
Levamisole as a Strategy Against Bacteria from Canine Otitis Externa: An In Vitro Evaluation


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Antimicrobial Susceptibility to Levamisole
2.3. Cytotoxicity Assay Against MRK5
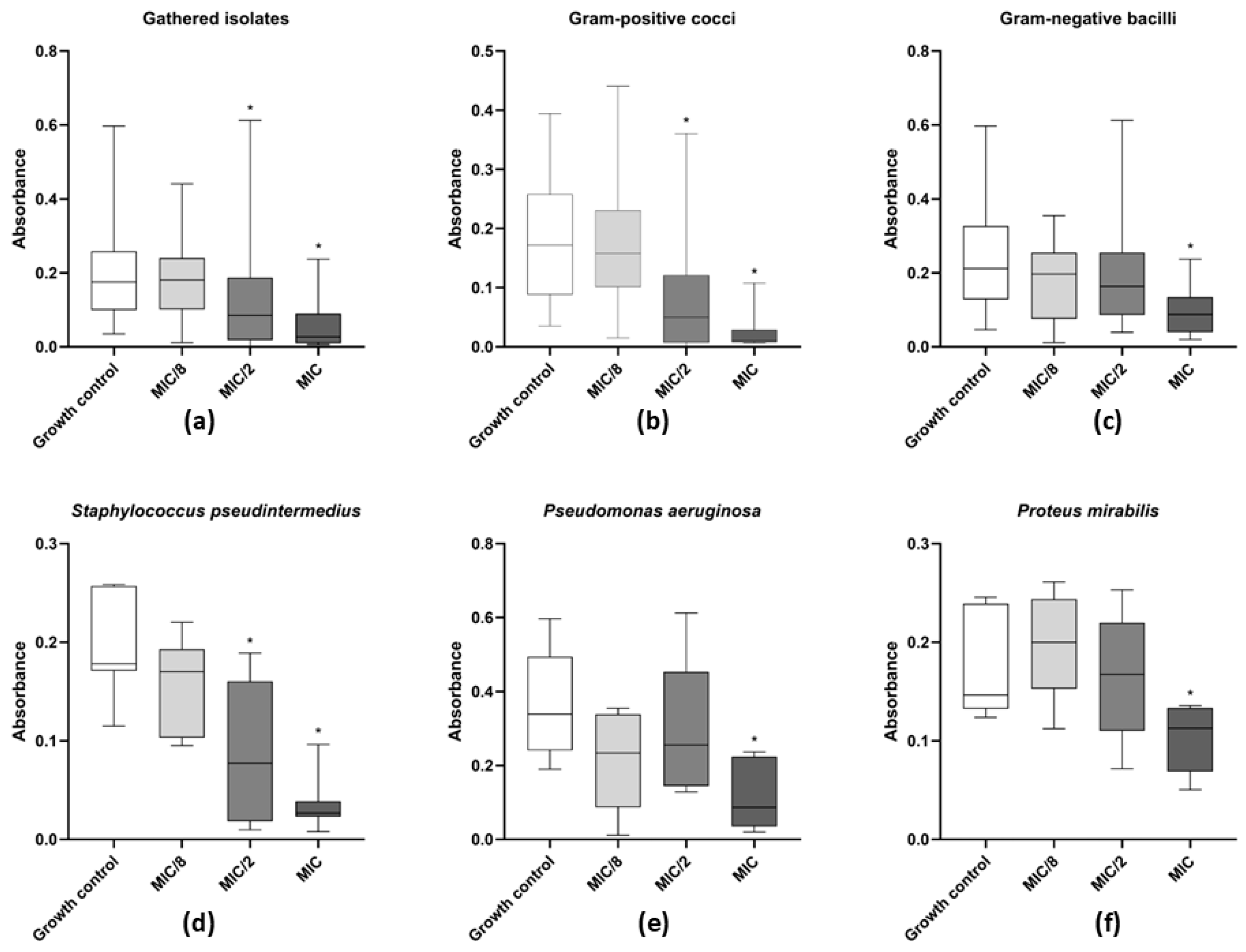
2.4. Susceptibility of Mature Biofilm to Levamisole
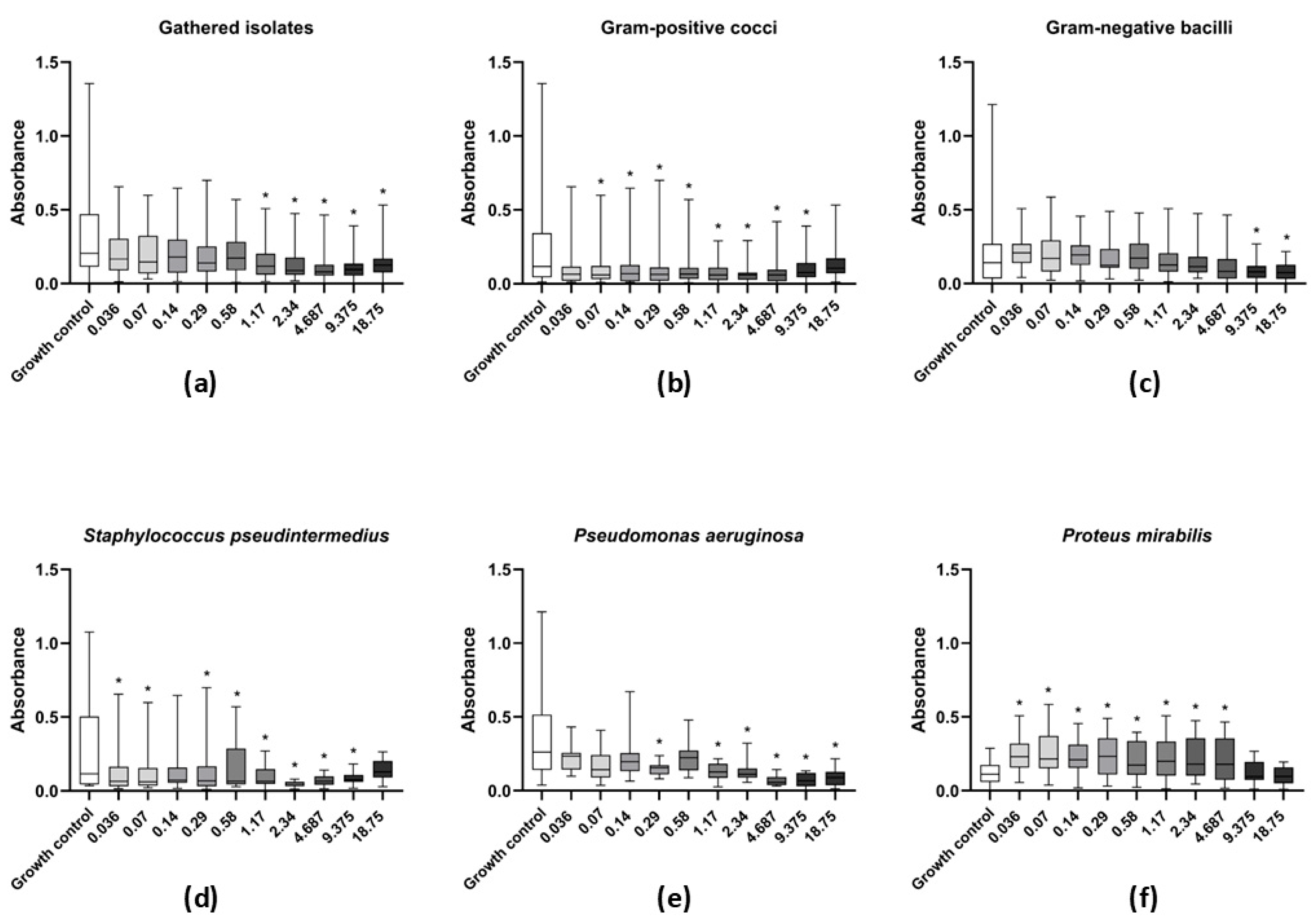
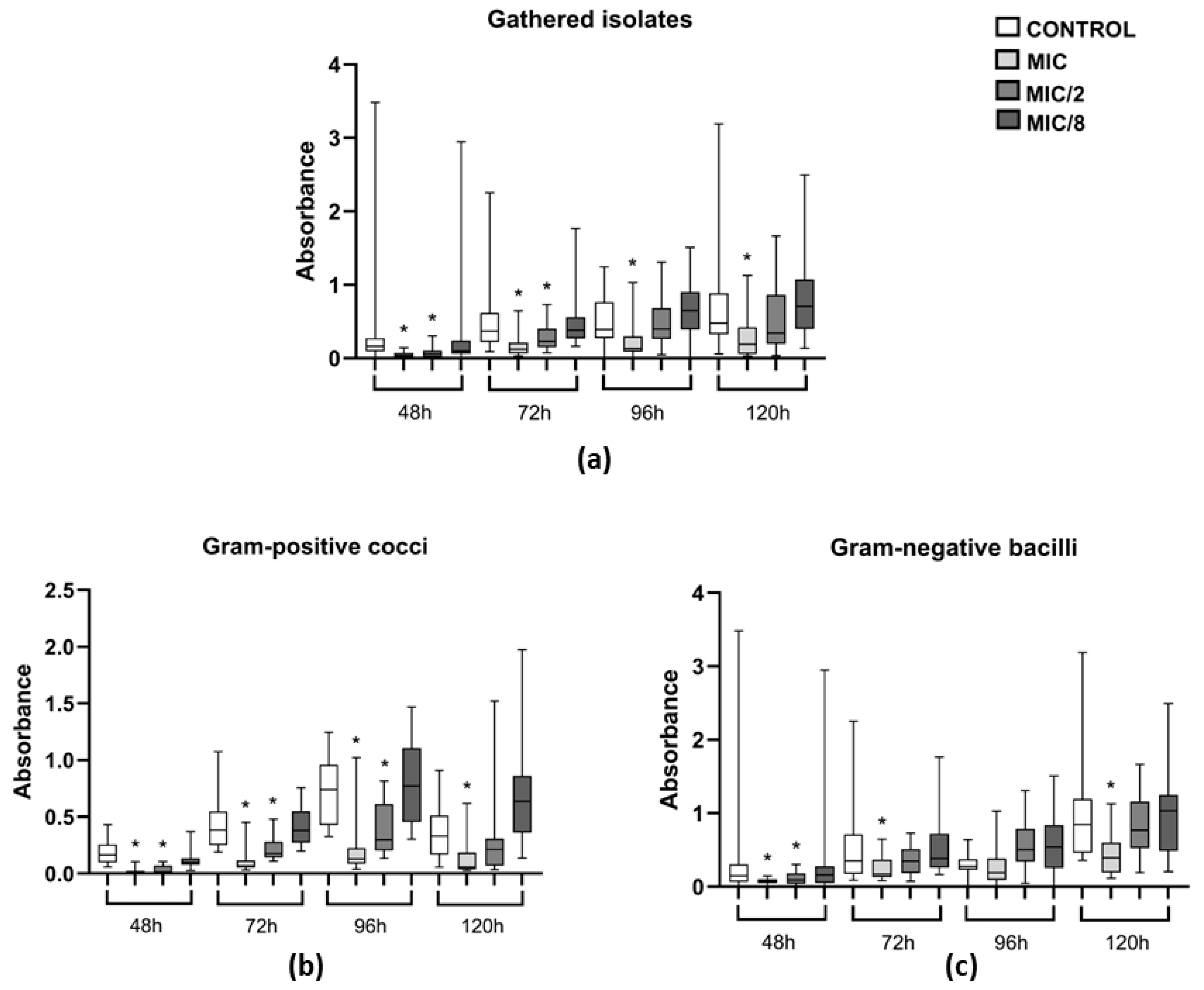
2.5. Activity of Levamisole on Biofilm Formation
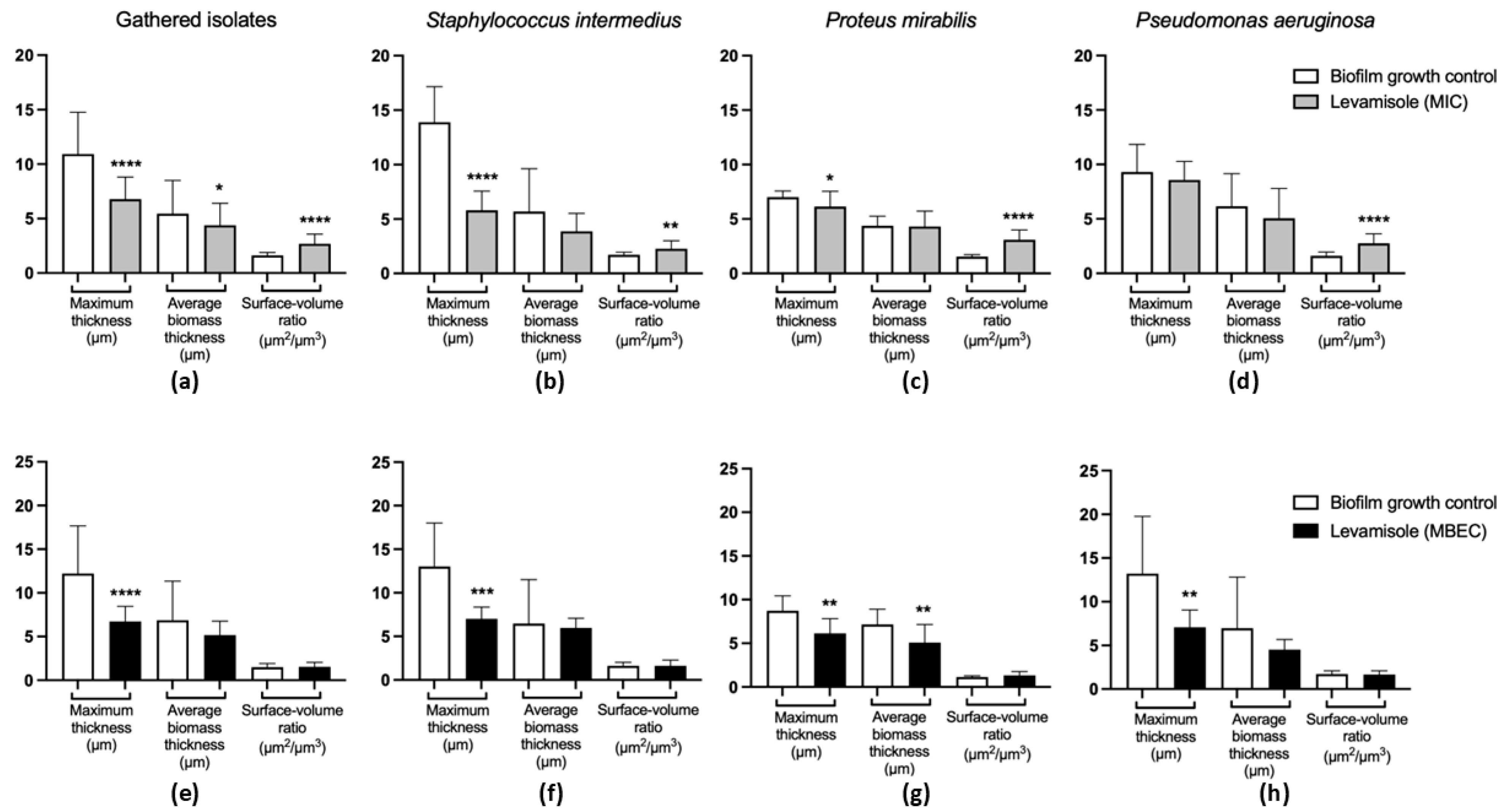
2.6. Assessment of Mature and Growing Biofilms Using Confocal Laser Scanning Microscopy (CLSM)
2.7. Statistical Analysis
3. Results
3.1. Minimum Inhibitory Concentrations (MICs) of Levamisole and Cytotoxicity Assays
3.2. Minimum Biofilm Inhibitory and Eradication Concentrations of Levamisole
3.3. Effect of Levamisole on Bacterial Biofilm Growth
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pye, C. Pseudomonas otitis externa in dogs. Can. Vet. J. 2018, 59, 1231–1234. [Google Scholar] [PubMed] [PubMed Central]
- Heine, P.A. Anatomy of the ear. Vet. Clin. N. Am. Small Anim. Pract. 2004, 34, 379–395. [Google Scholar] [CrossRef] [PubMed]
- Njaa, B.L.; Cole, L.K.; Tabacca, N. Practical otic anatomy and physiology of the dog and cat. Vet. Clin. N. Am. Small Anim. Pract. 2012, 42, 1109–1126. [Google Scholar] [CrossRef]
- Chan, W.Y.; Hickey, E.E.; Khazandi, M.; Page, S.W.; Trott, D.J.; Hill, P.B. In vitro antimicrobial activity of narasin and monensin in combination with adjuvants against pathogens associated with canine otitis externa. Vet. Dermatol. 2020, 31, 138–145. [Google Scholar] [CrossRef] [PubMed]
- May, E.R.; Ratliff, B.E.; Bemis, D.A. Antibacterial effect of N-acetylcysteine in combination with antimicrobials on common canine otitis externa bacterial isolates. Vet. Dermatol. 2019, 30, 531–e161. [Google Scholar] [CrossRef]
- Bajwa, J. Canine otitis externa—Treatment and complications. Can. Vet. J. 2019, 60, 97–99. [Google Scholar] [PubMed Central]
- Seeger, M.G.; Ries, A.S.; Gressler, L.T.; Botton, S.A.; Iglesias, B.A.; Cargnelutti, J.F. In vitro antimicrobial photodynamic therapy using tetra-cationic porphyrins against multidrug-resistant bacteria isolated from canine otitis. Photodiagn. Photodyn. Ther. 2020, 32, 101982. [Google Scholar] [CrossRef]
- Farfán, J.A.P.; Vega, L.G.A.; Espinoza, S.Y.C.; Magallanes, S.G.; Moreno, J.J.S. Methicillin-resistant Staphylococcus schleiferi subspecies coagulans associated with otitis externa and pyoderma in dogs. Open Vet. J. 2021, 11, 364–369. [Google Scholar]
- Kwon, J.; Yang, M.H.; Ko, H.J.; Kim, S.G.; Park, C.; Park, S.C. Antimicrobial resistance and virulence factors of Proteus mirabilis isolated from dog with chronic otitis externa. Pathogens 2022, 11, 1215. [Google Scholar] [CrossRef]
- Shoaib, M.; Aqib, A.I.; Ali, M.M.; Ijaz, M.; Sattar, H.; Ghaffar, A.; Sajid Hasni, M.; Bhutta, Z.A.; Ashfaq, K.; Kulyar, M.F.; et al. Tracking infection and genetic divergence of methicillin-resistant Staphylococcus aureus at pets, pet owners, and environment interface. Front. Vet. Sci. 2022, 9, 900480. [Google Scholar] [CrossRef]
- Naziri, Z.; Majlesi, M. Comparison of the prevalence, antibiotic resistance patterns, and biofilm formation ability of methicillin-resistant Staphylococcus pseudointermedius in healthy dogs and dogs with skin infections. Vet. Res. Commun. 2023, 47, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Hao, R.; Shoaib, M.; Tang, M.; Cao, Z.; Liu, G.; Zhang, Y.; Wang, S.; Shang, R.; Zhang, H.; Pu, W. Genomic insights into resistome, virulome, and mobilome as organic contaminants of ESKAPE pathogens and E. coli recovered from milk, farm workers, and environmental settings in Hainan, China. Emerg. Contam. 2024, 10, 100385. [Google Scholar] [CrossRef]
- Vlastarakos, P.; Nikolopoulos, T.P.; Maragoudakis, P.; Tzagaroulakis, A.; Ferekidis, E. Biofilms in ear, nose, and throat infections: How important are they? Laryngoscope 2007, 117, 668–673. [Google Scholar] [CrossRef]
- Secker, B.; Shaw, S.; Atterbury, R.J. Pseudomonas spp. in Canine Otitis Externa. Microorganisms 2023, 11, 2650. [Google Scholar] [CrossRef] [PubMed]
- Luciani, L.; Stefanetti, V.; Rampacci, E.; Valentini, P.L.; Capuozzo, R.; Passamonti, F. Comparison between clinical evaluations and laboratory findings and the impact of biofilm on antimicrobial susceptibility in vitro in canine otitis externa. Vet. Dermatol. 2023, 34, 586–596. [Google Scholar] [CrossRef]
- Paterson, S. Topical ear treatment—Options, indications and limitations of currenty therapy. J. Small Anim. Pract. 2016, 57, 668–678. [Google Scholar] [CrossRef]
- Viegas, F.M.; Santana, J.A.; Silva, B.A.; Xavier, R.G.C.; Bonisson, C.T.; Câmara, J.L.S.; Rennó, M.C.; Cunha, J.L.R.; Figueiredo, H.C.P.; Lobato, F.C.F.; et al. Occurrence and characterization of methicillin-resistant Staphylococcus spp. in diseased dogs in Brazil. PLoS ONE 2022, 17, e0269422. [Google Scholar] [CrossRef]
- Elfadadny, A.; Uchiyama, J.; Goto, K.; Imanishi, I.; Ragab, R.F.; Nageeb, W.M.; Iyori, K.; Toyoda, Y.; Tsukui, T.; Ide, K.; et al. Antimicrobial resistance and genotyping of Pseudomonas aeruginosa isolated from the ear canals of dogs in Japan. Front. Vet. Sci. 2023, 10, 1074127. [Google Scholar] [CrossRef]
- Scheinfeld, N.; Rosenberg, J.D.; Weinberg, J.M. Levamisole in Dermatology. Am. J. Clin. Dermatol. 2004, 5, 97–104. [Google Scholar] [CrossRef]
- Brilhante, R.S.N.; Caetano, E.P.; Lima, R.A.C.; Castelo Branco, D.S.C.M.; Serpa, R.; Oliveira, J.S.; Monteiro, A.J.; Rocha, M.F.G.; Cordeiro, R.A.; Sidrim, J.J.C. In vitro antifungal activity of miltefosine and levamisole: Their impct on ergosterol biosynthesis and cell permeability of dimorphic fungi. J. Appl. Microbiol. 2015, 119, 962–969. [Google Scholar] [CrossRef]
- Seleem, N.M.; Latif, H.K.A.; Shaldam, M.A.; El-Ganiny, A. Drugs with new lease of life as quórum sensing inhibitors: For combating MDR Acinetobacter baumannii infections. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1687–1702. [Google Scholar] [CrossRef] [PubMed]
- Favrot, C.; Steffan, J.; Seewald, W.; Picco, F. A prospective study on the clinical features of chronic canine atopic dermatitis and its diagnosis. Vet. Dermatol. 2010, 21, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Guedes, R.F.M.; Guedes, G.M.M.; Gomes, F.I.F.; Soares, A.C.C.F.; Pereira, V.C.; Freitas, A.S.; Amando, B.R.; Sidrim, J.J.C.; Cordeiro, R.A.; Rocha, M.F.G.R.; et al. Antimicrobial and antibiofilm effect of promethazine on bacterial isolates from canine otitis extern: An in vitro study. Microb. Pathog. 2024, 196, 106993. [Google Scholar] [CrossRef] [PubMed]
- CLSI M-100; Performance Standards for Antimicrobial Susceptibility Testing, Standards M02-A12, M07-A10, and M11-A8, 32th ed. CLSI M100-S32, Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022.
- Nuttall, T. Successful management of otitis externa. Practice 2016, 38 (Suppl. S2), 17–21. [Google Scholar] [CrossRef]
- Shoaib, M.; Tang, M.; Aqib, A.I.; Zhang, X.; Wu, Z.; Wen, Y.; Hou, X.; Xu, J.; Hao, R.; Wang, S.; et al. Dairy farm waste: A potential reservoir of diverse antibiotic resistance and virulence genes in aminoglycoside- and beta-lactam-resistant Escherichia coli in Gansu Province, China. Environ. Res. 2024, 263, 120190. [Google Scholar] [CrossRef]
- Shoaib, M.; Aqib, A.I.; Muzammil, I.; Majeed, N.; Bhutta, Z.A.; Kulyar, M.F.; Fatima, M.; Zaheer, C.-N.; Muneer, A.; Murtaza, M.; et al. MRSA compendium of epidemiology, transmission, pathophysiology, treatment, and prevention within one health framework. Front. Microbiol. 2023, 13, 1067284. [Google Scholar] [CrossRef]
- Costa, L.; Moreira, J.M.A.R.; Menezes, I.G.; Dutra, V.; Almeida, A.B.P.F. Antibiotic resistance profiles and activity of clove essential oil (Syzygium aromaticum) against Pseudomonas aeruginosa isolated of canine otitis. Vet. World. 2022, 15, 2499–2505. [Google Scholar] [CrossRef]
- World Health Organization. WHO Bacterial Priority Pathogens List, 2024: Bacterial Pathogens of Public Health Importance to Guide Research, Development and Strategies to Prevent and Control Antimicrobial Resistance; World Health Organization: Geneva, Switzerland, 2024; 72p, ISBN 978-92-4-009346-1. [Google Scholar]
- Scherer, C.B.; Botoni, L.S.; Bicalho, A.P.C. Resistência à meticilina em otite externa canina—Do diagnóstico ao tratamento. Medvep. Dermato. Rev. Educ. Contin. Dermatol. Alergol. Vet. 2014, 9, 224–233. [Google Scholar]
- Fontoura, E.G.; Valle, B.D.S.; Costa, A.L.; Capella, S.O.; Félix, S.R.; Mueller, E.N.; Nobre, M.O. Otite Externa em Pequenos Animais: Revisão de Literaatura. Medvep 2014, 12, 1–637. [Google Scholar]
- Ocanã, A.; Aguilera-Correa, J.J.; Domínguez-Jurado, E.; Pérez-Martínez, F.; Pérez-Tanoria, R.; López-Carretero, Y.; Masiá-Mondejar, J.; Castro-Osma, J.A.; Esteban, J.; Alonso-Moreno, C.; et al. A bis(pyrazolyl)methane derivated against clinical Sthaphylococcus aureus strains isolated from otitis externa. Laryngoscope Investig. Otolaryngol. 2022, 7, 283–290. [Google Scholar] [CrossRef]
- El-Gammal, G.A.; El-Gamal, A.M.; Rashed, M.A.; Kassab, A.S.; Saif, A.S.; Fadl, S.E. An experimental study of levamisole incorporated diet on fish health and resistance against Pseudomonas aeruginosa isolated from Oreochromis niloticus. Sci. Rep. 2025, 15, 14658. [Google Scholar] [CrossRef] [PubMed]
- Srikala, D.; Kumar, K.S.; Kumar, V.V.V.; Nagaraj, P.; Ayodhya, S.; Rao, D.S. Management of Mixed Infestation of Malasseziosis and Demodicosis in Dogs. Intas. Polivet. 2010, 11, 74–76. [Google Scholar]
- Gholami, M.H.; Rassouli, A.; Mirzaei, S.; Hashemi, F. The potential immunomodulatory effect of levamisole in humans and farm animals. J. Adv. Vet. Anim. Res. 2023, 10, 620–629. [Google Scholar] [CrossRef]
- Kalidari, G.A.; Soudkolaei, S.S.; Borji, H. Anthelmintic efficacy of fenbendazole and levamisole in native fowl in northern Iran. Parasites Vectors 2021, 14, 104. [Google Scholar] [CrossRef]
- Martin, R.J.; Robertson, A.P.; Buxton, S.K.; Beech, R.N.; Charvet, C.L.; Neveu, C. Levamisole receptors: A second awakening. Trends Parasitol. 2012, 28, 289–296. [Google Scholar] [CrossRef]
- Pfarr, K.M.; Krome, A.K.; Al-Obaidi, I.; Batchelor, H.; Vaillant, M.; Hoerauf, A.; Opoku, N.O.; Kuesel, A.C. The pipeline for drugs for control and elimination of neglected tropical diseases: 2. oral anti-infective drugs and drug combinations for off-label use. Parasites Vectors 2023, 16, 394. [Google Scholar] [CrossRef] [PubMed]
- Stogaus, R.; King, M.G. Is oral levamisole immunoestimulation in rats mediated by reduced levels of free plasma corticosterone? Int. J. Immunopharmacol. 1995, 17, 635–640. [Google Scholar] [CrossRef]
- Nesse, L.L.; Osland, A.M.; Vestby, L.K. The role of biofilms in the pathogenesis of animal bacterial infections. Microorganisms 2023, 11, 608. [Google Scholar] [CrossRef]
- Rabin, N.; Zheng, Y.; Opoku-Temeng, Y.D.; Bonsu, E.; Sintim, H.O. Biofilm formation mechanisms and targets for developing antibiofilm agentes. Future Med. Chem. 2015, 7, 493–512. [Google Scholar] [CrossRef]
- Chan, W.Y.; Hickey, E.E.; Page, S.W.; Trott, D.J.; Hill, P.B. Biofilm production by pathogens associated with canine otitis externa, and the antibiofilm activity of ionophores and antimicrobial adjuvants. J. Vet. Pharmacol. Ther. 2019, 42, 682–692. [Google Scholar] [CrossRef]
- Pati, B.A.; Kurata, W.E.; Horseman, T.S.; Pierce, L.M. Antibiofilm activity of chitosan/epsilon-poly-L-lysine hydrogels in a porcine ex vivo skin wound polymicrobial biofilm model. Wound Repair. Regen. 2021, 29, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Papich, M.G. Sounders Handbookof Veterinary Drugs Small and Large Animal, 4th ed.; Saunders: Philadelphia, PA, USA, 2015. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | N | Resistance a | MDR | Drugs | Biofilm-Forming Ability |
|---|---|---|---|---|---|
| Staphylococcus pseudintermedius (n = 11) | 5 | 10–13 | Yes | OXA *, CDM, CIP, ERI, GEN, LVX, STX, TET | Moderate |
| 3 | 5–7 | Yes | CDM, CIP, ERI, GEN, LVX, STX, TET | Strong/Moderate | |
| 3 | 1–2 | No | AMP, CDM, STX | Strong/Moderate | |
| Staphylococcus schleiferi (n = 3) | 1 | 7 | No | OXA *, CDM | Moderate |
| 2 | 0–1 | No | CDM | Strong/Moderate | |
| Staphylococcus hominis (n = 2) | 1 | 1 | No | CDM | Strong |
| 1 | 6 | Yes | CDM, STX, TEI, VAN | Moderate | |
| Staphylococcus aureus (n = 1) | 1 | 5 | Yes | CDM, CIP, LVX, STX, TET | Moderate |
| Staphylococcus capitis (n = 1) | 1 | 1 | No | CDM | Strong |
| Staphylococcus cohnii (n = 1) | 1 | 1 | No | CDM | Strong |
| Staphylococcus auricularis (n = 1) | 1 | 6 | Yes | CDM, LVX, STX, VAN | Weak |
| Staphylococcus epidermitis (n = 1) | 1 | 13 | Yes | AMO/SUL, AMP, AMC, CAX, CDM, CIP, DAP, ERI, GEN, LVX, LNZ, OXA, PEN, RIF, STX, TEI, TET, VAN | Moderate |
| Enterococcus faecalis (n = 3) | 2 | 2 | No | TET | Strong/Moderate |
| 1 | 4 | Yes | ERI, STR, TET | Strong | |
| Proteus mirabilis (n = 12) | 6 | 0 | No | - | Weak/Moderate/Strong |
| 1 | 8 | No | AMP, AMC, FEP, ETP, MER, PRL/TAZ, PRL | Moderate | |
| 2 | 4 | Yes | AMP, FOS, PRL, STX | Moderate | |
| 3 | 2–3 | No | AMP, FOS, PRL, STX | Moderate/Strong | |
| Pseudomonas aeruginosa (n = 11) | 6 | 0 | No | - | Weak/Moderate/Strong |
| 5 | 1–2 | No | GEN, IMI, LVX | Moderate/Strong | |
| Escherichia coli (n = 3) | 2 | 0 | No | - | Weak |
| 1 | 8 | Yes | AMP, AMC, AZT, CAX, CIP, CEFU, PRL, STX | Weak |
| Species (n) * | MIC (mg/mL) | MBIC (mg/mL) | MBEC (mg/mL) |
|---|---|---|---|
| Staphylococcus pseudintermedius (11) | 1.17 (5) | 2.34 (3) | 4.68 (1) |
| 0.58 (6) | 4.68 (2) | 9.37 (1) | |
| 9.37 (1) | 18.75 (2) | ||
| 18.75 (5) | >18.75 (7) | ||
| Staphylococcus schleiferi (3) | 1.17 (3) | 2.34 (2) | 4.68 (2) |
| 4.68 (1) | 9.37 (1) | ||
| Staphylococcus hominis (2) | 1.17 (2) | 2.34 (1) | 4.68 (1) |
| 18.75 (1) | >18.75 (1) | ||
| Staphylococcus aureus (1) | 0.58 | 2.34 | >18.75 |
| Staphylococcus capitis (1) | 0.58 | 9.37 | 18.75 |
| Staphylococcus cohnii (1) | 1.17 | 9.37 | 18.75 |
| Staphylococcus auricularis (1) | 0.58 | 18.75 | >18.75 |
| Staphylococcus epidermitis (1) | 1.17 | 9.37 | 18.75 |
| Enterococcus faecalis (3) | 2.34 (3) | 4.68 (1) | 18.75 (2) |
| 9.37 (2) | >18.75 (1) | ||
| Proteus mirabilis (12) | 1.17 (3) | 2.34 (1) | 4.68 (1) |
| 2.34 (9) | 4.68 (2) | 9.37 (1) | |
| 9.37 (8) | 18.75 (6) | ||
| 18.75 (1) | >18.75 (4) | ||
| Pseudomonas aeruginosa (11) | 1.17 (2) | 4.68 (4) | >18.75 (11) |
| 2.34 (9) | 9.37 (4) | ||
| 18.75 (3) | |||
| Escherichia coli (3) | 1.17 (3) | 4.68 (3) | 9.37 (1) |
| >18.75 (2) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guedes, R.F.M.; Soares, A.C.C.F.; Gomes, F.I.F.; Freitas, A.S.; Pereira, V.C.; Cordeiro, R.A.; Rocha, M.F.G.; Sidrim, J.J.C.; Barbosa, G.R.; Guedes, G.M.M.; et al. Levamisole as a Strategy Against Bacteria from Canine Otitis Externa: An In Vitro Evaluation. Vet. Sci. 2025, 12, 640. https://doi.org/10.3390/vetsci12070640
Guedes RFM, Soares ACCF, Gomes FIF, Freitas AS, Pereira VC, Cordeiro RA, Rocha MFG, Sidrim JJC, Barbosa GR, Guedes GMM, et al. Levamisole as a Strategy Against Bacteria from Canine Otitis Externa: An In Vitro Evaluation. Veterinary Sciences. 2025; 12(7):640. https://doi.org/10.3390/vetsci12070640
Chicago/Turabian StyleGuedes, Rodrigo F. M., Ana C. C. F. Soares, Francisco I. F. Gomes, Alyne S. Freitas, Vinicius C. Pereira, Rossana A. Cordeiro, Marcos F. G. Rocha, José J. C. Sidrim, Giovanna R. Barbosa, Glaucia M. M. Guedes, and et al. 2025. "Levamisole as a Strategy Against Bacteria from Canine Otitis Externa: An In Vitro Evaluation" Veterinary Sciences 12, no. 7: 640. https://doi.org/10.3390/vetsci12070640
APA StyleGuedes, R. F. M., Soares, A. C. C. F., Gomes, F. I. F., Freitas, A. S., Pereira, V. C., Cordeiro, R. A., Rocha, M. F. G., Sidrim, J. J. C., Barbosa, G. R., Guedes, G. M. M., & Castelo-Branco, D. S. C. M. (2025). Levamisole as a Strategy Against Bacteria from Canine Otitis Externa: An In Vitro Evaluation. Veterinary Sciences, 12(7), 640. https://doi.org/10.3390/vetsci12070640