
Felis Catus Gammaherpesvirus 1 DNAemia in Whole Blood from Therapeutically Immunosuppressed or Retrovirus-Infected Cats

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
- Patient Group 1 (P1, n = 18) comprised blood samples from cats undergoing therapeutic immunosuppression with cyclosporine at a minimum dose of 2 mg/kg per os. The following inclusion criteria were applied:
- 1.
- treatment with oral cyclosporine (Atopica®, Novartis Animal Health Inc., Australia or Neoral®, Novartis Pharmaceuticals Pty Ltd., Macquarie Park, NSW, Australia) for ≥7 days AND
- 2.
- clinical remission from the therapeutic indication for cyclosporine AND
- 3.
- negative FIV antibody and FeLV p27 antigen results
- Patient Group 2 (P2, n = 57) comprised blood samples from cats with progressive FeLV infection and negative FIV serology. Progressive FeLV infection was defined as a positive result on a FeLV p27 antigen PoC test and a positive result on an FeLV proviral DNA qPCR [20] on one (n = 39) or > one (n = 18) occasion.
- Patient Group 3 (P3, n = 5) comprised blood samples from cats co-infected with FIV and FeLV. Inclusion criteria were the same as for P2, except that these cats also had a positive FIV serology result using three PoC tests (SNAP® FIV/FeLV Combo test, IDEXX; Anigen Rapid FIV/FeLV Test Kit, Bionote; WITNESS® FeLV-FIV Test Kit, Zoetis) and had never been vaccinated against FIV.
3. Results
3.1. Frequency of FcaGHV1 DNAemia and Relative Risk in Patients Versus Controls
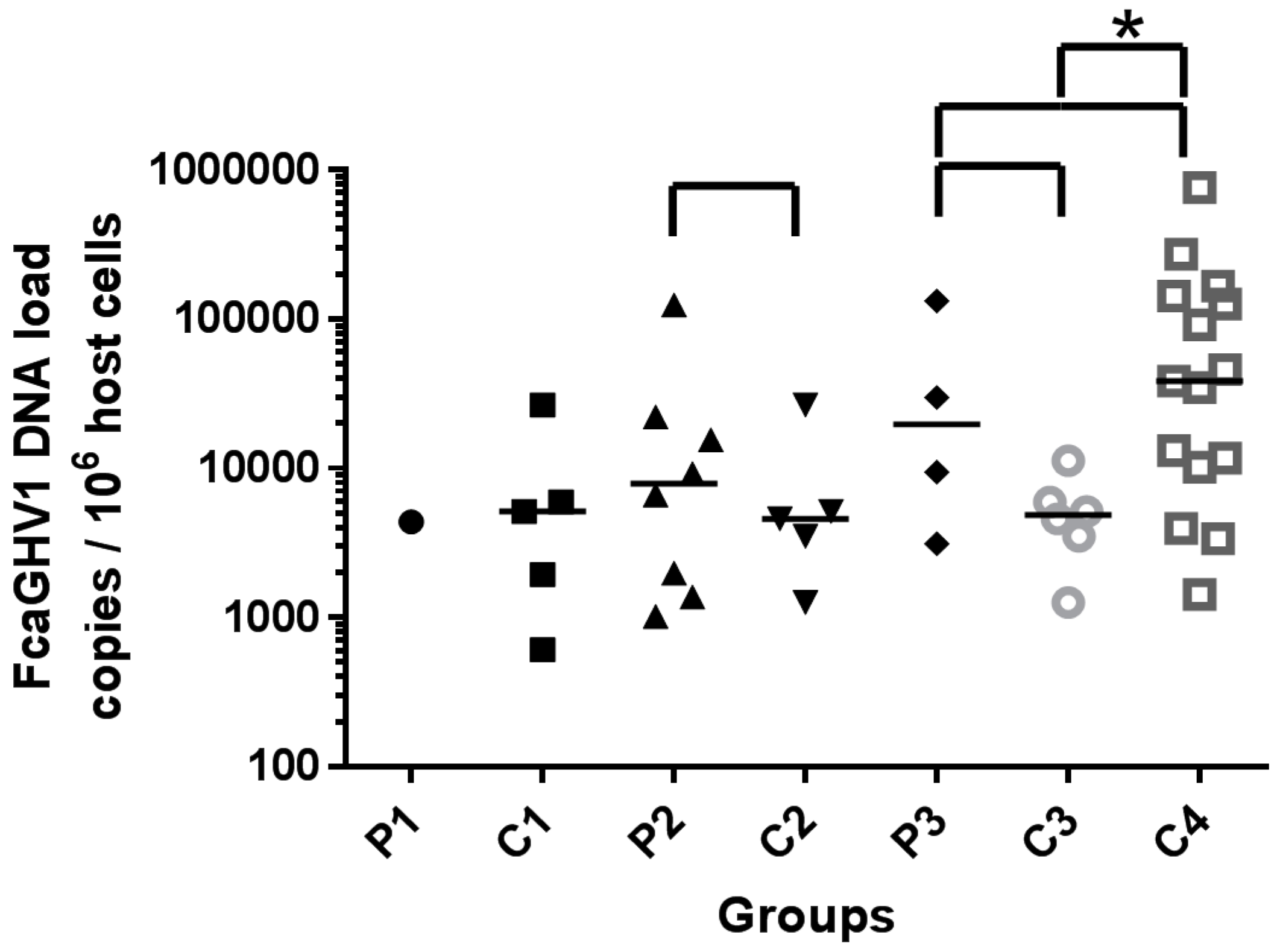
3.2. Whole Blood FcaGHV1 DNA Load
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Troyer, R.M.; Beatty, J.A.; Stutzman-Rodriguez, K.R.; Carver, S.; Lozano, C.C.; Lee, J.S.; Lappin, M.R.; Riley, S.P.D.; Serieys, L.E.K.; Logan, K.A.; et al. Novel gammaherpesviruses in north american domestic cats, bobcats, and pumas: Identification, prevalence, and risk factors. J. Virol. 2014, 88, 3914–3924. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M. Pathogenesis of gammaherpesvirus infections. Vet. Microbiol. 2006, 113, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Kyaw-Tanner, M.; Lee, C.; Robinson, W.F. Characterisation of lymphosarcomas in australian cats using polymerase chain reaction and immunohistochemical examination. Aust. Vet. J. 2001, 79, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Beatty, J. Viral causes of feline lymphoma: Retroviruses and beyond. Vet. J. 2014, 201, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Carbone, A.; Cesarman, E.; Spina, M.; Gloghini, A.; Schulz, T.F. Hiv-associated lymphomas and gamma-herpesviruses. Blood 2009, 113, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Heslop, H.E. Biology and treatment of epstein-barr virus–associated non-hodgkin lymphomas. Hematol. Am. Soc. Hematol. Educ. Program 2005, 2005, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Beatty, J.A.; Troyer, R.M.; Carver, S.; Barrs, V.R.; Espinasse, F.; Conradi, O.; Stutzman-Rodriguez, K.; Chan, C.C.; Tasker, S.; Lappin, M.R.; et al. Felis catus gammaherpesvirus 1; a widely endemic potential pathogen of domestic cats. Virology 2014, 460, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Ertl, R.; Korb, M.; Langbein-Detsch, I.; Klein, D. Prevalence and risk factors of gammaherpesvirus infection in domestic cats in central europe. Virol. J. 2015, 12, 1–4. [Google Scholar] [CrossRef] [PubMed]
- McLuckie, A.; Tasker, S.; Dhand, N.K.; Spencer, S.; Beatty, J.A. High prevalence of felis catus gammaherpesvirus 1 infection in haemoplasma-infected cats supports co-transmission. Vet. J. 2016, 214, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Stutzman-Rodriguez, K.; Rovnak, J.; VandeWoude, S.; Troyer, R.M. Domestic cats seropositive for felis catus gammaherpesvirus 1 are often qpcr negative. Virology 2016, 498, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Ljungman, P.; Griffiths, P.; Paya, C. Definitions of cytomegalovirus infection and disease in transplant recipients. Clin. Infect. Dis. 2002, 34, 1094–1097. [Google Scholar] [CrossRef] [PubMed]
- Piriou, E.R.; van Dort, K.; Nanlohy, N.M.; Miedema, F.; van Oers, M.H.; van Baarle, D. Altered ebv viral load setpoint after hiv seroconversion is in accordance with lack of predictive value of ebv load for the occurrence of aids-related non-hodgkin lymphoma. J. Immunol. 2004, 172, 6931–6937. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.X.; Kim, S.C.; Chima, C.O.; Israel, B.F.; Lawless, K.M.; Eagan, P.A.; Elmore, S.; Moore, D.T.; Schichman, S.A.; Swinnen, L.J.; et al. Epstein-barr viral load as a marker of lymphoma in aids patients. J. Med. Virol. 2005, 75, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Stevens, S.J.C.; Blank, B.S.N.; Smits, P.H.M.; Meenhorst, P.L.; Middeldorp, J.M. High epstein-barr virus (ebv) DNA loads in hiv-infected patients: Correlation with antiretroviral therapy and quantitative ebv serology. Off. J. Int. AIDS Soc. 2002, 16, 993–1001. [Google Scholar] [CrossRef]
- Dehee, A.; Asselot, C.; Piolot, T.; Jacomet, C.; Rozenbaum, W.; Vidaud, M.; Garbarg-Chenon, A.; Nicolas, J.C. Quantification of epstein-barr virus load in peripheral blood of human immunodeficiency virus-infected patients using real-time pcr. J. Med. Virol. 2001, 65, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Roberts, E.S.; VanLare, K.A.; Strehlau, G.; Peyrou, M.; Roycroft, L.M.; King, S. Safety, tolerability, and pharmacokinetics of 6-month daily dosing of an oral formulation of cyclosporine (atopica for cats®) in cats. J. Vet. Pharmacol. Ther. 2014, 37, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Gregory, C.R. Immunosuppresive agents. In Kirk’s Current Veterinary Therapy: Xiv Small Animal Practice; Bonagura, J.D., Ed.; Saunders Elsevier: St. Louis, MO, USA, 2009; Volume XIV, pp. 254–259. [Google Scholar]
- Hoover, E.A.; Mullins, J.I. Feline leukemia virus infection and diseases. J. Am. Vet. Med. Assoc. 1991, 199, 1287–1297. [Google Scholar] [PubMed]
- Pedersen, N.C.; Torten, M.; Rideout, B.; Sparger, E.; Tonachini, T.; Luciw, P.A.; Ackley, C.; Levy, N.; Yamamoto, J. Feline leukemia virus infection as a potentiating cofactor for the primary and secondary stages of experimentally induced feline immunodeficiency virus infection. J. Virol. 1990, 64, 598–606. [Google Scholar] [PubMed]
- Westman, M.E.; Malik, R.; Hall, E.; Sheehy, P.A.; Norris, J.M. Comparison of three feline leukaemia virus (felv) point-of-care antigen test kits using blood and saliva. Comp. Immunol. Microbiol. Infect. Dis. 2017, 50, 88–96. [Google Scholar] [CrossRef] [PubMed]
- IRIS. Iris Staging of CKD (Modified 2015). Available online: http://www.iris-kidney.com/pdf/staging-of-ckd.pdf (accessed on 10 October 2016).
- McLuckie, A.J.; Barrs, V.R.; Smith, A.L.; Beatty, J.A. Detection of felis catus gammaherpesvirus 1 (fcaghv1) in peripheral blood b- and t-lymphocytes in asymptomatic, naturally-infected domestic cats. Virology 2016, 497, 211–216. [Google Scholar] [CrossRef] [PubMed]
- TerWee, J.A.; Carlson, J.K.; Sprague, W.S.; Sondgeroth, K.S.; Shropshire, S.B.; Troyer, J.L.; VandeWoude, S. Prevention of immunodeficiency virus induced cd4+ t-cell depletion by prior infection with a non-pathogenic virus. Virology 2008, 377, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Lowe, A.D.; Campbell, K.L.; Graves, T. Glucocorticoids in the cat. Vet. Dermatol. 2008, 19, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Schmiedt, C.W.; Grimes, J.A.; Holzman, G.; McAnulty, J.F. Incidence and risk factors for development of malignant neoplasia after feline renal transplantation and cyclosporine-based immunosuppression. Vet. Comp. Oncol. 2009, 7, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Ngo, S.T.; Steyn, F.J.; McCombe, P.A. Gender differences in autoimmune disease. Front. Neuroendocrinol. 2014, 35, 347–369. [Google Scholar] [CrossRef] [PubMed]
- Pollard, K.M. Gender differences in autoimmunity associated with exposure to environmental factors. J. Autoimmun. 2012, 38, J177–J186. [Google Scholar] [CrossRef] [PubMed]
- Gershwin, L.J. Autoimmune diseases in small animals. Vet. Clin. N. Am. 2010, 40, 439–457. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.C. A review of immunologic diseases of the dog. Vet. Immunol. Immunopathol. 1999, 69, 251–342. [Google Scholar] [CrossRef]
- Oohashi, E.; Yamada, K.; Oohashi, M.; Ueda, J. Chronic progressive polyarthritis in a female cat. J. Vet. Med. Sci./Jpn. Soc. Vet. Sci. 2010, 72, 511–514. [Google Scholar] [CrossRef]
- Kohn, B.; Weingart, C.; Eckmann, V.; Ottenjann, M.; Leibold, W. Primary immune-mediated hemolytic anemia in 19 cats: Diagnosis, therapy, and outcome (1998–2004). J. Vet. Int. Med. 2006, 20, 159–166. [Google Scholar]
- Stevens, S.J.C.; Verschuuren, E.A.M.; Verkuijlen, S.; Van den Brule, A.J.C.; Meijer, C.; Middeldorp, J.M. Role of epstein-barr virus DNA load monitoring in prevention and early detection of post-transplant lymphoproliferative disease. Leuk. Lymphoma 2002, 43, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Durham, A.C.; Mariano, A.D.; Holmes, E.S.; Aronson, L. Characterization of post transplantation lymphoma in feline renal transplant recipients. J. Comp. Pathol. 2014, 150, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Wormser, C.; Mariano, A.; Holmes, E.S.; Aronson, L.R.; Volk, S.W. Post-transplant malignant neoplasia associated with cyclosporine-based immunotherapy: Prevalence, risk factors and survival in feline renal transplant recipients. Vet. Comp. Oncol. 2016, 14, e126–e134. [Google Scholar] [CrossRef] [PubMed]
- Khan, G.; Jawad, M. Global burden of deaths from epstein-barr virus attributable malignancies 1990–2010. Infect. Agents Cancer 2014, 9, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Carver, S.; Beatty, J.A.; Troyer, R.M.; Harris, R.L.; Stutzman-Rodriguez, K.; Barrs, V.R.; Chan, C.C.; Tasker, S.; Lappin, M.R.; VandeWoude, S. Closing the gap on causal processes of infection risk from cross-sectional data: Structural equation models to understand infection and co-infection. Parasit. Vectors 2015, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kozić, S.; Vince, A.; Beš, J.I.; Rode, O.Đ.; Lepej, S.Ž.; Poljak, M.; Bozic, M.; Kessler, H.H. Evaluation of a commercial real-time pcr assay for quantitation of epstein-barr virus DNA in different groups of patients. J. Virol. Methods 2006, 135, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Petrara, M.R.; Freguja, R.; Gianesin, K.; Zanchetta, M.; De Rossi, A. Epstein-barr virus-driven lymphomagenesis in the context of human immunodeficiency virus type 1 infection. Front. Microbiol. 2013, 4, 311. [Google Scholar] [CrossRef] [PubMed]
- Bienzle, D. Fiv in cats—A useful model of hiv in people? Vet. Immunol. Immunopathol. 2014, 159, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Elder, J.H.; Lin, Y.-C.; Fink, E.; Grant, C.K. Feline immunodeficiency virus (fiv) as a model for study of lentivirus infections: Parallels with hiv. Curr. HIV Res. 2010, 8, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.I. Epstein-barr virus vaccines. Clin. Transl. Immunol. 2015, 4, 6. [Google Scholar] [CrossRef] [PubMed]
 bracket. Significant difference highlighted by *.
bracket. Significant difference highlighted by *.
bracket. Significant difference highlighted by *.
bracket. Significant difference highlighted by *.

{kind=link}
| Group | (n =) | Description | Age (Years) | Sex (n =) | |||
|---|---|---|---|---|---|---|---|
| Median | Range | IQR a | M b | F c | |||
| P1 | 18 | therapeutic immunosuppression | 5.7 | 0.7–18.5 | 3.0–12.3 | 4 | 14 |
| C1 | 58 | controls matched to P1 | 6.5 | 0.7–20.2 | 3.7–10.8 | 13 | 45 |
| P2 | 57 | FeLV-infected | 3.9 | 0.4–15.3 | 2.0–8.0 | 35 | 22 |
| C2 | 57 | controls matched to P2 | 4.7 | 0.5–16.0 | 2.7–7.9 | 35 | 22 |
| P3 | 5 | FIV/FeLV-co-infected | 9 | 0.8–12.5 | 2.9–11.3 | 5 | 0 |
| C3 | 39 | controls matched to P3 | 6 | 1.1–16.0 | 3.4–9.3 | 39 | 0 |
| C4 | 21 | historical FIV-infected | 12 | 2.5–17.1 | 8.5–13.7 | 21 | 0 |
| Group | FcaGHV1-Infected | FcaGHV1 Negative | Prevalence (%) | Contingency Table Results | |||
|---|---|---|---|---|---|---|---|
| Groups Compared | Fisher’s Exact Test a | RR b | 95% CI c | ||||
| P1 | 1 | 17 | 5.6 | P1 vs. C1 | 1.000 | 0.5556 | 0.07145 to 4.320 |
| C1 | 5 | 53 | 8.6 | ||||
| P2 | 8 | 49 | 14.0 | P2 vs. C2 | 0.5572 | 1.268 | 0.7892 to 2.039 |
| C2 | 5 | 52 | 8.8 | ||||
| P3 | 4 | 1 | 80.0 | P3 vs. C3 | 0.0068 | 5.200 | 2.207 to 12.25 |
| C3 | 6 | 33 | 15.4 | C4 vs. C3 | <0.0001 | 4.643 | 2.119 to 10.17 |
| C4 | 15 | 6 | 71.4 | P3 vs. C4 | 1.000 | 1.120 | 0.6691 to 1.875 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McLuckie, A.J.; Barrs, V.R.; Wilson, B.; Westman, M.E.; Beatty, J.A. Felis Catus Gammaherpesvirus 1 DNAemia in Whole Blood from Therapeutically Immunosuppressed or Retrovirus-Infected Cats. Vet. Sci. 2017, 4, 16. https://doi.org/10.3390/vetsci4010016
McLuckie AJ, Barrs VR, Wilson B, Westman ME, Beatty JA. Felis Catus Gammaherpesvirus 1 DNAemia in Whole Blood from Therapeutically Immunosuppressed or Retrovirus-Infected Cats. Veterinary Sciences. 2017; 4(1):16. https://doi.org/10.3390/vetsci4010016
Chicago/Turabian StyleMcLuckie, Alicia J., Vanessa R. Barrs, Bethany Wilson, Mark E. Westman, and Julia A. Beatty. 2017. "Felis Catus Gammaherpesvirus 1 DNAemia in Whole Blood from Therapeutically Immunosuppressed or Retrovirus-Infected Cats" Veterinary Sciences 4, no. 1: 16. https://doi.org/10.3390/vetsci4010016
APA StyleMcLuckie, A. J., Barrs, V. R., Wilson, B., Westman, M. E., & Beatty, J. A. (2017). Felis Catus Gammaherpesvirus 1 DNAemia in Whole Blood from Therapeutically Immunosuppressed or Retrovirus-Infected Cats. Veterinary Sciences, 4(1), 16. https://doi.org/10.3390/vetsci4010016