
Bovine Papillomavirus Genotypic Diversity and a Putative Novel Viral Type in Ecuador

 , , , , , and
, , , , , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
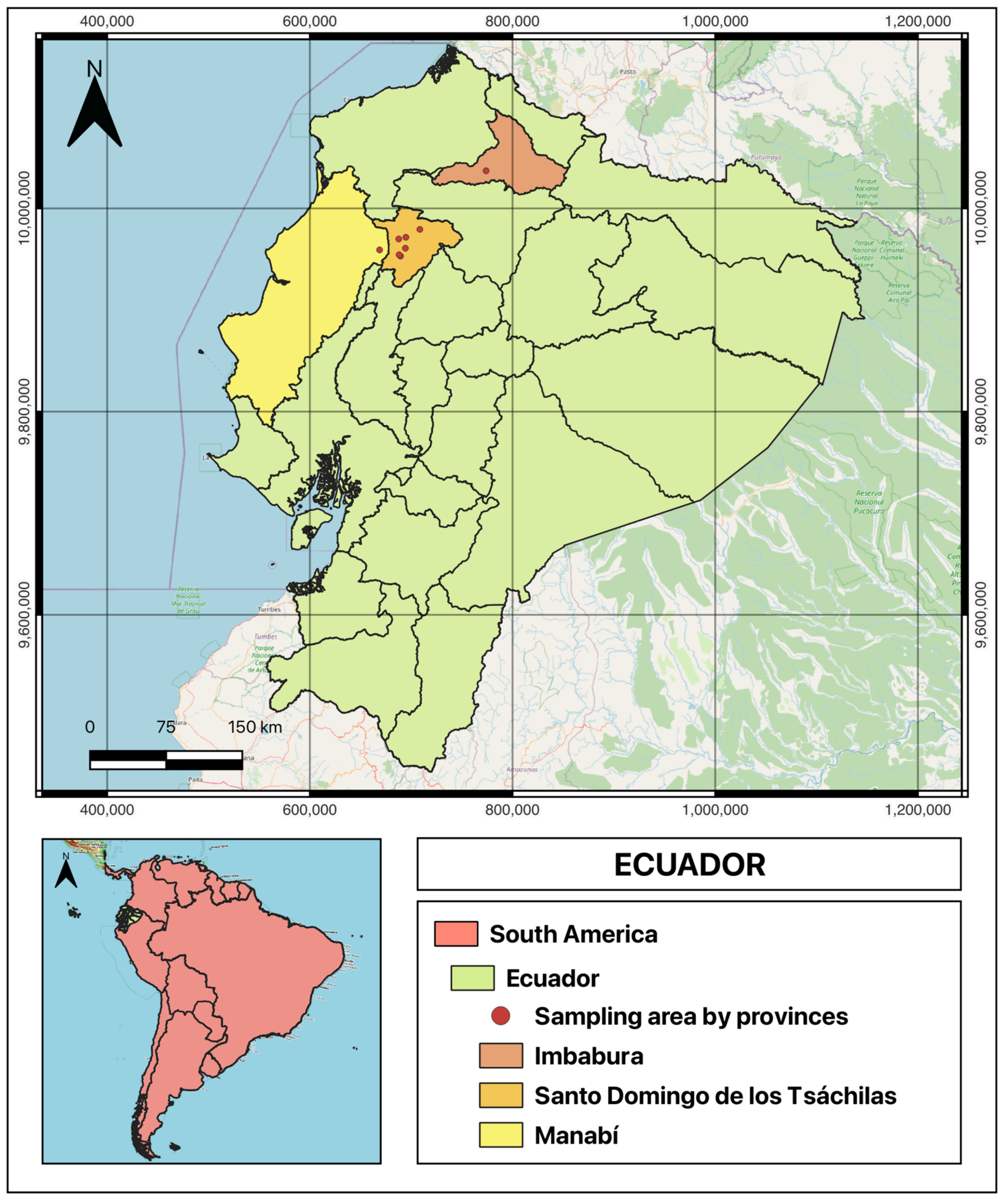
2.1. Sampling
2.2. DNA Extraction
2.3. Identification of Bovine Papillomavirus: Amplification and Sequencing
2.4. Phylogenetic and Data Analysis
2.5. Ethics Approval and Consent to Participate
3. Results
3.1. Samples
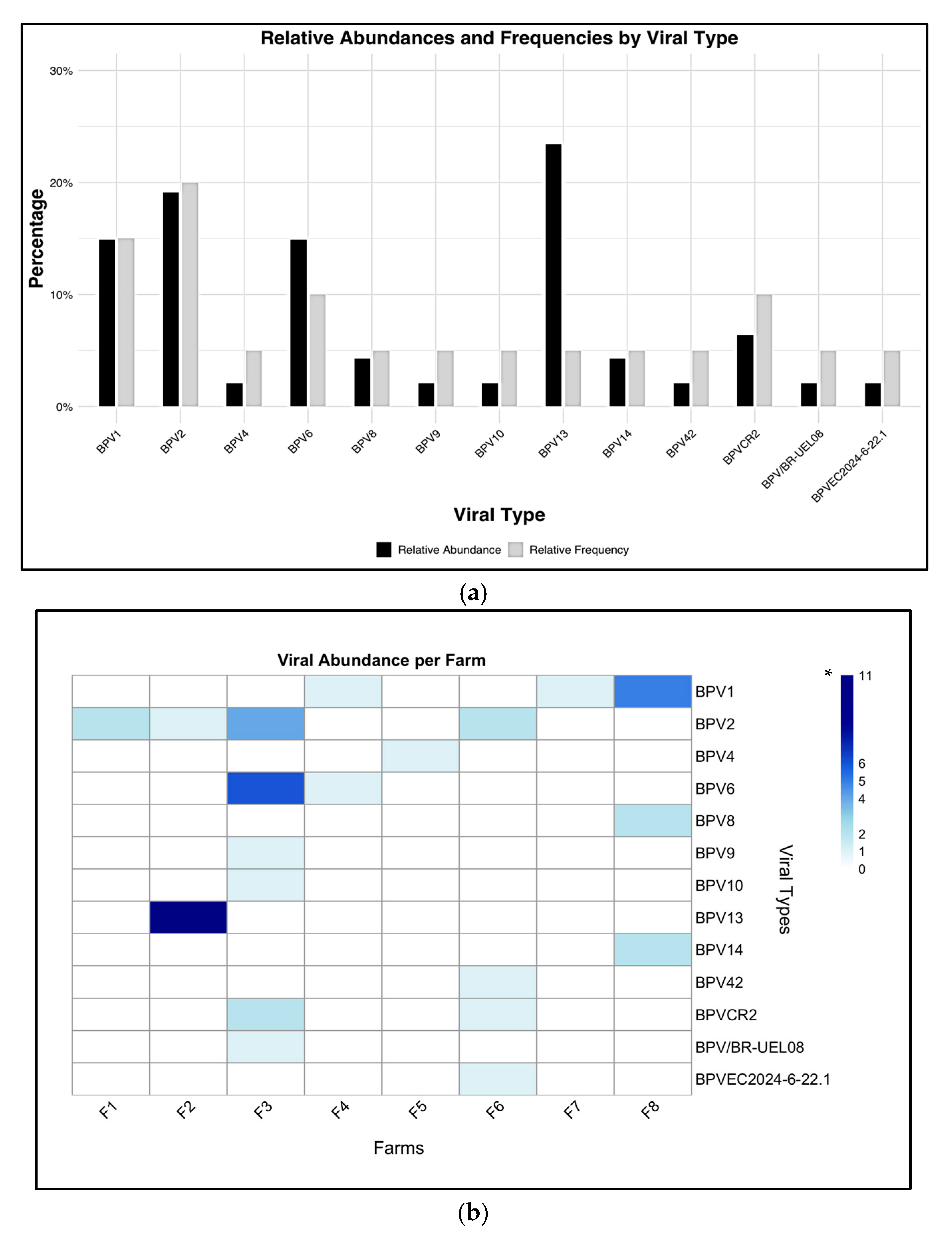
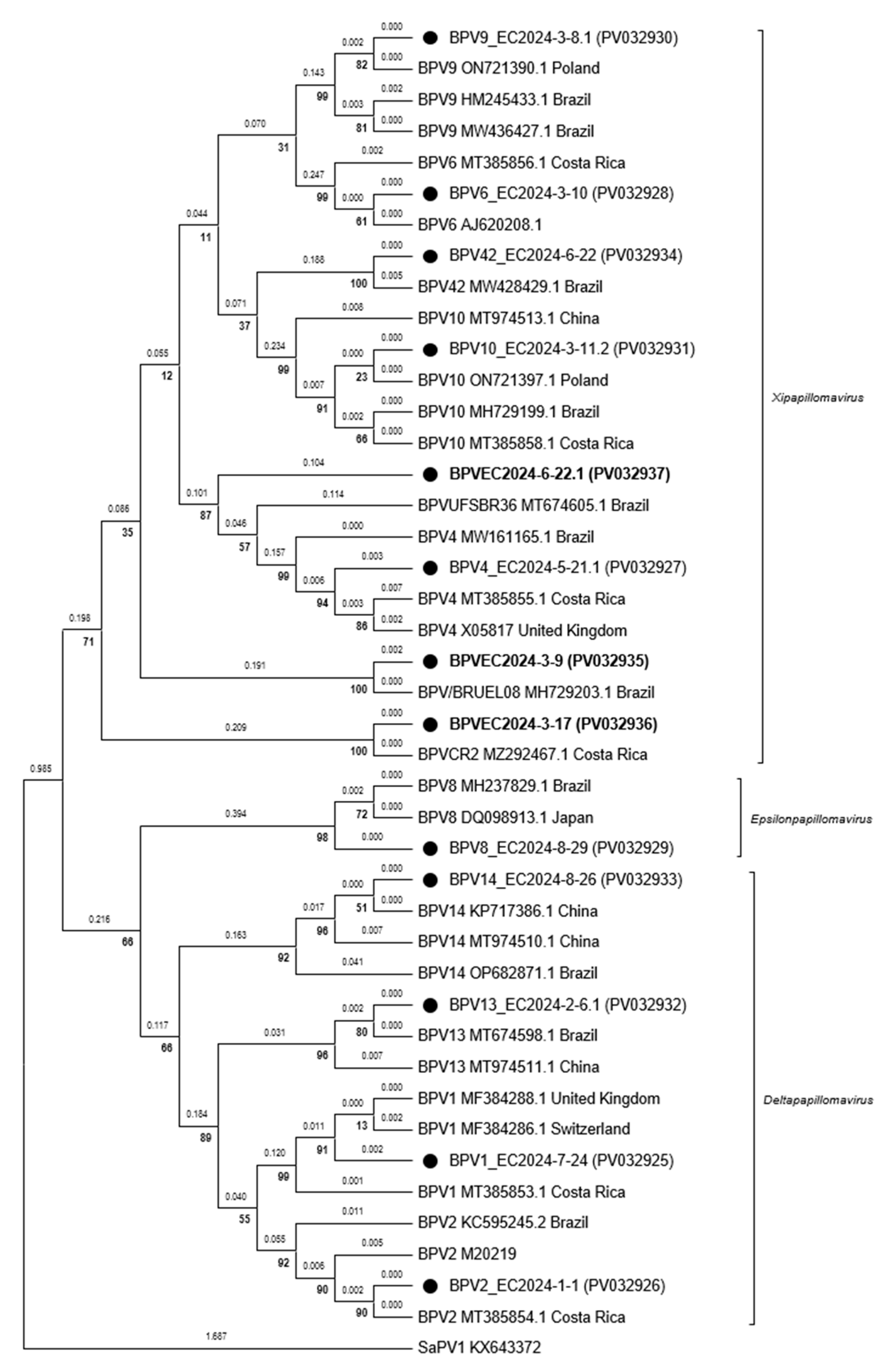
3.2. Amplification, Sequencing and Phylogeny
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Araldi, R.P.; Assaf, S.M.R.; Carvalho, R.F.d.; Carvalho, M.A.C.R.d.; Souza, J.M.d.; Magnelli, R.F.; Módolo, D.G.; Roperto, F.P.; Stocco, R.d.C.; Beçak, W. Papillomaviruses: A systematic review. Genet. Mol. Biol. 2017, 40, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Daudt, C.; Da Silva, F.R.C.; Lunardi, M.; Alves, C.B.D.T.; Weber, M.N.; Cibulski, S.P.; Alfieri, A.F.; Alfieri, A.A.; Canal, C.W. Papillomaviruses in ruminants: An update. Transbound. Emerg. Dis. 2018, 65, 1381–1395. [Google Scholar] [CrossRef] [PubMed]
- Lunardi, M.; Alcindo, A.; Alejandro, R.; Fernandes, A. Bovine Papillomaviruses—Taxonomy and Genetic Features. In Current Issues in Molecular Virology—Viral Genetics and Biotechnological Applications; InTech: Rijeka, Croatia, 2013. [Google Scholar]
- Ugochukwu, I.C.I.; Aneke, C.I.; Idoko, I.S.; Sani, N.A.; Amoche, A.J.; Mshiela, W.P.; Ede, R.E.; Ibrahim, N.D.G.; Njoku, C.I.O.; Sackey, A.K.B. Bovine papilloma: Aetiology, pathology, immunology, disease status, diagnosis, control, prevention and treatment: A review. Comp. Clin. Pathol. 2019, 28, 737–745. [Google Scholar] [CrossRef]
- Van Doorslaer, K. Evolution of the Papillomaviridae. Virology 2013, 445, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Finlay, M.; Yuan, Z.; Burden, F.; Trawford, A.; Morgan, I.M.; Saveria Campo, M.; Nasir, L. The detection of Bovine Papillomavirus type 1 DNA in flies. Virus Res. 2009, 144, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, V.; Cardoso, M.; Carneiro, A.; Silva, R.; Teixeira, M.; Wanderley, A.; Wanderley, E.; Franco, E. Descrição clínica e histopatológica da papilomatose cutânea bovina (BPV). Ciência Anim. Bras. 2008, 9, 1079–1088. [Google Scholar]
- Radostits, O.M.; Gay, C.C.; Hinchcliff, K.W.; Constable, P.D. Veterinary Medicine: A Textbook of the Diseases of Cattle, Sheep, Goats, Pigs and Horses, 10th ed.; Elsevier: Amsterdam, The Netherlands, 2006; pp. 1421–1423. [Google Scholar]
- Campo, M.S. Bovine papillomavirus: Old system, new lessons? In Papillomavirus Research: From Natural History to Vaccine and Beyond; Caister Academic Press: Wymondham, UK, 2006; pp. 1–33. [Google Scholar]
- PaVe: The Papillomavirus Episteme. Available online: https://pave.niaid.nih.gov/ (accessed on 1 July 2023).
- Roperto, S.; Russo, V.; Leonardi, L.; Martano, M.; Corrado, F.; Riccardi, M.G.; Roperto, F. Bovine Papillomavirus Type 13 Expression in the Urothelial Bladder Tumours of Cattle. Transbound. Emerg. Dis. 2016, 63, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Sauthier, J.T.; Daudt, C.; da Silva, F.R.C.; Alves, C.D.B.T.; Mayer, F.Q.; Bianchi, R.M.; Driemeier, D.; Streit, R.S.A.; Staats, C.C.; Canal, C.W.; et al. The genetic diversity of “papillomavirome” in bovine teat papilloma lesions. Anim. Microbiome 2021, 3, 51. [Google Scholar] [CrossRef] [PubMed]
- Löhr, C.V.; Juan-Sallés, C.; Rosas-Rosas, A.; García, A.P.; Garner, M.M.; Teifke, J.P. sarcoids in captive zebras (equus burchellii): Association with bovine papillomavirus type 1 infection. J. Zoo Wildl. Med. 2005, 36, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Figueirêdo, R.P.; Santos, G.F.; Oliveira, L.B.; Santos, L.A.B.O.; Barreto, D.M.; Cândido, A.L.; Campos, A.C.; Azevedo, E.O.; Batista, M.V.A. High Genotypic Diversity, Putative New Types and Intra-Genotype Variants of Bovine Papillomavirus in Northeast Brazil. Pathogens 2020, 9, 748. [Google Scholar] [CrossRef] [PubMed]
- Alfaro-Mora, R.; Castro-Vásquez, R.; Zobba, R.; Antuofermo, E.; Burrai, G.P.; Alberti, A.; Dolz, G. Molecular typing of bovine papillomaviruses in Costa Rica. Vet. Res. Commun. 2023, 47, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Lennin, S.; Andrade, O. Detección Y genotipificación molecular del virus de papiloma bovino en lesiones de animales afectados por papilomatosis cutánea. Centrosur Agrar. 2020, 1, 34–45. [Google Scholar]
- Vásquez Bonilla, M.M.; Guerrero-Freire, M.S.; Ledesma, Y.; Laglaguano, J.C.; de Waard, J.H. A rapid and inexpensive 96-well DNA-extraction method from blood using silicon dioxide powder (Glassmilk). Biol. Methods Protoc. 2024, 9, bpae079. [Google Scholar] [CrossRef] [PubMed]
- Forslund, O.; Antonsson, A.; Nordin, P.; Stenquist, B.; Göran Hansson, B. A broad range of human papillomavirus types detected with a general PCR method suitable for analysis of cutaneous tumours and normal skin. J. Gen. Virol. 1999, 80, 2437–2443. [Google Scholar] [CrossRef] [PubMed]
- Crespo, S.E.I.; Lunardi, M.; Otonel, R.A.A.; Headley, S.A.; Alfieri, A.F.; Alfieri, A.A. Genetic characterization of a putative new type of bovine papillomavirus in the Xipapillomavirus 1 species in a Brazilian dairy herd. Virus Genes 2019, 55, 682–687. [Google Scholar] [CrossRef] [PubMed]
- Pyrek, P.; Bednarski, M.; Popiel, J.; Siedlecka, M.; Karwańska, M. Genetic Evaluation of Bovine Papillomavirus Types Associated with Teat Papillomatosis in Polish Dairy Cattle with the Report of a New Putative Type. Pathogens 2023, 12, 1278. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.C.R.; Batista, M.V.A.; Silva, M.A.R.; Balbino, V.Q.; Freitas, A.C. Detection of Bovine Papillomavirus Types, Co-Infection and a Putative New BPV11 Subtype in Cattle. Transbound. Emerg. Dis. 2012, 59, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-García, A.; Ponce-de-León-Rosales, S.; Cantú-de-León, D.; Fragoso-Ontiveros, V.; Martínez-Ramírez, I.; Orozco-Colín, A.; Mohar, A.; Lizano, M. Impact of human papillomavirus coinfections on the risk of high-grade squamous intraepithelial lesion and cervical cancer. Gynecol. Oncol. 2014, 134, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, A.K.; Katki, H.A.; Hildesheim, A.; Rodríguez, A.C.; Quint, W.; Schiffman, M.; Van Doorn, L.-J.; Porras, C.; Wacholder, S.; Gonzalez, P.; et al. Human Papillomavirus Infection with Multiple Types: Pattern of Coinfection and Risk of Cervical Disease. J. Infect. Dis. 2011, 203, 910–920. [Google Scholar] [CrossRef] [PubMed]
- Senapati, R.; Nayak, B.; Kar, S.K.; Dwibedi, B. HPV genotypes co-infections associated with cervical carcinoma: Special focus on phylogenetically related and non-vaccine targeted genotypes. PLoS ONE 2017, 12, e0187844. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | GenBank Accession Number | Genus | Reference Viral Type | |||
|---|---|---|---|---|---|---|
| NCBI | Id (%) | PaVe | Id (%) | |||
| BPV1_EC2024-7-24 sample F724_241123 | PV032925 | Delta | BPV1 (MT385853.1) | 100 | BPV1 (X02346) | 99.3 |
| BPV2_EC2024-1-1 sample F101_250823 | PV032926 | Delta | BPV2 (LC426022.1) | 100 | BPV2 (M20219) | 99.3 |
| BPV4_EC2024-5-21.1 sample F521.1_211123 | PV032927 | Xi | BPV4 (OP682875.1) | 99.29 | BPV4 (X05817) | 99.3 |
| BPV6_EC2024-3-10 sample F310_061023 | PV032928 | Xi | BPV6 (MH729201.1) | 100 | BPV6 (AB331651) | 100 |
| BPV8_EC2024-8-29 sample F829_091223 | PV032929 | Epsilon | BPV8 (MH237829.1) | 99.75 | BPV8 (DQ098913) | 99.8 |
| BPV9_EC2024-3-8.1 sample F308.1_061023 | PV032930 | Xi | BPV9 (ON721390.1) | 100 | BPV9 (AB331650) | 99.1 |
| BPV10_EC2024-3-11.2 sample F311.2_061023 | PV032931 | Xi | BPV10 (ON721397.1) | 100 | BPV10 (AB331651) | 99.8 |
| BPV13_EC2024-2-6.1 sample F206.1_310823 | PV032932 | Delta | BPV13 (MG818475.1) | 100 | BPV13 (JQ798171) | 100 |
| BPV14_EC2024-8-26 sample F826_261123 | PV032933 | Delta | BPV14 (KR868228.1) | 99.77 | BPV14 (KP276343) | 99.8 |
| BPV42_EC2024-6-22 sample F622_241123 | PV032934 | Xi | BPV42 (MW428429.1) | 99.54 | BPV43 (MW428429.1) | 99.5 |
| BPVEC2024-3-9 sample F309_061023 | PV032935 | Xi | BPV-BR-UEL08 (MH729203.1) | 99.77 | BPV40 (MW428425.1) | 78.27 |
| BPVEC2024-3-17 sample F317_061023 | PV032936 | Xi | BPV-CR2 (MZ292467.1) | 100 | BVP12 (JF834523) | 86.4 |
| BPVEC2024-6-22.1 sample F622.1_241123 | PV032937 | Xi | BPV-UFSBR36 (MT674605.1) | 81.71 | BPV4 (X05817.1) | 77.88 |
| Farm | Bovine | Sample’s Code | PaVe | Body’s Part |
|---|---|---|---|---|
| 2 | 04 | F204_310823 | BPV2 | Neck |
| F204.1_310823 | BPV13 | Neck | ||
| 3 | 08 | F308_061023 | * | Ear |
| F308.1_061023 | BPV9 | Mouth | ||
| F308.2_061023 | BPV2 | Navel | ||
| 3 | 09 | F309_061023 | BPV-BR-UEL08 | Ear |
| F309.1_061023 | BPV_CR2 | Udder | ||
| 3 | 11 | F311_061023 | ctioBPV2 | Chest |
| F311.1_061023 | BPV6 | Ear | ||
| F311.2_061023 | BPV10 | Udder | ||
| 3 | 12 | F312_061023 | BPV2 | Udder |
| F312.1_061023 | * | Udder | ||
| 6 | 22 | F622_241123 | BPV42 | Nose |
| F622.1_241123 | BPVEC2024-6-22.1 | Neck | ||
| F622.2_241123 | BPV_CR2 | Back | ||
| 8 | 26 | F826_261123 | BPV14 | Back |
| F826.1_261123 | BPV1 | Neck | ||
| F826.2_261123 | BPV2 | Neck |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carvajal-Reina, D.J.; Bedoya-Páez, F.; Guerrero-Freire, M.S.; Ledesma, Y.; Vasco-Julio, D.; de Waard, J.H.; Reyna-Bello, A. Bovine Papillomavirus Genotypic Diversity and a Putative Novel Viral Type in Ecuador. Vet. Sci. 2025, 12, 672. https://doi.org/10.3390/vetsci12070672
Carvajal-Reina DJ, Bedoya-Páez F, Guerrero-Freire MS, Ledesma Y, Vasco-Julio D, de Waard JH, Reyna-Bello A. Bovine Papillomavirus Genotypic Diversity and a Putative Novel Viral Type in Ecuador. Veterinary Sciences. 2025; 12(7):672. https://doi.org/10.3390/vetsci12070672
Chicago/Turabian StyleCarvajal-Reina, Diego J., Fausto Bedoya-Páez, Mónica Salomé Guerrero-Freire, Yanua Ledesma, David Vasco-Julio, Jacobus H. de Waard, and Armando Reyna-Bello. 2025. "Bovine Papillomavirus Genotypic Diversity and a Putative Novel Viral Type in Ecuador" Veterinary Sciences 12, no. 7: 672. https://doi.org/10.3390/vetsci12070672
APA StyleCarvajal-Reina, D. J., Bedoya-Páez, F., Guerrero-Freire, M. S., Ledesma, Y., Vasco-Julio, D., de Waard, J. H., & Reyna-Bello, A. (2025). Bovine Papillomavirus Genotypic Diversity and a Putative Novel Viral Type in Ecuador. Veterinary Sciences, 12(7), 672. https://doi.org/10.3390/vetsci12070672