2,4-Thiazolidinedione in Well-Fed Lactating Dairy Goats: II. Response to Intra-Mammary Infection



 ,
, 
Abstract
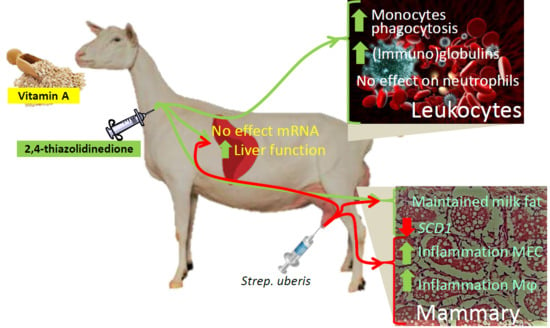
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Animal Management
- -
- CTR-control, untouched left mammary gland from goats receiving an intrajugular injection of saline;
- -
- CTR-IMI, right mammary gland infused with Strep uberis from goats receiving an intrajugular injection of saline;
- -
- TZD-control, untouched left mammary gland from goats receiving an intrajugular injection of 2,4-TZD;
- -
- TZD-IMI, right mammary gland infused with Strep uberis from goats receiving an intrajugular injection of 2,4-TZD.
2.2. Measurements, Sample Collection, and Blood Metabolites
2.3. Blood Metabolites and Inflammatory Markers
2.4. Phagocytosis and Percent Leukocytes
2.5. Macrophages Isolation from Milk
2.6. Euthanasia and Tissue Collection
2.7. RT-qPCR
2.8. Statistical Analysis
3. Results
3.1. Body Temperature and Feed Intake
3.2. Milk yield, SCC and Milk Composition
3.3. Blood Biomarkers
3.4. Differential Leukocyte Count and Phagocytosis
3.5. Gene Expression in Liver and MEC
3.6. Gene Expression in Macrophages
4. Discussion
4.1. Response to IMI in Comparison with the Prior Study
4.2. Treatment with 2,4-TZD Allows Maintaining Milk Fat Synthesis after IMI but Not through SCD1
4.3. Treatment with 2,4-TZD Modestly Improves the Liver Response to Mammary Infection
4.4. Treatment with 2,4-Thiazolidinedione Increases Plasma Globulin and Increases Monocyte Phagocytosis
4.5. Treatment with 2,4-TZD has a Moderate Effect on Oxidative Parameters
4.6. IMI but Not 2,4-TZD Affects the Transcriptome of MEC and Macrophages
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rinaldi, M.; Li, R.W.; Capuco, A.V. Mastitis associated transcriptomic disruptions in cattle. Vet. Immunol. Immunopathol. 2010, 138, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.M. Understanding the Basics of Mastitis. 2009. Available online: https://vtechworks.lib.vt.edu/bitstream/handle/10919/48392/404-233_pdf.pdf?sequence=1 (accessed on 1 June 2019).
- Erskine, R.J.; Walker, R.D.; Bolin, C.A.; Bartlett, P.C.; White, D.G. Trends in antibacterial susceptibility of mastitis pathogens during a seven-year period. J. Dairy Sci. 2002, 85, 1111–1118. [Google Scholar] [CrossRef]
- Heinrichs, A.J.; Costello, S.S.; Jones, C.M. Control of heifer mastitis by nutrition. Vet. Microbiol. 2009, 134, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Erskine, R.J. Nutrition and mastitis. Vet. Clin. N. Am. Food Anim. Pract. 1993, 9, 551–561. [Google Scholar] [CrossRef]
- LeBlanc, S.J.; Herdt, T.H.; Seymour, W.M.; Duffield, T.F.; Leslie, K.E. Peripartum serum vitamin E, retinol, and beta-carotene in dairy cattle and their associations with disease. J. Dairy Sci. 2004, 87, 609–619. [Google Scholar] [CrossRef]
- Sordillo, L.M.; Shafer-Weaver, K.; DeRosa, D. Immunobiology of the mammary gland. J. Dairy Sci. 1997, 80, 1851–1865. [Google Scholar] [CrossRef]
- Bionaz, M.; Osorio, J.; Loor, J.J. TRIENNIAL LACTATION SYMPOSIUM: Nutrigenomics in dairy cows: Nutrients, transcription factors, and techniques. J. Anim. Sci. 2015, 93, 5531–5553. [Google Scholar] [CrossRef] [PubMed]
- Bionaz, M.; Chen, S.; Khan, M.J.; Loor, J.J. Functional Role of PPARs in Ruminants: Potential Targets for Fine-Tuning Metabolism during Growth and Lactation. PPAR Res. 2013, 2013, 684159. [Google Scholar] [CrossRef]
- Yessoufou, A.; Wahli, W. Multifaceted roles of peroxisome proliferator-activated receptors (PPARs) at the cellular and whole organism levels. Swiss Med. Wkly. 2010, 140, w13071. [Google Scholar] [CrossRef]
- Rosa, F.; Osorio, J.S.; Trevisi, E.; Yanqui-Rivera, F.; Estill, C.T.; Bionaz, M. 2,4-Thiazolidinedione Treatment Improves the Innate Immune Response in Dairy Goats with Induced Subclinical Mastitis. PPAR Res. 2017, 2017, 7097450. [Google Scholar] [CrossRef] [PubMed]
- Jaaf, S.; Rosa, F.; Moridi, M.; Osorio, J.; Lohakare, J.; Trevisi, E.; Filley, S.; Estill, C.T.; Cherian, G.; Bionaz, M. 2,4-Thiazolidinedione in Well-Fed Lactating Dairy Goats: I. Effect on Adiposity and Milk Fat Synthesis. Vet. Sci. 2019, 6, 45. [Google Scholar] [CrossRef] [PubMed]
- Bionaz, M.; Trevisi, E.; Calamari, L.; Librandi, F.; Ferrari, A.; Bertoni, G. Plasma paraoxonase, health, inflammatory conditions, and liver function in transition dairy cows. J. Dairy Sci. 2007, 90, 1740–1750. [Google Scholar] [CrossRef]
- Trevisi, E.; Amadori, M.; Cogrossi, S.; Razzuoli, E.; Bertoni, G. Metabolic stress and inflammatory response in high-yielding, periparturient dairy cows. Res. Vet. Sci. 2012, 93, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Richoux, R.; Perruchot, M.H.; Boutinaud, M.; Mayol, J.F.; Gagnaire, V. Flow Cytometry Approach to Quantify the Viability of Milk Somatic Cell Counts after Various Physico-Chemical Treatments. PLoS ONE 2015, 10, e0146071. [Google Scholar] [CrossRef] [PubMed]
- Chawla, A. Control of macrophage activation and function by PPARs. Circ. Res. 2010, 106, 1559–1569. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3. [Google Scholar] [CrossRef]
- Knez, M.; Graham, R.D.; Welch, R.M.; Stangoulis, J.C. New perspectives on the regulation of iron absorption via cellular zinc concentrations in humans. Crit. Rev. Food Sci. Nutr. 2017, 57, 2128–2143. [Google Scholar] [CrossRef] [PubMed]
- Hoeben, D.; Burvenich, C.; Trevisi, E.; Bertoni, G.; Hamann, J.; Bruckmaier, R.M.; Blum, J.W. Role of endotoxin and TNF-alpha in the pathogenesis of experimentally induced coliform mastitis in periparturient cows. J. Dairy Res. 2000, 67, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Fasulkov, I.; Karadaev, M.; Vasilev, N.; Urumova, V.; Mircheva, T. Determination of plasma fibrinogen and haptoglobin, hematological and blood biochemical changes in Bulgarian local goats with experimentally induced Staphylococcus aureus mastitis. Turk. J. Vet. Anim. Sci. 2014, 38, 439–444. [Google Scholar] [CrossRef]
- Jankelson, I.R.; Gargill, S.L. Bilirubin Liver Function Test. N. Engl. J. Med. 1931, 204, 547–549. [Google Scholar] [CrossRef]
- Bertoni, G.; Trevisi, E. Use of the liver activity index and other metabolic variables in the assessment of metabolic health in dairy herds. Vet. Clin. Food Anim. Pract. 2013, 29, 413–431. [Google Scholar] [CrossRef] [PubMed]
- Busher, J.T. Serum Albumin and Globulin. Clinical Methods: The History, Physical, and Laboratory Examinations, 3rd ed.; Walker, H.K., Hall, W.D., Hurst, J.W., Eds.; Butterworths: Boston, MA, USA, 1990. [Google Scholar]
- O’Connell, T.X.; Horita, T.J.; Kasravi, B. Understanding and interpreting serum protein electrophoresis. Am. Fam. Physician 2005, 71, 105–112. [Google Scholar] [PubMed]
- Bobbo, T.; Ruegg, P.L.; Fiore, E.; Gianesella, M.; Morgante, M.; Pasotto, D.; Gallo, L.; Bittante, G.; Cecchinato, A. Short communication: Association between udder health status and blood serum proteins in dairy cows. J. Dairy Sci. 2017, 100, 9775–9780. [Google Scholar] [CrossRef] [PubMed]
- Croasdell, A.; Duffney, P.F.; Kim, N.; Lacy, S.H.; Sime, P.J.; Phipps, R.P. PPARgamma and the Innate Immune System Mediate the Resolution of Inflammation. PPAR Res. 2015, 2015, 549691. [Google Scholar] [CrossRef] [PubMed]
- Mimche, P.N.; Thompson, E.; Taramelli, D.; Vivas, L. Curcumin enhances non-opsonic phagocytosis of Plasmodium falciparum through up-regulation of CD36 surface expression on monocytes/macrophages. J. Antimicrob. Chemother. 2012, 67, 1895–1904. [Google Scholar] [CrossRef] [PubMed]
- Dale, D.C.; Boxer, L.; Liles, W.C. The phagocytes: Neutrophils and monocytes. Blood 2008, 112, 935–945. [Google Scholar] [CrossRef]
- Wellnitz, O.; Bruckmaier, R.M. The innate immune response of the bovine mammary gland to bacterial infection. Vet. J. 2012, 192, 148–152. [Google Scholar] [CrossRef]
- Buitenhuis, B.; Rontved, C.M.; Edwards, S.M.; Ingvartsen, K.L.; Sorensen, P. In depth analysis of genes and pathways of the mammary gland involved in the pathogenesis of bovine Escherichia coli-mastitis. BMC Genom. 2011, 12, 130. [Google Scholar] [CrossRef]
- Moyes, K.M.; Drackley, J.K.; Morin, D.E.; Bionaz, M.; Rodriguez-Zas, S.L.; Everts, R.E.; Lewin, H.A.; Loor, J.J. Gene network and pathway analysis of bovine mammary tissue challenged with Streptococcus uberis reveals induction of cell proliferation and inhibition of PPARgamma signaling as potential mechanism for the negative relationships between immune response and lipid metabolism. BMC Genom. 2009, 10, 542. [Google Scholar] [CrossRef]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Ricote, M.; Li, A.C.; Willson, T.M.; Kelly, C.J.; Glass, C.K. The peroxisome proliferator-activated receptor-gamma is a negative regulator of macrophage activation. Nature 1998, 391, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Ting, A.T.; Seed, B. PPAR-gamma agonists inhibit production of monocyte inflammatory cytokines. Nature 1998, 391, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Penas, F.; Mirkin, G.A.; Vera, M.; Cevey, A.; Gonzalez, C.D.; Gomez, M.I.; Sales, M.E.; Goren, N.B. Treatment in vitro with PPARalpha and PPARgamma ligands drives M1-to-M2 polarization of macrophages from T. cruzi-infected mice. Biochim. Biophys. Acta 2015, 1852, 893–904. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, T.; Koga, H.; Shimotohno, K. Transcriptional activity of peroxisome proliferator-activated receptor gamma is modulated by SUMO-1 modification. J. Biol. Chem. 2004, 279, 29551–29557. [Google Scholar] [CrossRef]
- Diezko, R.; Suske, G. Ligand binding reduces SUMOylation of the peroxisome proliferator-activated receptor gamma (PPARgamma) activation function 1 (AF1) domain. PLoS ONE 2013, 8, e66947. [Google Scholar] [CrossRef]
- Pascual, G.; Fong, A.L.; Ogawa, S.; Gamliel, A.; Li, A.C.; Perissi, V.; Rose, D.W.; Willson, T.M.; Rosenfeld, M.G.; Glass, C.K. A SUMOylation-dependent pathway mediates transrepression of inflammatory response genes by PPAR-gamma. Nature 2005, 437, 759–763. [Google Scholar] [CrossRef]
- Haschemi, A.; Chin, B.Y.; Jeitler, M.; Esterbauer, H.; Wagner, O.; Bilban, M.; Otterbein, L.E. Carbon monoxide induced PPARgamma SUMOylation and UCP2 block inflammatory gene expression in macrophages. PLoS ONE 2011, 6, e26376. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Group | Day from IMI | SEM 1 | p-Value 2 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 0.5 | 1 | 2 | 3 | 6 | 11 | Z | Time | Z × T | |||||
| Glucose (mM) | CTR | 3.57 | 3.86 | 3.47 | 3.58 | 3.32 | 3.48 | 3.47 | 0.10 | 0.79 | <0.01 | 0.58 | ||
| TZD | 3.57 | 3.72 | 3.56 | 3.52 | 3.36 | 3.50 | 3.33 | |||||||
| NEFA (mM) | CTR | 0.07 | 0.05 | 0.08 | 0.06 | 0.07 | 0.06 | 0.11 | 0.03 | 0.61 | <0.01 | 0.79 | ||
| TZD | 0.07 | 0.08 | 0.11 | 0.08 | 0.07 | 0.06 | 0.13 | |||||||
| TAG (mM) | CTR | 0.19 | 0.16 | 0.17 | 0.18 | 0.19 | 0.19 | 0.13 | 0.02 | 0.97 | <0.01 | 0.76 | ||
| TZD | 0.19 | 0.16 | 0.19 | 0.19 | 0.17 | 0.16 | 0.14 | |||||||
| BHBA (mM) | CTR | 0.41 | 0.43 | 0.38 | 0.43 | 0.42 | 0.38 | 0.32 | 0.03 | 0.81 | <0.01 | 0.27 | ||
| TZD | 0.41 | 0.40 | 0.43 | 0.41 | 0.35 | 0.36 | 0.34 | |||||||
| Urea (mM) | CTR | 5.97 | 6.20 | 6.81 | 5.61 | 6.54 | 6.45 | 8.05 | 0.57 | 0.78 | <0.01 | 0.35 | ||
| TZD | 5.97 | 6.65 | 6.77 | 5.76 | 6.30 | 6.40 | 6.90 | |||||||
| Cholesterol (mM) | CTR | 2.18 | 2.23 | 2.28 | 2.31 | 2.33 | 2.40 | 2.32 | 0.23 | 0.90 | <0.01 | 0.88 | ||
| TZD | 2.18 | 2.26 | 2.30 | 2.40 | 2.40 | 2.41 | 2.39 | |||||||
| Creatinine (μM) | CTR | 71.9 | 70.0 | 72.6 | 70.2 | 70.4 | 72.2 | 75.8 | 2.5 | 0.69 | <0.01 | 0.33 | ||
| TZD | 71.9 | 71.2 | 73.4 | 72.9 | 71.8 | 73.6 | 77.3 | |||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosa, F.; Moridi, M.; Osorio, J.S.; Lohakare, J.; Trevisi, E.; Filley, S.; Estill, C.; Bionaz, M. 2,4-Thiazolidinedione in Well-Fed Lactating Dairy Goats: II. Response to Intra-Mammary Infection. Vet. Sci. 2019, 6, 52. https://doi.org/10.3390/vetsci6020052
Rosa F, Moridi M, Osorio JS, Lohakare J, Trevisi E, Filley S, Estill C, Bionaz M. 2,4-Thiazolidinedione in Well-Fed Lactating Dairy Goats: II. Response to Intra-Mammary Infection. Veterinary Sciences. 2019; 6(2):52. https://doi.org/10.3390/vetsci6020052
Chicago/Turabian StyleRosa, Fernanda, Misagh Moridi, Johan S. Osorio, Jayant Lohakare, Erminio Trevisi, Shelby Filley, Charles Estill, and Massimo Bionaz. 2019. "2,4-Thiazolidinedione in Well-Fed Lactating Dairy Goats: II. Response to Intra-Mammary Infection" Veterinary Sciences 6, no. 2: 52. https://doi.org/10.3390/vetsci6020052
APA StyleRosa, F., Moridi, M., Osorio, J. S., Lohakare, J., Trevisi, E., Filley, S., Estill, C., & Bionaz, M. (2019). 2,4-Thiazolidinedione in Well-Fed Lactating Dairy Goats: II. Response to Intra-Mammary Infection. Veterinary Sciences, 6(2), 52. https://doi.org/10.3390/vetsci6020052