Diversity of Diptera Species in Estonian Pig Farms

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
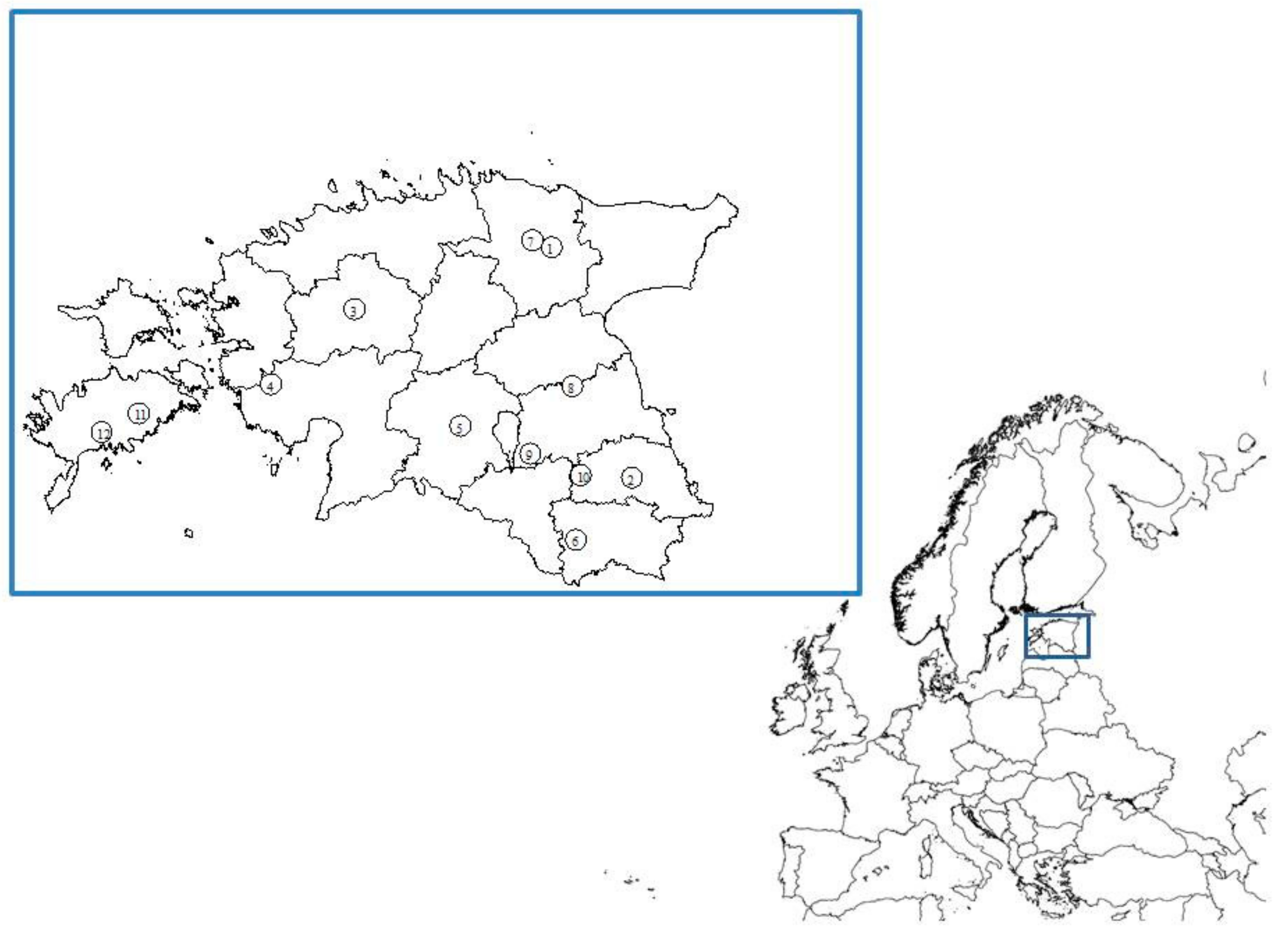
2.1. Sampling
2.2. Questionnaire
2.3. Species Determination Based on Mitochondrial DNA
2.4. Data Analysis
3. Results
3.1. Farm Characteristics
3.2. Collected Species and Their Abundance
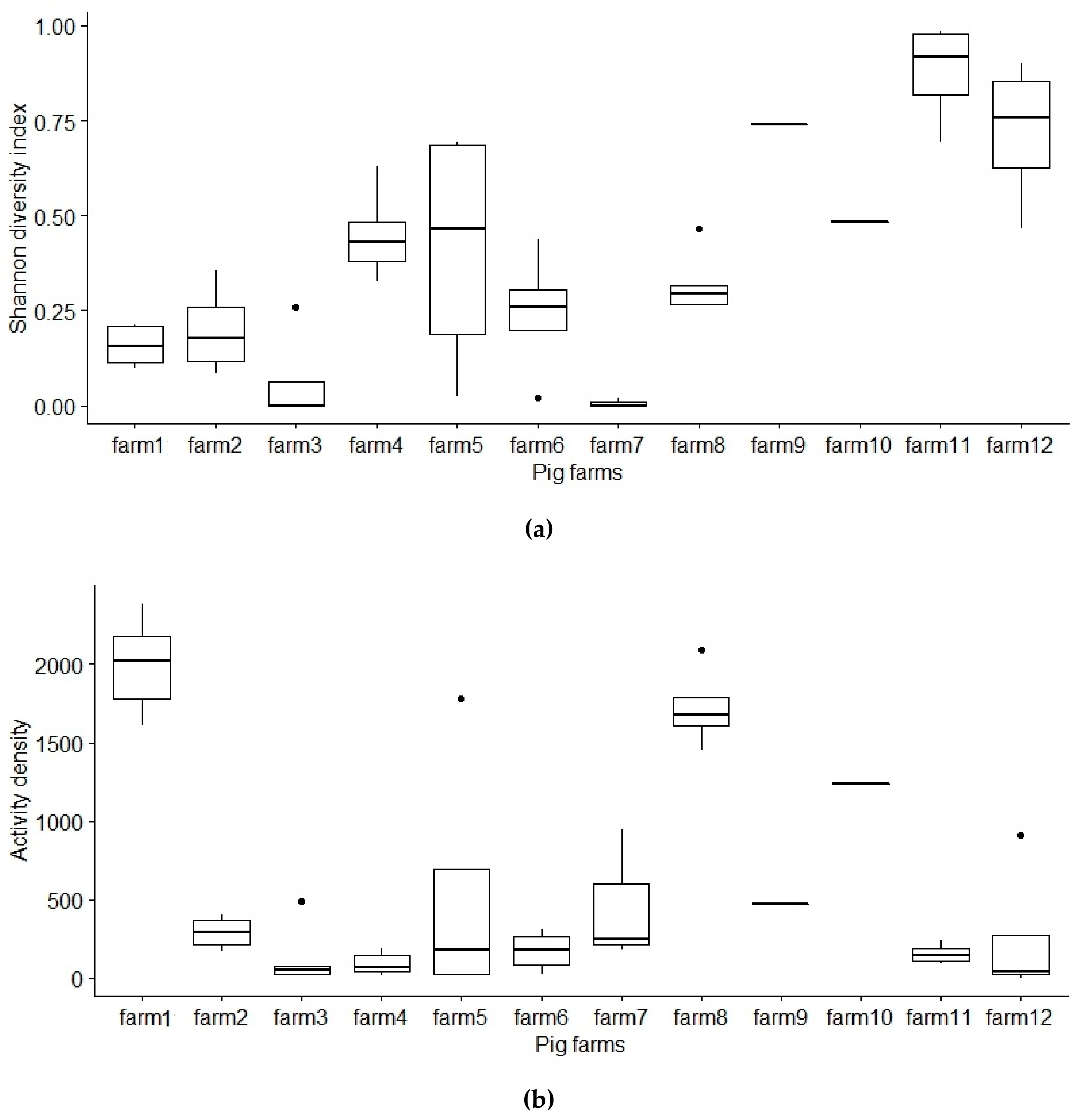
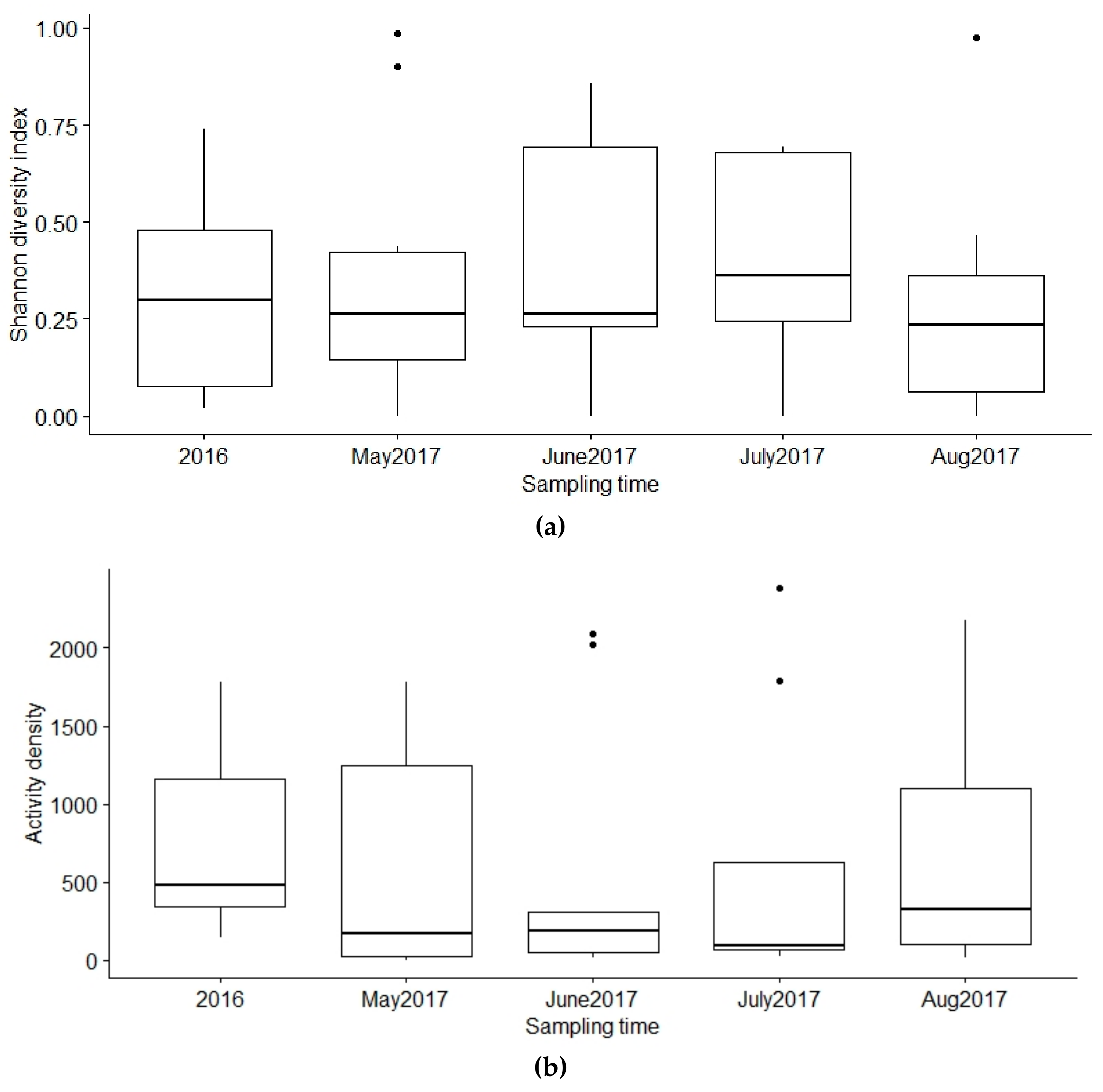
3.3. Activity Density
3.4. Diversity of Diptera in Farms
3.5. Parameters Affecting the Activity Density of Diptera
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Birkemoe, T.; Sverdrup-Thygeson, A. Stable fly (Stomoxys calcitrans) and house fly (Musca domestica) densities: A comparison of three monitoring methods on pig farms. J. Pest. Sci. 2011, 84, 273–280. [Google Scholar] [CrossRef]
- Malik, A.; Singh, N.; Satya, S. House fly (Musca domestica): A review of control strategies for a challenging pest. J. Environ. Sci. Heal. B. 2007, 42, 453–469. [Google Scholar] [CrossRef] [PubMed]
- Adler, P.H.; Tuten, H.C.; Nelder, M.P. Arthropods of medicoveterinary importance in zoos. Annu. Rev. Entomol. 2011, 56, 123–142. [Google Scholar] [CrossRef] [PubMed]
- Graczyk, T.K.; Knight, R.; Gilman, R.H.; Cranfield, M.R. The role of non-biting flies in the epidemiology of human infectious diseases. Microbes Infect. 2001, 3, 231–235. [Google Scholar] [CrossRef]
- Doyle, M.S.; Swope, B.N.; Hogsette, J.A.; Burkhalter, K.L.; Savage, H.M.; Nasci, R.S. Vector competence of the stable fly (Diptera: Muscidae) for west nile virus. J. Med. Entomol. 2011, 48, 656–668. [Google Scholar] [CrossRef]
- Mramba, F.; Broce, A.B.; Zurek, L. Vector competence of stable flies, Stomoxys calcitrans L. (Diptera: Muscidae), for Enterobacter sakazakii. J. Vector Ecol. 2007, 32, 134–139. [Google Scholar] [CrossRef]
- Fischer, O.; Matlova, L.; Dvorska, L.; Svastova, P.; Bartl, J.; Melicharek, I.; Weston, R.T.; Pavlik, I. Diptera as vectors of mycobacterial infections in cattle and pigs. Med. Vet. Entomol. 2001, 15, 208–211. [Google Scholar] [CrossRef]
- Khamesipour, F.; Lankarani, K.B.; Honarvar, B.; Kwenti, T.E. A systematic review of human pathogens carried by the housefly (Musca domestica L.). BMC Public Health 2018, 18. [Google Scholar] [CrossRef]
- Nayduch, D.; Burrus, R.G. Flourishing in filth: House fly-microbe interactions across life history. Ann. Entomol. Soc. Am. 2017, 110, 6–18. [Google Scholar] [CrossRef]
- Albuquerque, T.A.; Zurek, L. Temporal changes in the bacterial community of animal feces and their correlation with stable fly oviposition, larval development, and adult fitness. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef]
- Issa, R. Musca domestica acts as transport vector hosts. Bull. Nat. Res. Centre 2019, 43, 73. [Google Scholar] [CrossRef]
- Olesen, A.S.; Lohse, L.; Hansen, M.F.; Boklund, A.; Halasa, T.; Belsham, G.J.; Rasmussen, T.B.; Botner, A.; Bodker, R. Infection of pigs with African swine fever virus via ingestion of stable flies (Stomoxys calcitrans). Transbound. Emerg. Dis. 2018, 65, 1152–1157. [Google Scholar] [CrossRef] [PubMed]
- Baldacchino, F.; Muenworn, V.; Desquesnes, M.; Desoli, F.; Charoenviriyaphap, T.; Duvallet, G. Transmission of pathogens by Stomoxys flies (Diptera, Muscidae): A review. Parasite 2013, 20. [Google Scholar] [CrossRef] [PubMed]
- Nurmoja, I.; Mõtus, K.; Kristian, M.; Niine, T.; Schulz, K.; Depner, K.; Viltrop, A. Epidemiological analysis of the 2015-2017 African swine fever outbreaks in Estonia. Prev. Vet. Med. 2018. [Google Scholar] [CrossRef]
- Nurmoja, I.; Schulz, K.; Staubach, C.; Sauter-Louis, C.; Depner, K.; Conraths, F.J.; Viltrop, A. Development of African swine fever epidemic among wild boar in Estonia—two different areas in the epidemiological focus. Sci. Rep. 2017, 7, 12562. [Google Scholar] [CrossRef]
- Alvarez, J.; Bicout, D.; Boklund, A.; Butner, A.; Depner, K.; More, S.J.; Roberts, H.; Stahl, K.; Thulke, H.H.; Viltrop, A.; et al. Research gap analysis on African swine fever. Efsa J. 2019, 17, 60. [Google Scholar] [CrossRef]
- Mellor, P.S.; Kitching, R.P.; Wilkinson, P.J. Mechanical transmission of capribox virus and African swine fever virus by Stomyxos calcitrans. Res. Vet. Sci. 1987, 43, 109–112. [Google Scholar] [CrossRef]
- Herm, R.; Tummeleht, L.; Jürison, M.; Vilem, A.; Viltrop, A. Trace amounts of African swine fever virus DNA detected in insects collected from an infected pig farm in Estonia. Vet. Med. Sci. 2019. [Google Scholar] [CrossRef]
- Lilja, T.; Troell, K.; Kirik, H.; Lindstrom, A. A distinct group of north European Aedes vexans as determined by mitochondrial and nuclear markers. Med. Vet. Entomol. 2018, 32, 282–289. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Hebert, P.D.N.; Ratnasingham, S.; DeWaard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Biol. Sci. 2003, 270, S96–S99. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Carew, M.E.; Pettigrove, V.; Hoffmann, A.A. The utility of DNA markers in classical taxonomy: Using cytochrome oxidase I markers to differentiate Australian Cladopelma (Diptera: Chironomidae) midges. Ann. Entomol. Soc. Am. 2005, 98, 587–594. [Google Scholar] [CrossRef]
- Jürgenstein, S.; Kurina, O.; Põldmaa, K. The Mycetophila ruficollis Meigen (Diptera, Mycetophilidae) group in Europe: Elucidating species delimitation with COI and ITS2 sequence data. Zookeys 2015, 508, 15–51. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence slignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Gardiner, M.M.; Landis, D.A.; Gratton, C.; Schmidt, N.; O’Neal, M.; Mueller, E.; Chacon, J.; Heimpel, G.E. Landscape composition influences the activity density of Carabidae and Arachnida in soybean fields. Biol. Control. 2010, 55, 11–19. [Google Scholar] [CrossRef]
- Yi, Z.; Jinchao, F.; Dayuan, X.; Weiguo, S.; Axmacher, J.C. A comparison of terrestrial arthropod sampling methods. J. Res. Ecol. 2012, 3, 174–182, 179. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Gerhardt, R.R.; Hribar, L.J. Chapter 11—Flies (Diptera). In Medical and Veterinary Entomology (Third Edition); Mullen, G.R., Durden, L.A., Eds.; Academic Press:: Cambridge, MA, USA, 2019; pp. 171–190. [Google Scholar] [CrossRef]
- Forster, M.; Sievert, K.; Messler, S.; Klimpel, S.; Pfeffer, K. Comprehensive study on the occurrence and distribution of pathogenic microorganisms carried by synanthropic flies caught at different rural locations in Germany. J. Med. Entomol. 2009, 46, 1164–1166. [Google Scholar] [CrossRef]
- Rozkošný, R.; Gregor, F.; Barták, M.; Vaňhara, J. The Muscidae (Diptera) of Central Europe; Masaryk University: Brno, Czech Republic, 2002; p. 107. [Google Scholar]
- Skidmore, P. The Biology of the Muscidae of the World; Springer: Haarlem, The Netherlands, 1985; pp. 112 and 201. [Google Scholar]
- McOrist, S.; Blunt, R.; Gebhart, C.J. Pig-associated Lawsonia intracellularis in various on-farm dipterous fly stages. J. Swine Health Prod. 2011, 19, 277–283. [Google Scholar]
- Chvála, M.; Lyneborg, L.; Moucha, J. The Horse Flies of Europe: (Diptera, Tabanidae); Entomological Society of Copenhagen: Copenhagen, Denmark, 1972; pp. 38–40. [Google Scholar]
- Baldacchino, F.; Desquesnes, M.; Mihok, S.; Foil, L.D.; Duvallet, G.; Jittapalapong, S. Tabanids: Neglected subjects of research, but important vectors of disease agents! Infect. Genet. Evol. 2014, 28, 596–615. [Google Scholar] [CrossRef]
- Becker, N.; Petric, D.; Zgomba, M.; Boase, C.; Dahl, C.; Madon, M.; Kaiser, A. Mosquitoes and Their Control., Second Edition; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1–577. [Google Scholar]
- Brugman, V.A.; Medlock, J.M.; Logan, J.G.; Wilson, A.J.; Lindsay, S.W.; Fooks, A.R.; Mertens, P.P.C.; Johnson, N.; Carpenter, S.T. Bird-biting mosquitoes on farms in southern England. Vet. Rec. 2018, 183, 474. [Google Scholar] [CrossRef] [PubMed]
- Boukraa, S.; De la Grandiere, M.A.; Bawin, T.; Raharimalala, F.N.; Zimmer, J.Y.; Haubruge, E.; Thiry, E.; Francis, F. Diversity and ecology survey of mosquitoes potential vectors in Belgian equestrian farms: A threat prevention of mosquito-borne equine arboviruses. Prev. Vet. Med. 2016, 124, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Hameed, M.; Liu, K.; Anwar, M.N.; Wahaab, A.; Li, C.X.; Di, D.; Wang, X.; Khan, S.; Xu, J.P.; Li, B.B.; et al. A viral metagenomic analysis reveals rich viral abundance and diversity in mosquitoes from pig farms. Transbound. Emerg. Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Otake, S.; Dee, S.A.; Rossow, K.D.; Moon, R.D.; Pijoan, C. Mechanical transmission of porcine reproductive and respiratory syndrome virus by mosquitoes, Aedes vexans (Meigen). Can. J. Vet. Res. 2002, 66, 191–195. [Google Scholar]
- Nazni, W.A.; Luke, H.; Wan Rozita, W.M.; Abdullah, A.G.; Sa’diyah, I.; Azahari, A.H.; Zamree, I.; Tan, S.B.; Lee, H.L.; Sofian, M.A. Determination of the flight range and dispersal of the house fly, Musca domestica (L.) using mark release recapture technique. Trop. Biomed. 2005, 22, 53–61. [Google Scholar] [PubMed]
- Beresford, D.V.; Sutcliffe, J.F. Local infestation or long-distance migration? The seasonal recolonization of dairy farms by Stomoxys calcitrans (Diptera: Muscidae) in South Central Ontario, Canada. J. Econ. Entomol. 2009, 102, 788–798. [Google Scholar] [CrossRef]
- Hogsette, J.A.; Ruff, J.P. Stable fly (Diptera, Muscidae) migration in Northwest Florida. Envir. Entomol. 1985, 14, 170–175. [Google Scholar] [CrossRef]
- Schoof, H.F.; Siverly, R.E. Multiple Release Studies on the Dispersion of Musca domestica at Phoenix, Arizona. J. Econ. Entomol. 1954, 47, 830–838. [Google Scholar] [CrossRef]
- Lempereur, L.; Sohier, C.; Smeets, F.; Marechal, F.; Berkvens, D.; Madder, M.; Francis, F.; Losson, B. Dispersal capacity of Haematopota spp. and Stomoxys calcitrans using a mark-release-recapture approach in Belgium. Med. Vet. Entomol. 2018, 32, 298–303. [Google Scholar] [CrossRef]
- Zurek, L.; Ghosh, A. Insects represent a link between food animal farms and the urban environment for antibiotic resistance traits. Appl. Environ. Microbiol. 2014, 80, 3562–3567. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Kambhampati, S.; Zurek, L. Assessment of house fly dispersal between rural and urban habitats in Kansas, USA. J. Kansas Entomol. Soc. 2010, 83, 172–188. [Google Scholar] [CrossRef]
- Skovgard, H.; Nachman, G. Population dynamics of stable flies Stomoxys calcitrans (Diptera: Muscidae) at an organic dairy farm in Denmark based on mark-recapture with destructive sub-sampling. Environ. Entomol. 2012, 41, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Estonian Weather Service. Available online: http://www.ilmateenistus.ee/kliima/climate-maps/temperature/?lang=en/ (accessed on 19 November 2019).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/Groups | Percentage of Total No. of Individuals (%) | Number of Individuals | Number of Farms Where Found |
|---|---|---|---|
| Drosophilidae spp. | 6.6 | 12,306 | 12 |
| Muscidae | |||
| Fannia canicularis (Linnaeus, 1761) | 0.0005 | 1 | 1 |
| Hydrotaea dentipes (Fabricius, 1805) | 0.3 | 541 | 7 |
| Musca domestica Linnaeus, 1758 | 88.5 | 165,203 | 12 |
| Pyrellia vivida Robineau-Desvoidy, 1830 | 0.002 | 4 | 2 |
| Stomoxys calcitrans (Linnaeus, 1758) | 1.2 | 2156 | 5 |
| Calliphoridae | |||
| Pollenia rudis (Fabricius, 1794) | 0.0005 | 1 | 1 |
| Sepsidae | |||
| Sepsis violacea Meigen, 1826 | 0.01 | 20 | 1 |
| Tabanidae | |||
| Haematopota pluvialis (Linnaeus, 1758) | 0.002 | 4 | 2 |
| Chrysops (Chrysops) relictus Meigen, 1820 | 0.0005 | 1 | 1 |
| Culicidae | |||
| Aedes (Aedimorphus) vexans (Meigen, 1830) | 0.0005 | 1 | 1 |
| Anopheles (Anopheles) maculipennis s.l. Meigen, 1818 | 0.002 | 4 | 2 |
| Coquillettidia (Coquillettidia) richiardii (Ficalbi, 1889) | 0.0005 | 1 | 1 |
| Culex (Culex) pipiens s.l. Linnaeus, 1758 | 0.006 | 11 | 4 |
| Ochlerotatus (Ochlerotatus) annulipes (Meigen, 1830) | 0.0005 | 1 | 1 |
| Ochlerotatus (Ochlerotatus) cataphylla (Dyar, 1916) | 0.01 | 20 | 3 |
| Ochlerotatus (Ochlerotatus) intrudens (Dyar, 1919) | 0.001 | 2 | 1 |
| Ochlerotatus (Ochlerotatus) pullatus (Coquillett, 1904) | 0.0016 | 3 | 3 |
| Ochlerotatus (Ochlerotatus) punctor (Kirby, 1837) | 0.03 | 67 | 6 |
| Ochlerotatus (Rusticoidus) rusticus (Rossi, 1790) | 0.001 | 2 | 1 |
| Ochlerotatus (Ochlerotatus) sticticus (Meigen, 1838) | 0.0016 | 3 | 3 |
| Others | |||
| Diptera spp. a | 0.05 | 94 | 9 |
| Arthropoda spp. b | 3.4 | 6257 | 12 |
| In total | 186,703 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tummeleht, L.; Jürison, M.; Kurina, O.; Kirik, H.; Jeremejeva, J.; Viltrop, A. Diversity of Diptera Species in Estonian Pig Farms. Vet. Sci. 2020, 7, 13. https://doi.org/10.3390/vetsci7010013
Tummeleht L, Jürison M, Kurina O, Kirik H, Jeremejeva J, Viltrop A. Diversity of Diptera Species in Estonian Pig Farms. Veterinary Sciences. 2020; 7(1):13. https://doi.org/10.3390/vetsci7010013
Chicago/Turabian StyleTummeleht, Lea, Margret Jürison, Olavi Kurina, Heli Kirik, Julia Jeremejeva, and Arvo Viltrop. 2020. "Diversity of Diptera Species in Estonian Pig Farms" Veterinary Sciences 7, no. 1: 13. https://doi.org/10.3390/vetsci7010013
APA StyleTummeleht, L., Jürison, M., Kurina, O., Kirik, H., Jeremejeva, J., & Viltrop, A. (2020). Diversity of Diptera Species in Estonian Pig Farms. Veterinary Sciences, 7(1), 13. https://doi.org/10.3390/vetsci7010013