TLR-4 signalling pathway: MyD88 independent pathway up-regulation in chicken breeds upon LPS treatment. Vet. Res. Commun. 2015, 39, 73–78. [Google Scholar] [CrossRef]
- Wu, Z.; Rothwell, L.; Hu, T.; Kaiser, P. Chicken CD14, unlike mammalian CD14, is trans-membrane rather than GPI-anchored. Dev. Comp. Immunol. 2009, 33, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Li, G.; Li, J.; Tian, Y.; Wang, D.; Shen, J.; Tao, Z.; Xu, J.; Lu, L. Cloning, expression and bioinformatics analysis of the duck TLR 4 gene. Br. Poult. Sci. 2012, 53, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Huang, Z.; Chen, Y.; Zhang, Y.; Rong, G.; Mu, C.; Xu, Q.; Chen, G. Molecular cloning and functional analysis of the duck TLR4 gene. Int. J. Mol. Sci. 2013, 14, 18615–18628. [Google Scholar] [CrossRef] [PubMed]
- Keestra, A.M.; de Zoete, M.R.; van Aubel, R.A.; van Putten, J.P. Functional characterization of chicken TLR5 reveals species-specific recognition of flagellin. Mol. Immunol. 2008, 45, 1298–1307. [Google Scholar] [CrossRef] [PubMed]
- de Zoete, M.R.; Keestra, A.M.; Roszczenko, P.; van Putten, J.P.M. Activation of Human and Chicken Toll-Like Receptors by Campylobacter spp. Immunol. Infect. 2010, 78, 1229–1238. [Google Scholar] [CrossRef]
- de Zoete, M.R.; Keestra, A.M.; Wagenaar, J.A.; van Putten, J.P.M. Reconstitution of a Functional Toll-like Receptor 5 Binding Site in Campylobacter jejuni Flagellin. J. Biol. Chem. 2010, 285, 12149–12158. [Google Scholar] [CrossRef]
- Fang, Q.; Pan, Z.; Geng, S.; Kang, X.; Huang, J.; Sun, X.; Li, Q.; Cai, Y.; Jiao, X. Molecular cloning, characterization and expression of goose Toll-like receptor 5. Mol. Immunol. 2012, 52, 117–124. [Google Scholar] [CrossRef]
- Cheng, Y.; Sun, Y.; Wang, H.; Shi, S.; Yan, Y.; Li, J.; Ding, C.; Sun, J. Cloning, expression and functional analysis of the duck Toll-like receptor 5 (TLR5) gene. J. Vet. Sci. 2015, 16, 37–46. [Google Scholar] [CrossRef]
- Xiong, D.; Pan, Z.; Kang, X.; Wang, J.; Song, L.; Jiao, X. Molecular cloning and functional analysis of duck Toll-like receptor 5. Res. Vet. Sci. 2014, 97, 43–45. [Google Scholar] [CrossRef]
- Philbin, V.J.; Iqbal, M.; Boyd, Y.; Goodchild, M.J.; Beal, R.K.; Bumstead, N.; Young, J.; Smith, A.L. Identification and characterization of a functional, alternatively spliced Toll-like receptor 7 (TLR7) and genomic disruption of TLR8 in chickens. Immunology 2005, 114, 507–521. [Google Scholar] [CrossRef]
- Stewart, C.R.; Bagnaud-Baule, A.; Karpala, A.J.; Lowther, S.; Mohr, P.G.; Wise, T.G.; Lowenthal, J.W.; Bean, A.G. Toll-like receptor 7 ligands inhibit influenza A infection in chickens. J. Interferon. Cytokine Res. 2012, 32, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Chen, S.; Zhao, Q.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Liu, F.; Chen, X.; Cheng, A. Molecular cloning, tissue distribution, and immune function of goose TLR7. Immunol. Lett. 2015, 163, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Jiao, P.; Yuan, R.; Song, Y.; Cui, P.; Guo, X.; Zheng, B.; Jia, W.; Qi, W.; Ren, T.; et al. Goose Toll-like receptor 7 (TLR7), myeloid differentiation factor 88 (MyD88) and antiviral molecules involved in anti-H5N1 highly pathogenic avian influenza virus response. Vet. Immunol. Immunopathol. 2013, 153, 99–106. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, M.R.W.; Xia, J.; Smith, A.L.; Magor, K.E. The duck toll like receptor 7: Genomic organization, expression and function. Mol. Immunol. 2008, 45, 2055–2061. [Google Scholar] [CrossRef] [PubMed]
- Davis, B.K.; Roberts, R.A.; Huang, M.T.; Willingham, S.B.; Conti, B.J.; Brickey, W.J.; Barker, B.R.; Kwan, M.; Taxman, D.J.; Accavitti-Loper, M.A.; et al. Cutting edge: NLRC5-dependent activation of the inflammasome. J. Immunol. 2011, 186, 1333–1337. [Google Scholar] [CrossRef] [PubMed]
- Boyd, A.C.; Peroval, M.Y.; Hammond, J.A.; Prickett, M.D.; Young, J.R.; Smith, A.L. TLR15 is unique to avian and reptilian lineages and recognizes a yeast-derived agonist. J. Immunol. 2012, 189, 4930–4938. [Google Scholar] [CrossRef]
- Ramasamy, K.T.; Reddy, M.R.; Verma, P.C.; Murugesan, S. Expression analysis of turkey (Meleagris gallopavo) toll-like receptors and molecular characterization of avian specific TLR15. Mol. Biol. Rep. 2012, 39, 8539–8549. [Google Scholar] [CrossRef]
- Voogdt, C.G.P.; Merchant, M.E.; Wagenaar, J.A.; van Putten, J.P.M. Evolutionary Regression and Species-Specific Codon Usage of TLR15. Front. Immunol. 2018, 9, 2626. [Google Scholar] [CrossRef]
- de Zoete, M.R.; Bouwman, L.I.; Keestra, A.M.; van Putten, J.P.M. Cleavage and activation of a Toll-like receptor by microbial proteases. Proc. Natl. Acad. Sci. USA 2011, 108, 4968–4973. [Google Scholar] [CrossRef]
- Higgs, R.; Cormican, P.; Cahalane, S.; Allan, B.; Lloyd, A.T.; Meade, K.; James, T.; Lynn, D.J.; Babiuk, L.A.; O’Farrelly, C. Induction of a Novel Chicken Toll-Like Receptor following Salmonella enterica Serovar Typhimurium Infection. Immunol. Infect. 2006, 74, 1692–1698. [Google Scholar] [CrossRef]
- Shaughnessy, R.G.; Meade, K.G.; Cahalane, S.; Allan, B.; Reiman, C.; Callanan, J.J.; O’Farrelly, C. Innate immune gene expression differentiates the early avian intestinal response between Salmonella and Campylobacter. Vet. Immunol. Immunopathol. 2009, 132, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Oven, I.; Resman Rus, K.; Dusanic, D.; Bencina, D.; Keeler, C.L., Jr.; Narat, M. Diacylated lipopeptide from Mycoplasma synoviae mediates TLR15 induced innate immune responses. Vet. Res. 2013, 44, 99. [Google Scholar] [CrossRef] [PubMed]
- Palti, Y. Toll-like receptors in bony fish: From genomics to function. Dev. Comp. Immunol. 2011, 35, 1263–1272. [Google Scholar] [CrossRef] [PubMed]
- Pelka, K.; Bertheloot, D.; Reimer, E.; Phulphagar, K.; Schmidt, S.V.; Christ, A.; Stahl, R.; Watson, N.; Miyake, K.; Hacohen, N.; et al. The chaperone UNC93B1 regulates Toll-like receptor stability independently of endosomal TLR transport. Immunity 2018, 48, 911–922 e917. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Wu, X.; Jia, R.; Wang, M.; Chen, S.; Liu, M.; Zhu, D.; Zhao, X.; Wu, Y.; Yang, Q.; et al. CpG oligodeoxynucleotide-specific duck TLR21 mediates activation of NF-κB signaling pathway and plays an important role in the host defence of DPV infection. Mol. Immunol. 2019, 106, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Yan, B.; Chen, S.; Chen, H.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Liu, F.; Yang, Q.; et al. CpG oligodeoxynucleotide-specific goose TLR21 initiates an anti-viral immune response against NGVEV but not AIV strain H9N2 infection. Immunobiology 2016, 221, 454–461. [Google Scholar] [CrossRef]
- Broz, P.; Dixit, V.M. Inflammasomes: Mechanism of assembly, regulation and signalling. Nat. Rev. Immunol. 2016, 16, 407–420. [Google Scholar] [CrossRef]
- Cridland, J.A.; Curley, E.Z.; Wykes, M.N.; Schroder, K.; Sweet, M.J.; Roberts, T.L.; Ragan, M.A.; Kassahn, K.S.; Stacey, K.J. The mammalian PYHIN gene family: Phylogeny, evolution and expression. BMC Evol. Biol. 2012, 12, 140. [Google Scholar] [CrossRef]
- Evavold, C.L.; Kagan, J.C. Inflammasomes: threat-assessment organelles of the innate immune system. Immunity 2019, 51, 609–624. [Google Scholar] [CrossRef]
- Orning, P.; Lien, E.; Fitzgerald, K.A. Gasdermins and their role in immunity and inflammation. J. Exp. Med. 2019, 216, 2453–2465. [Google Scholar] [CrossRef]
- Moore, C.B.; Bergstralh, D.T.; Duncan, J.A.; Lei, Y.; Morrison, T.E.; Zimmermann, A.G.; Accavitti-Loper, M.A.; Madden, V.J.; Sun, L.; Ye, Z.; et al. NLRX1 is a regulator of mitochondrial antiviral immunity. Nature 2008, 451, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Mo, J.; Swanson, K.V.; Wen, H.; Petrucelli, A.; Gregory, S.M.; Zhang, Z.; Schneider, M.; Jiang, Y.; Fitzgerald, K.A.; et al. NLRC3, a member of the NLR family of proteins, is a negative regulator of innate immune signaling induced by the DNA sensor STING. Immunity 2014, 40, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Yoshihama, S.; Roszik, J.; Downs, I.; Meissner, T.B.; Vijayan, S.; Chapuy, B.; Sidiq, T.; Shipp, M.A.; Lizee, G.A.; Kobayashi, K.S. NLRC5/MHC class I transactivator is a target for immune evasion in cancer. Proc. Natl. Acad. Sci. USA 2016, 113, 5999–6004. [Google Scholar] [CrossRef] [PubMed]
- Arnoult, D.; Soares, F.; Tattoli, I.; Castanier, C.; Philpott, D.J.; Girardin, S.E. An N-terminal addressing sequence targets NLRX1 to the mitochondrial matrix. J. Cell Sci. 2009, 122, 3161–3168. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Wen, H.; Yu, Y.; Taxman, D.J.; Zhang, L.; Widman, D.G.; Swanson, K.V.; Wen, K.W.; Damania, B.; Moore, C.B.; et al. The mitochondrial proteins NLRX1 and TUFM form a complex that regulates type I interferon and autophagy. Immunity 2012, 36, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yao, Y.; Qiu, X.; Wang, G.; Hu, Z.; Chen, S.; Wu, Z.; Yuan, N.; Gao, H.; Wang, J.; et al. Listeria hijacks host mitophagy through a novel mitophagy receptor to evade killing. Nat. Immunol. 2019, 20, 433–446. [Google Scholar] [CrossRef]
- Hong, M.; Yoon, S.-I.; Wilson, I.A. Structure and functional characterization of the RNA-binding element of the NLRX1 innate immune modulator. Immunity 2012, 36, 337–347. [Google Scholar] [CrossRef]
- Swanson, K.V.; Deng, M.; Ting, J.P.Y. The NLRP3 inflammasome: Molecular activation and regulation to therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef]
- He, Y.; Zeng, M.Y.; Yang, D.; Motro, B.; Núñez, G. NEK7 is an essential mediator of NLRP3 activation downstream of potassium efflux. Nature 2016, 530, 354–357. [Google Scholar] [CrossRef]
- Li, R.; Lin, J.; Hou, X.; Han, S.; Weng, H.; Xu, T.; Li, N.; Chai, T.; Wei, L. Characterization and roles of cherry valley duck NLRP3 in innate immunity during avian pathogenic escherichia coli infection. Front. Immunol. 2018, 9, 2300. [Google Scholar] [CrossRef]
- Schneider, M.; Zimmermann, A.G.; Roberts, R.A.; Zhang, L.; Swanson, K.V.; Wen, H.; Davis, B.K.; Allen, I.C.; Holl, E.K.; Ye, Z.; et al. The innate immune sensor NLRC3 attenuates Toll-like receptor signaling via modification of the signaling adaptor TRAF6 and transcription factor NF-κB. Nat. Immunol. 2012, 13, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Meissner, T.B.; Li, A.; Biswas, A.; Lee, K.-H.; Liu, Y.-J.; Bayir, E.; Iliopoulos, D.; van den Elsen, P.J.; Kobayashi, K.S. NLR family member NLRC5 is a transcriptional regulator of MHC class I genes. Proc. Natl. Acad. Sci. USA 2010, 107, 13794–13799. [Google Scholar] [CrossRef] [PubMed]
- Meissner, T.B.; Li, A.; Liu, Y.-J.; Gagnon, E.; Kobayashi, K.S. The nucleotide-binding domain of NLRC5 is critical for nuclear import and transactivation activity. Biochem. Biophys. Res. Commun. 2012, 418, 786–791. [Google Scholar] [CrossRef] [PubMed]
- Staehli, F.; Ludigs, K.; Heinz, L.X.; Seguin-Estevez, Q.; Ferrero, I.; Braun, M.; Schroder, K.; Rebsamen, M.; Tardivel, A.; Mattmann, C.; et al. NLRC5 deficiency selectively impairs MHC class I- dependent lymphocyte killing by cytotoxic T cells. J. Immunol. 2012, 188, 3820–3828. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Cui, J.; Li, Q.; Zou, J.; Wang, H.Y.; Wang, R.F. Enhanced TLR-induced NF-kappaB signaling and type I interferon responses in NLRC5 deficient mice. Cell Res. 2012, 22, 822–835. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Zhu, L.; Xia, X.; Wang, H.Y.; Legras, X.; Hong, J.; Ji, J.; Shen, P.; Zheng, S.; Chen, Z.J.; et al. NLRC5 negatively regulates the NF-kappaB and type I interferon signaling pathways. Cell 2010, 141, 483–496. [Google Scholar] [CrossRef]
- Cadena, C.; Hur, S. Filament-like Assemblies of Intracellular Nucleic Acid Sensors: Commonalities and Differences. Mol. Cell 2019, 76, 243–254. [Google Scholar] [CrossRef]
- Myong, S.; Cui, S.; Cornish, P.V.; Kirchhofer, A.; Gack, M.U.; Jung, J.U.; Hopfner, K.P.; Ha, T. Cytosolic viral sensor RIG-I is a 5’-triphosphate-dependent translocase on double-stranded RNA. Science 2009, 323, 1070–1074. [Google Scholar] [CrossRef]
- Cadena, C.; Ahmad, S.; Xavier, A.; Willemsen, J.; Park, S.; Park, J.W.; Oh, S.W.; Fujita, T.; Hou, F.; Binder, M.; et al. Ubiquitin-dependent and-independent roles of E3 ligase RIPLET in innate immunity. Cell 2019, 177, 1187–1200. [Google Scholar] [CrossRef]
- Sun, Y.; Ding, N.; Ding, S.S.; Yu, S.; Meng, C.; Chen, H.; Qiu, X.; Zhang, S.; Yu, Y.; Zhan, Y.; et al. Goose RIG-I functions in innate immunity against Newcastle disease virus infections. Mol. Immunol. 2013, 53, 321–327. [Google Scholar] [CrossRef]
- Barber, M.R.; Aldridge, J.R.; Webster, R.G.; Magor, K.E. Association of RIG-I with innate immunity of ducks to influenza. Proc. Natl. Acad. Sci. USA 2010, 107, 5913–5918. [Google Scholar] [CrossRef] [PubMed]
- Miranzo-Navarro, D.; Magor, K.E. Activation of duck RIG-I by TRIM25 is independent of anchored ubiquitin. PLoS ONE 2014, 9, e86968. [Google Scholar] [CrossRef]
- Hayman, T.J.; Hsu, A.C.; Kolesnik, T.B.; Dagley, L.F.; Willemsen, J.; Tate, M.D.; Baker, P.J.; Kershaw, N.J.; Kedzierski, L.; Webb, A.I.; et al. RIPLET, and not TRIM25, is required for endogenous RIG-I-dependent antiviral responses. Immunol. Cell Biol. 2019, 97, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Peisley, A.; Wu, B.; Xu, H.; Chen, Z.J.; Hur, S. Structural basis for ubiquitin-mediated antiviral signal activation by RIG-I. Nature 2014, 509, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Barber, M.R.W.; Aldridge, J.R.; Fleming-Canepa, X.; Wang, Y.-D.; Webster, R.G.; Magor, K.E. Identification of avian RIG-I responsive genes during influenza infection. Mol. Immunol. 2013, 54, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Fournier, P.; Wilden, H.; Schirrmacher, V. Importance of retinoic acid-inducible gene I and of receptor for type I interferon for cellular resistance to infection by Newcastle disease virus. Int. J. Oncol. 2012, 40, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Biswas, M.; Kumar, S.R.P.; Allen, A.; Yong, W.; Nimmanapalli, R.; Samal, S.K.; Elankumaran, S. Cell-type-specific innate immune response to oncolytic Newcastle disease virus. Viral Immunol. 2012, 25, 268–276. [Google Scholar] [CrossRef]
- Kawai, T.; Takahashi, K.; Sato, S.; Coban, C.; Kumar, H.; Kato, H.; Ishii, K.J.; Takeuchi, O.; Akira, S. IPS-1, an adaptor triggering RIG-I- and Mda5-mediated type I interferon induction. Nat. Immunol. 2005, 6, 981–988. [Google Scholar] [CrossRef]
- Vazquez, C.; Horner, S.M. MAVS Coordination of Antiviral Innate Immunity. J. Virol. 2015, 89, 6974–6977. [Google Scholar] [CrossRef]
- Zhao, B.; Shu, C.; Gao, X.; Sankaran, B.; Du, F.; Shelton, C.L.; Herr, A.B.; Ji, J.-Y.; Li, P. Structural basis for concerted recruitment and activation of IRF-3 by innate immune adaptor proteins. Proc. Natl. Acad. Sci. USA 2016, 113, E3403–E3412. [Google Scholar] [CrossRef]
- Sun, Y.; Mao, X.; Zheng, H.; Wu, W.; Rehman, Z.U.; Liao, Y.; Meng, C.; Qiu, X.; Tan, L.; Song, C.; et al. Goose MAVS functions in RIG-I-mediated IFN-β signaling activation. Dev. Comp. Immunol. 2019, 93, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhai, Y.; Fan, Y.; Chen, H.; Zhang, A.; Jin, H.; Luo, R. Molecular cloning and functional characterization of duck mitochondrial antiviral-signaling protein (MAVS). Dev. Comp. Immunol. 2016, 56, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Peisley, A.; Jo, M.H.; Lin, C.; Wu, B.; Orme-Johnson, M.; Walz, T.; Hohng, S.; Hur, S. Kinetic mechanism for viral dsRNA length discrimination by MDA5 filaments. Proc. Natl. Acad. Sci. USA 2012, 109, E3340–E3349. [Google Scholar] [CrossRef] [PubMed]
- Lang, X.; Tang, T.; Jin, T.; Ding, C.; Zhou, R.; Jiang, W. TRIM65-catalized ubiquitination is essential for MDA5-mediated antiviral innate immunity. J. Exp. Med. 2017, 214, 459–473. [Google Scholar] [CrossRef]
- Karpala, A.J.; Stewart, C.; McKay, J.; Lowenthal, J.W.; Bean, A.G.D. Characterization of Chicken Mda5 Activity: Regulation of IFN-β in the Absence of RIG-I Functionality. J. Immunol. Res. 2011, 186, 5397–5405. [Google Scholar] [CrossRef]
- Hayashi, T.; Watanabe, C.; Suzuki, Y.; Tanikawa, T.; Uchida, Y.; Saito, T. Chicken MDA5 senses short double-stranded RNA with implications for antiviral response against avian influenza viruses in chicken. J. Innate Immun. 2014, 6, 58–71. [Google Scholar] [CrossRef]
- Wei, L.; Cui, J.; Song, Y.; Zhang, S.; Han, F.; Yuan, R.; Gong, L.; Jiao, P.; Liao, M. Duck MDA5 functions in innate immunity against H5N1 highly pathogenic avian influenza virus infections. Vet. Res. 2014, 45, 66. [Google Scholar] [CrossRef]
- Wei, L.M.; Jiao, P.R.; Song, Y.F.; Han, F.; Cao, L.; Yang, F.; Ren, T.; Liao, M. Identification and expression profiling analysis of goose melanoma differentiation associated gene 5 (MDA5) gene. Poult. Sci. 2013, 92, 2618–2624. [Google Scholar] [CrossRef]
- Satoh, T.; Kato, H.; Kumagai, Y.; Yoneyama, M.; Sato, S.; Matsushita, K.; Tsujimura, T.; Fujita, T.; Akira, S.; Takeuchi, O. LGP2 is a positive regulator of RIG-I– and MDA5-mediated antiviral responses. Proc. Natl. Acad. Sci. USA 2010, 107, 1512–1517. [Google Scholar] [CrossRef]
- Liniger, M.; Summerfield, A.; Zimmer, G.; McCullough, K.C.; Ruggli, N. Chicken cells sense influenza a virus infection through MDA5 and CARDIF signaling involving LGP2. J. Virol. 2012, 86, 705–717. [Google Scholar] [CrossRef]
- Huo, H.; Zhao, L.; Wang, D.; Chen, X.; Chen, H. LGP2 plays a critical role in MDA5-mediated antiviral activity against duck enteritis virus. Mol. Immunol. 2019, 116, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Song, Y.; Cui, J.; Qu, N.; Wang, N.; Ouyang, G.; Liao, M.; Jiao, P. Cloning, characterization, and expression analysis of LGP2 cDNA from goose, Anser cygnoides. Poult. Sci. 2016, 95, 2290–2296. [Google Scholar] [CrossRef] [PubMed]
- Jiao, P.R.; Wei, L.M.; Song, Y.F.; Cui, J.; Zhang, S.; Han, F.; Yuan, R.Y.; Liao, M. Molecular cloning and immune responsive expression of LGP2 gene, a pivotal member of the RLR gene family from Muscovy duck Cairina moschata. Poult. Sci. 2015, 94, 1170–1176. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Ghosh, A.; Sarkar, S.N. OASL-a new player in controlling antiviral innate immunity. Curr. Opin. Virol. 2015, 12, 15–19. [Google Scholar] [CrossRef]
- Choi, U.Y.; Kang, J.-S.; Hwang, Y.S.; Kim, Y.-J. Oligoadenylate synthase-like (OASL) proteins: Dual functions and associations with diseases. Exp. Mol. Med. 2015, 47, e144–e144. [Google Scholar] [CrossRef]
- Rong, E.; Wang, X.; Chen, H.; Yang, C.; Hu, J.; Liu, W.; Wang, Z.; Chen, X.; Zheng, H.; Pu, J.; et al. Molecular mechanisms for the adaptive switching between the OAS/RNase L and OASL/RIG-I pathways in birds and mammals. Front. Immunol. 2018, 9, 1398. [Google Scholar] [CrossRef]
- Ghosh, A.; Shao, L.; Sampath, P.; Zhao, B.; Patel, N.V.; Zhu, J.; Behl, B.; Parise, R.A.; Beumer, J.H.; O’Sullivan, R.J.; et al. Oligoadenylate-synthetase-family protein OASL inhibits activity of the DNA sensor cGAS during DNA virus infection to limit interferon production. Immunity 2019, 50, 51–63 e55. [Google Scholar] [CrossRef]
- Chen, S.; Yang, C.; Zhang, J.; Wu, Z.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Yang, Q.; Wu, Y.; et al. Conserved active-site residues associated with OAS enzyme activity and ubiquitin-like domains are not required for the antiviral activity of goOASL protein against avian tembusu virus. Viruses 2018, 10. [Google Scholar] [CrossRef]
- Tag-El-Din-Hassan, H.T.; Sasaki, N.; Moritoh, K.; Torigoe, D.; Maeda, A.; Agui, T. The chicken 2’-5’ oligoadenylate synthetase A inhibits the replication of West Nile virus. Jpn. J. Vet. Res. 2012, 60, 95–103. [Google Scholar]
- Tatsumi, R.; Sekiya, S.; Nakanishi, R.; Mizutani, M.; Kojima, S.; Sokawa, Y. Function of ubiquitin-like domain of chicken 2’-5’-oligoadenylate synthetase in conformational stability. J. Interferon Cytokine Res. 2003, 23, 667–676. [Google Scholar] [CrossRef]
- Tag-El-Din-Hassan, H.T.; Sasaki, N.; Torigoe, D.; Morimatsu, M.; Agui, T. Analysis of the relationship between enzymatic and antiviral activities of the chicken oligoadenylate synthetase-like. J. Interferon Cytokine Res. 2017, 37, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Liu, F.; Chen, S.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Sun, K.; Yang, Q.; Wu, Y.; et al. Identification of 2’-5’-Oligoadenylate Synthetase-Like gene in goose: Gene structure, expression patterns, and antiviral activity against Newcastle disease virus. J. Interferon Cytokine Res. 2016, 36, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Gal-Ben-Ari, S.; Barrera, I.; Ehrlich, M.; Rosenblum, K. PKR: A kinase to remember. Front. Mol. Neurosci. 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- McKenna, S.A.; Kim, I.; Liu, C.W.; Puglisi, J.D. Uncoupling of RNA binding and PKR kinase activation by viral inhibitor RNAs. J. Mol. Biol. 2006, 358, 1270–1285. [Google Scholar] [CrossRef]
- Dar, A.C.; Dever, T.E.; Sicheri, F. Higher-order substrate recognition of eIF2α by the RNA-dependent protein kinase PKR. Cell 2005, 122, 887–900. [Google Scholar] [CrossRef]
- Ko, J.H.; Asano, A.; Kon, Y.; Watanabe, T.; Agui, T. Characterization of the chicken PKR: Polymorphism of the gene and antiviral activity against vesicular stomatitis virus. Jpn. J. Vet. Res. 2004, 51, 123–133. [Google Scholar]
- Liu, W.J.; Yang, Y.T.; Huang, Y.M.; Zhou, D.R.; Xu, D.N.; Cao, N.; Jiang, D.L.; Pan, J.Q.; Tian, Y.B. Identification of Goose PKR Gene: Structure, Expression Profiling, and Antiviral Activity Against Newcastle Disease Virus. J. Interferon Cytokine Res. 2018, 38, 333–340. [Google Scholar] [CrossRef]
- Zhang, Z.; Kim, T.; Bao, M.; Facchinetti, V.; Jung, S.Y.; Ghaffari, A.A.; Qin, J.; Cheng, G.; Liu, Y.-J. DDX1, DDX21, and DHX36 helicases form a complex with the adaptor molecule TRIF to sense dsRNA in dendritic cells. Immunity 2011, 34, 866–878. [Google Scholar] [CrossRef]
- Yoo, J.-S.; Takahasi, K.; Ng, C.S.; Ouda, R.; Onomoto, K.; Yoneyama, M.; Lai, J.C.; Lattmann, S.; Nagamine, Y.; Matsui, T.; et al. DHX36 enhances RIG-I signaling by facilitating PKR-mediated antiviral stress granule formation. PLoS Pathog. 2014, 10, e1004012. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Chan, C.-H.; Chen, C.-M.; Tsai, Y.-S.; Tsai, T.-Y.; Wu Lee, Y.-H.; You, L.-R. Targeted inactivation of murine Ddx3x: Essential roles of Ddx3x in placentation and embryogenesis. Human Mol. Genet. 2016, 25, 2905–2922. [Google Scholar] [CrossRef][Green Version]
- Szappanos, D.; Tschismarov, R.; Perlot, T.; Westermayer, S.; Fischer, K.; Platanitis, E.; Kallinger, F.; Novatchkova, M.; Lassnig, C.; Müller, M.; et al. The RNA helicase DDX3X is an essential mediator of innate antimicrobial immunity. PLoS Pathog. 2018, 14, e1007397. [Google Scholar] [CrossRef]
- Oshiumi, H.; Sakai, K.; Matsumoto, M.; Seya, T. DEAD/H BOX 3 (DDX3) helicase binds the RIG-I adaptor IPS-1 to up-regulate IFN-β-inducing potential. Eur. J. Immunol. 2010, 40, 940–948. [Google Scholar] [CrossRef] [PubMed]
- Soulat, D.; Bürckstümmer, T.; Westermayer, S.; Goncalves, A.; Bauch, A.; Stefanovic, A.; Hantschel, O.; Bennett, K.L.; Decker, T.; Superti-Furga, G. The DEAD-box helicase DDX3X is a critical component of the TANK-binding kinase 1-dependent innate immune response. EMBO J. 2008, 27, 2135–2146. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.; Cheng, Y.; Wang, H.; Yan, Y.; Sun, J. Chicken DDX3X Activates IFN-β via the chSTING-chIRF7-IFN-β Signaling Axis. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Cao, Y.; Ling, T.; Li, P.; Wu, S.; Peng, D.; Wang, Y.; Jia, X.; Chen, S.; Xu, A.; et al. DDX23, an Evolutionary Conserved dsRNA Sensor, Participates in Innate Antiviral Responses by Pairing With TRIF or MAVS. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Moore, R.; Xu, X.; Barber, G.N. DDX24 negatively regulates cytosolic RNA-mediated innate immune signaling. PLoS Pathog. 2013, 9, e1003721. [Google Scholar] [CrossRef]
- Oshiumi, H.; Miyashita, M.; Okamoto, M.; Morioka, Y.; Okabe, M.; Matsumoto, M.; Seya, T. DDX60 is involved in RIG-I-dependent and independent antiviral responses, and its function is attenuated by virus-induced EGFR activation. Cell Rep. 2015, 11, 1193–1207. [Google Scholar] [CrossRef]
- Wang, Y.; Yuan, S.; Jia, X.; Ge, Y.; Ling, T.; Nie, M.; Lan, X.; Chen, S.; Xu, A. Mitochondria-localised ZNFX1 functions as a dsRNA sensor to initiate antiviral responses through MAVS. Nat. Cell Biol. 2019, 21, 1346–1356. [Google Scholar] [CrossRef]
- Tamayo, R.; Pratt, J.T.; Camilli, A. Roles of cyclic diguanylate in the regulation of bacterial pathogenesis. Annu. Rev. Microbiol. 2007, 61, 131–148. [Google Scholar] [CrossRef]
- Xia, P.; Wang, S.; Xiong, Z.; Zhu, X.; Ye, B.; Du, Y.; Meng, S.; Qu, Y.; Liu, J.; Gao, G.; et al. The ER membrane adaptor ERAdP senses the bacterial second messenger c-di-AMP and initiates anti-bacterial immunity. Nat. Immunol. 2018, 19, 141–150. [Google Scholar] [CrossRef]
- McFarland, A.P.; Luo, S.; Ahmed-Qadri, F.; Zuck, M.; Thayer, E.F.; Goo, Y.A.; Hybiske, K.; Tong, L.; Woodward, J.J. Sensing of bacterial cyclic dinucleotides by the oxidoreductase RECON promotes NF-κB activation and shapes a proinflammatory antibacterial state. Immunity 2017, 46, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Oppenheimer-Shaanan, Y.; Wexselblatt, E.; Katzhendler, J.; Yavin, E.; Ben-Yehuda, S. c-di-AMP reports DNA integrity during sporulation in Bacillus subtilis. EMBO Rep. 2011, 12, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Corrigan, R.M.; Abbott, J.C.; Burhenne, H.; Kaever, V.; Grundling, A. c-di-AMP is a new second messenger in Staphylococcus aureus with a role in controlling cell size and envelope stress. PLoS Pathog. 2011, 7, e1002217. [Google Scholar] [CrossRef] [PubMed]
- Woodward, J.J.; Iavarone, A.T.; Portnoy, D.A. c-di-AMP secreted by intracellular Listeria monocytogenes activates a host type I interferon response. Science 2010, 328, 1703–1705. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Omura, H.; Ishitani, R.; Nureki, O. Cyclic GMP–AMP as an endogenous second messenger in innate immune signaling by cytosolic DNA. Annu. Rev. Biochem. 2017, 86, 541–566. [Google Scholar] [CrossRef] [PubMed]
- Andreeva, L.; Hiller, B.; Kostrewa, D.; Lässig, C.; de Oliveira Mann, C.C.; Jan Drexler, D.; Maiser, A.; Gaidt, M.; Leonhardt, H.; Hornung, V.; et al. cGAS senses long and HMGB/TFAM-bound U-turn DNA by forming protein–DNA ladders. Nature 2017, 549, 394–398. [Google Scholar] [CrossRef]
- Hornung, V.; Hartmann, R.; Ablasser, A.; Hopfner, K.-P. OAS proteins and cGAS: Unifying concepts in sensing and responding to cytosolic nucleic acids. Nat. Rev. Immunol. 2014, 14, 521–528. [Google Scholar] [CrossRef]
- Srikanth, S.; Woo, J.S.; Wu, B.; El-Sherbiny, Y.M.; Leung, J.; Chupradit, K.; Rice, L.; Seo, G.J.; Calmettes, G.; Ramakrishna, C.; et al. The Ca2+ sensor STIM1 regulates the type I interferon response by retaining the signaling adaptor STING at the endoplasmic reticulum. Nat. Immunol. 2019, 20, 152–162. [Google Scholar] [CrossRef]
- de Oliveira Mann, C.C.; Orzalli, M.H.; King, D.S.; Kagan, J.C.; Lee, A.S.Y.; Kranzusch, P.J. Modular architecture of the STING C-terminal tail allows interferon and NF-κB signaling adaptation. Cell Rep. 2019, 27, 1165–1175. [Google Scholar] [CrossRef]
- Gentili, M.; Lahaye, X.; Nadalin, F.; Nader, G.P.F.; Puig Lombardi, E.; Herve, S.; De Silva, N.S.; Rookhuizen, D.C.; Zueva, E.; Goudot, C.; et al. The N-terminal domain of cGAS determines preferential association with centromeric DNA and innate immune activation in the nucleus. Cell Rep. 2019, 26, 2377–2393.e2313. [Google Scholar] [CrossRef]
- Wu, X.; Wu, F.-H.; Wang, X.; Wang, L.; Siedow, J.N.; Zhang, W.; Pei, Z.-M. Molecular evolutionary and structural analysis of the cytosolic DNA sensor cGAS and STING. Nucleic Acid Res. 2014, 42, 8243–8257. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Liu, Y.; Xu, Z.; Zhang, Y.; Luo, D.; Gao, Y.; Qian, Y.; Bao, C.; Liu, C.; Zhang, Y.; et al. Avian oncogenic herpesvirus antagonizes the cGAS-STING DNA-sensing pathway to mediate immune evasion. PLoS Pathog. 2019, 15, e1007999. [Google Scholar] [CrossRef]
- Gao, L.; Li, K.; Zhang, Y.; Liu, Y.; Liu, C.; Zhang, Y.; Gao, Y.; Qi, X.; Cui, H.; Wang, Y.; et al. Inhibition of DNA-sensing pathway by Marek’s disease virus VP23 protein through suppression of interferon regulatory factor 7 activation. J. Virol. 2019, 93, e01934-01918. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhu, W.; Ding, C.; Niu, Q.; Wang, H.; Yan, Y.; Sun, J. IRF7 Is Involved in Both STING and MAVS Mediating IFN-β Signaling in IRF3-Lacking Chickens. J. Immunol. 2019, 203, 1930–1942. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wu, Z.; Zhang, J.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Sun, K.; Yang, Q.; Wu, Y.; et al. Duck stimulator of interferon genes plays an important role in host anti-duck plague virus infection through an IFN-dependent signalling pathway. Cytokine 2018, 102, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Liu, Y.; Shi, S.; Niu, Q.; Zhu, W.; Wang, Z.; Ma, J.; Wang, H.; Yan, Y.; Sun, J. Functional characterization of duck STING in IFN-β induction and Anti-H9N2 avian influenza viruses infections. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, W.; Wu, Y.; Wang, T.; Wu, S.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Zhao, X.; et al. Binding of the Duck Tembusu Virus Protease to STING Is Mediated by NS2B and Is Crucial for STING Cleavage and for Impaired Induction of IFN-β. J. Immunol. 2019, 203, 3374–3385. [Google Scholar] [CrossRef]
- Zhang, Z.; Yuan, B.; Bao, M.; Lu, N.; Kim, T.; Liu, Y.-J. The helicase DDX41 senses intracellular DNA mediated by the adaptor STING in dendritic cells. Nat. Immunol. 2011, 12, 959–965. [Google Scholar] [CrossRef]
- Parvatiyar, K.; Zhang, Z.; Teles, R.M.; Ouyang, S.; Jiang, Y.; Iyer, S.S.; Zaver, S.A.; Schenk, M.; Zeng, S.; Zhong, W.; et al. The helicase DDX41 recognizes the bacterial secondary messengers cyclic di-GMP and cyclic di-AMP to activate a type I interferon immune response. Nat. Immunol. 2012, 13, 1155–1161. [Google Scholar] [CrossRef]
- Cheng, Y.; Liu, Y.; Wang, Y.; Niu, Q.; Gao, Q.; Fu, Q.; Ma, J.; Wang, H.; Yan, Y.; Ding, C.; et al. Chicken DNA virus sensor DDX41 activates IFN-β signaling pathway dependent on STING. Dev. Comp. Immunol. 2017, 76, 334–342. [Google Scholar] [CrossRef]
- Li, Y.; Li, H.; Su, N.; Liu, D.; Luo, R.; Jin, H. Molecular cloning and functional characterization of duck DDX41. Dev. Comp. Immunol. 2018, 88, 183–189. [Google Scholar] [CrossRef]
- Kim, T.; Pazhoor, S.; Bao, M.; Zhang, Z.; Hanabuchi, S.; Facchinetti, V.; Bover, L.; Plumas, J.; Chaperot, L.; Qin, J.; et al. Aspartate-glutamate-alanine-histidine box motif (DEAH)/RNA helicase A helicases sense microbial DNA in human plasmacytoid dendritic cells. Proc. Natl. Acad. Sci. USA 2010, 107, 15181–15186. [Google Scholar] [CrossRef]
- Sato, H.; Oshiumi, H.; Takaki, H.; Hikono, H.; Seya, T. Evolution of the DEAD box helicase family in chicken: Chickens have no DHX9 ortholog. Microbiol. Immunol. 2015, 59, 633–640. [Google Scholar] [CrossRef]
- Wang, L.; Wen, M.; Cao, X. Nuclear hnRNPA2B1 initiates and amplifies the innate immune response to DNA viruses. Science 2019, 365, eaav0758. [Google Scholar] [CrossRef]
- Persing, D.H.; Coler, R.N.; Lacy, M.J.; Johnson, D.A.; Baldridge, J.R.; Hershberg, R.M.; Reed, S.G. Taking toll: Lipid A mimetics as adjuvants and immunomodulators. Trends Microbiol. 2002, 10, s32–s37. [Google Scholar] [CrossRef]
- Kanzler, H.; Barrat, F.J.; Hessel, E.M.; Coffman, R.L. Therapeutic targeting of innate immunity with Toll-like receptor agonists and antagonists. Nature Med. 2007, 13, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Tesar, B.M.; Walker, W.E.; Goldstein, D.R. Dual signaling of MyD88 and TRIF is critical for maximal TLR4-induced dendritic cell maturation. J. Immunol. 2008, 181, 1849–1858. [Google Scholar] [CrossRef] [PubMed]
- McAleer, J.P.; Vella, A.T. Understanding how lipopolysaccharide impacts CD4 T-cell immunity. Crit. Rev. Immunol. 2008, 28, 281–299. [Google Scholar] [CrossRef] [PubMed]
- Mata-Haro, V.; Cekic, C.; Martin, M.; Chilton, P.M.; Casella, C.R.; Mitchell, T.C. The vaccine adjuvant monophosphoryl lipid A as a TRIF-biased agonist of TLR4. Science 2007, 316, 1628–1632. [Google Scholar] [CrossRef]
- Bavananthasivam, J.; Alkie, T.N.; Matsuyama-Kato, A.; Hodgins, D.C.; Sharif, S. Characterization of innate responses induced by in ovo administration of encapsulated and free forms of ligands of Toll-like receptor 4 and 21 in chicken embryos. Res. Vet. Sci. 2019, 125, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Barjesteh, N.; Brisbin, J.T.; Behboudi, S.; Nagy, E.; Sharif, S. Induction of antiviral responses against avian influenza virus in embryonated chicken eggs with toll-like receptor ligands. Viral Immunol. 2015, 28, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Bavananthasivam, J.; Alkie, T.N.; Astill, J.; Abdul-Careem, M.F.; Wootton, S.K.; Behboudi, S.; Yitbarek, A.; Sharif, S. In ovo administration of Toll-like receptor ligands encapsulated in PLGA nanoparticles impede tumor development in chickens infected with Marek’s disease virus. Vaccine 2018, 36, 4070–4076. [Google Scholar] [CrossRef]
- Bavananthasivam, J.; Read, L.; Astill, J.; Yitbarek, A.; Alkie, T.N.; Abdul-Careem, M.F.; Wootton, S.K.; Behboudi, S.; Sharif, S. The effects of in ovo administration of encapsulated Toll-like receptor 21 ligand as an adjuvant with Marek’s disease vaccine. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Jenkins, K.A.; Lowenthal, J.W.; Kimpton, W.; Bean, A.G. The in vitro and in ovo responses of chickens to TLR9 subfamily ligands. Dev. Comp. Immunol. 2009, 33, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Gunawardana, T.; Foldvari, M.; Zachar, T.; Popowich, S.; Chow-Lockerbie, B.; Ivanova, M.V.; Tikoo, S.; Kurukulasuriya, S.; Willson, P.; Gomis, S.; et al. Protection of neonatal broiler chickens following in ovo delivery of oligodeoxynucleotides containing CpG motifs (CpG-ODN) formulated with carbon nanotubes or liposomes. Avian Dis. 2015, 59, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Cader, M.S.; Amarasinghe, A.; Palomino-Tapia, V.; Ahmed-Hassan, H.; Bakhtawar, K.; Nagy, E.; Sharif, S.; Gomis, S.; Abdul-Careem, M.F. In ovo CpG DNA delivery increases innate and adaptive immune cells in respiratory, gastrointestinal and immune systems post-hatch correlating with lower infectious laryngotracheitis virus infection. PLoS ONE 2018, 13, e0193964. [Google Scholar] [CrossRef] [PubMed]
- De Silva Senapathi, U.; Abdul-Cader, M.S.; Amarasinghe, A.; van Marle, G.; Czub, M.; Gomis, S.; Abdul-Careem, M.F. The in ovo delivery of CpG oligonucleotides protects against infectious bronchitis with the recruitment of immune cells into the respiratory tract of chickens. Viruses 2018, 10. [Google Scholar] [CrossRef]
- Dalloul, R.A.; Lillehoj, H.S.; Okamura, M.; Xie, H.; Min, W.; Ding, X.; Heckert, R.A. In vivo effects of CpG oligodeoxynucleotide on Eimeria infection in chickens. Avian Dis. 2004, 48, 783–790. [Google Scholar] [CrossRef]
- Patel, B.A.; Gomis, S.; Dar, A.; Willson, P.J.; Babiuk, L.A.; Potter, A.; Mutwiri, G.; Tikoo, S.K. Oligodeoxynucleotides containing CpG motifs (CpG-ODN) predominantly induce Th1-type immune response in neonatal chicks. Dev. Comp. Immunol. 2008, 32, 1041–1049. [Google Scholar] [CrossRef]
- Parvizi, P.; Abdul-Careem, M.F.; Mallick, A.I.; Haq, K.; Haghighi, H.R.; Orouji, S.; Heidari, M.; Behboudi, S.; Sharif, S. The effects of administration of ligands for Toll-like receptor 4 and 21 against Marek’s disease in chickens. Vaccine 2014, 32, 1932–1938. [Google Scholar] [CrossRef]
- Li, Y.; Meng, F.; Cui, S.; Fu, J.; Wang, Y.; Cui, Z.; Chang, S.; Zhao, P. Cooperative effects of immune enhancer TPPPS and different adjuvants on antibody responses induced by recombinant ALV-J gp85 subunit vaccines in SPF chickens. Vaccine 2017, 35, 1594–1598. [Google Scholar] [CrossRef] [PubMed]
- Dar, A.; Tikoo, S.; Potter, A.; Babiuk, L.A.; Townsend, H.; Gerdts, V.; Mutwiri, G. CpG-ODNs induced changes in cytokine/chemokines genes expression associated with suppression of infectious bronchitis virus replication in chicken lungs. Vet. Immunol. Immunopathol. 2014, 160, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Parvizi, P.; Mallick, A.I.; Haq, K.; Haghighi, H.R.; Orouji, S.; Thanthrige-Don, N.; Paul, M.; Brisbin, J.T.; Read, L.R.; Behboudi, S.; et al. A toll-like receptor 3 ligand enhances protective effects of vaccination against Marek’s disease virus and hinders tumor development in chickens. Viral Immunol. 2012, 25, 394–401. [Google Scholar] [CrossRef]
- Genovese, K.J.; He, H.; Lowry, V.K.; Nisbet, D.J.; Kogut, M.H. Dynamics of the avian inflammatory response to Salmonella following administration of the toll-like receptor 5 agonist flagellin. FEMS Immunol. Med. Microbiol. 2007, 51, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Chaung, H.-C.; Cheng, L.-T.; Hung, L.-H.; Tsai, P.-C.; Skountzou, I.; Wang, B.; Compans, R.W.; Lien, Y.-Y. Salmonella flagellin enhances mucosal immunity of avian influenza vaccine in chickens. Vet. Microbiol. 2012, 157, 69–77. [Google Scholar] [CrossRef]


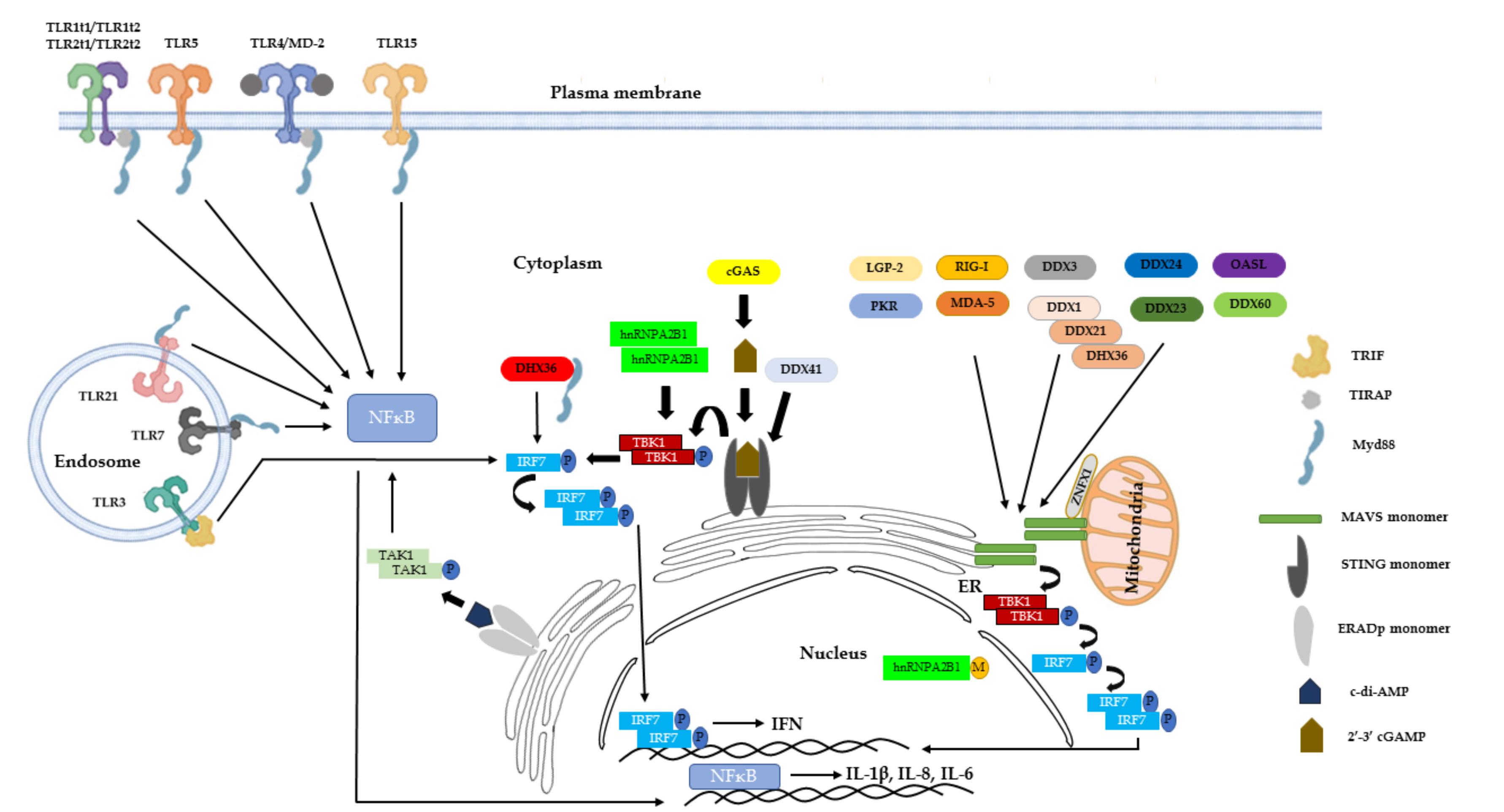{kind=link}
| Pattern Recognition Receptor (PRR) Class | Human | Chicken | Duck | Goose | Ligand and/or Function |
|---|---|---|---|---|---|
| Toll-like receptor (TLR) | TLR2/TLR1 | TLR2t1/TLR1t1, TLR2t1/TLR1t2, TLR2t2/TLR1t1 | TLR2t1/TLR1t1, TLR2t1/TLR1t2, TLR2t2/TLR1t1 | TLR2/TLR1 | Tri-acylated lipopetides |
| TLR2/TLR6 TLR2t2/TLR16, TLR2t1/TLR1LB | TLR2t2/TLR1t1, TLR2t1/TLR1t2 | TLR2t2/TLR1t1, TLR2t1/TLR1t2 | TLR2/TLR1 | Di-acylated lipopetides | |
| TLR3 | TLR3 | TLR3 | TLR3 | dsRNA | |
| TLR4/MD-2 | TLR4/MD-2 | TLR4/MD-2 | TLR4/MD-2 | LPS | |
| TLR5 | TLR5 | TLR5 | TLR5 | Flagellin | |
| TLR7 | TLR7 | TLR7 | TLR7 | ssRNA | |
| TLR8 | non-functional | non-functional | non-functional | ssRNA | |
| TLR9 | TLR21 | TLR21 | TLR21 | DNA | |
| Absent | TLR15 | TLR15 | TLR15 | Protease | |
| DEAD/H box helicase | RIG-I | Absent | RIG-I | RIG-I | 5’-ppp, short dsRNA |
| MDA5 | MDA5 | MDA5 | MDA5 | long dsRNA; chMDA5 also senses and responds to short dsRNA | |
| LGP2 | LGP2 | LGP2 | LGP2 | dsRNA and RIG-I/MDA5 regulation | |
| DDX1/DDX21/DHX36 | DDX1/DDX21/DHX36 | DDX1/DDX21/DHX36 | DDX1/DDX21/DHX36 | dsRNA | |
| DDX3 | DDX3 | DDX3 | DDX3 | dsRNA | |
| DDX23 | DDX23 | DDX23 | DDX23 | dsRNA | |
| DDX24 | DDX24 | DDX24 | DDX24 | dsRNA | |
| DDX60 | DDX60 | DDX60 | DDX60 | RIG-I sentinel and RNA degradation via exosome | |
| DDX41 | DDX41 | DDX41 | DDX41 | dsDNA | |
| DHX36 | DHX36 | DHX36 | DHX36 | CpG-A [ODN2216] DNA | |
| DHX9 | Absent | Absent | Absent | CpG-B [ODN2006] DNA | |
| Template independent nucleotidyl transferases | OAS1/2/3 | OASL | OASL | OASL | dsRNA |
| cGAS | cGAS | cGAS | cGAS | dsDNA | |
| Cyclic di-nucleotide sensor | STING | STING | STING | STING | 2’-3’ cGAMP |
| ERADp | ERADp | ERADp | ERADp | c-di-AMP | |
| RECON | Absent | Absent | Absent | c-di-AMP; NFκB negative regulator | |
| Other PRRs | AIM2 | Absent | Absent | Absent | dsDNA |
| IFI16 | Absent | Absent | Absent | dsDNA | |
| ZBP1/DAI | Absent | Absent | Absent | dsDNA | |
| HnRNPA2B1 | HnRNPA2B1 | HnRNPA2B1 | HnRNPA2B1 | dsDNA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neerukonda, S.N.; Katneni, U. Avian Pattern Recognition Receptor Sensing and Signaling. Vet. Sci. 2020, 7, 14. https://doi.org/10.3390/vetsci7010014
Neerukonda SN, Katneni U. Avian Pattern Recognition Receptor Sensing and Signaling. Veterinary Sciences. 2020; 7(1):14. https://doi.org/10.3390/vetsci7010014
Chicago/Turabian StyleNeerukonda, Sabari Nath, and Upendra Katneni. 2020. "Avian Pattern Recognition Receptor Sensing and Signaling" Veterinary Sciences 7, no. 1: 14. https://doi.org/10.3390/vetsci7010014
APA StyleNeerukonda, S. N., & Katneni, U. (2020). Avian Pattern Recognition Receptor Sensing and Signaling. Veterinary Sciences, 7(1), 14. https://doi.org/10.3390/vetsci7010014