
Meloxicam Alleviates Oxidative Stress Through Nrf2/HO-1 Activation in Bovine Endometrial Epithelial Cells

 ,
, 
 ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Drugs and Reagents
2.2. Cell Culture
2.3. Treatment Design
2.4. Cell Viability
2.5. Flow Cytometry Analysis of ROS
2.6. Determination of the MDA, SOD, CAT, and GSH Levels
2.7. Real-Time Quantitative PCR
2.8. Western Blot Analysis
2.9. Immunofluorescence Staining
2.10. Statistical Analysis
3. Results
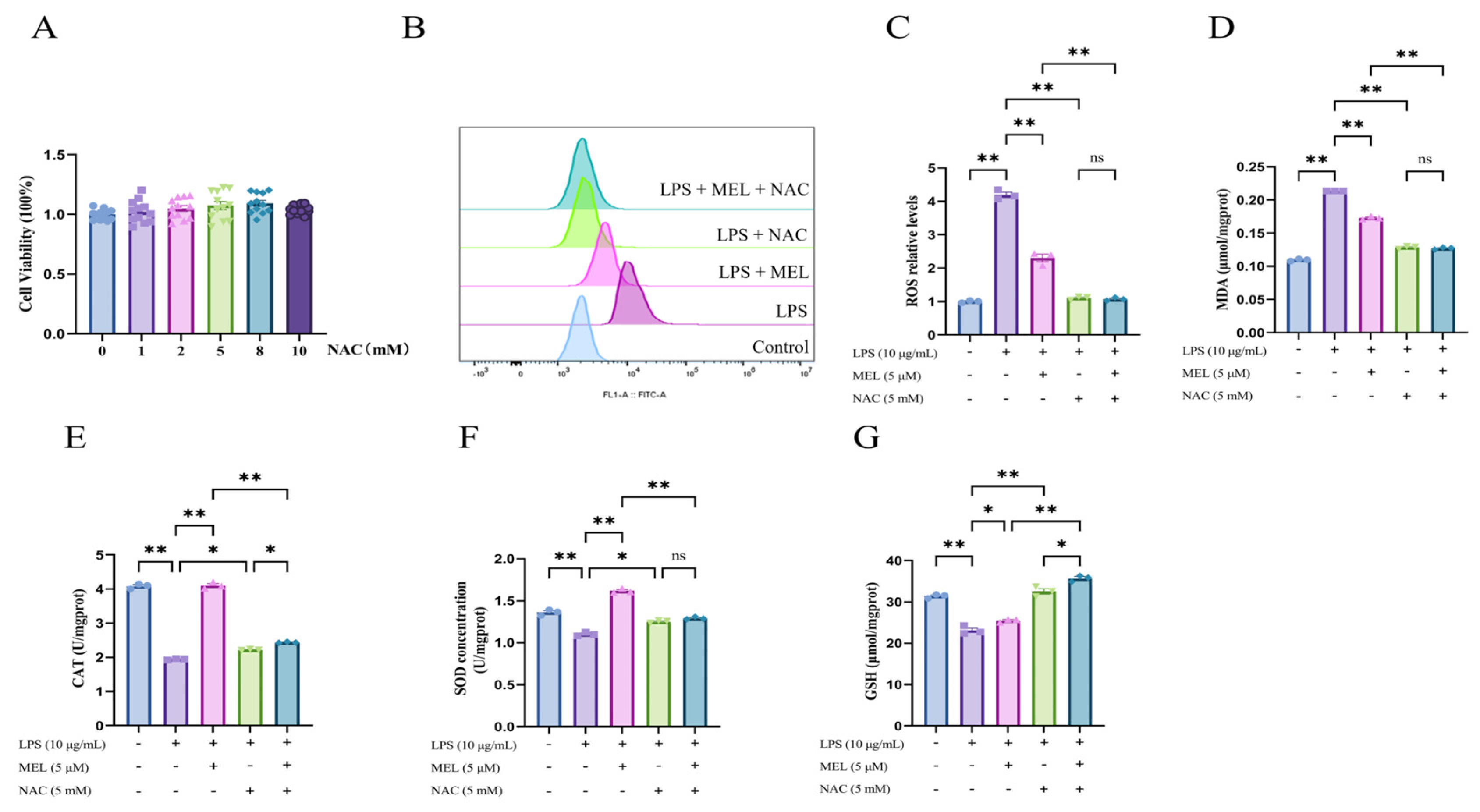
3.1. Meloxicam Inhibited ROS Accumulation and Increased the Levels of Antioxidant Indicators
3.2. Meloxicam Alleviated BEEC Oxidative Stress by Activating the Nrf2 Pathway
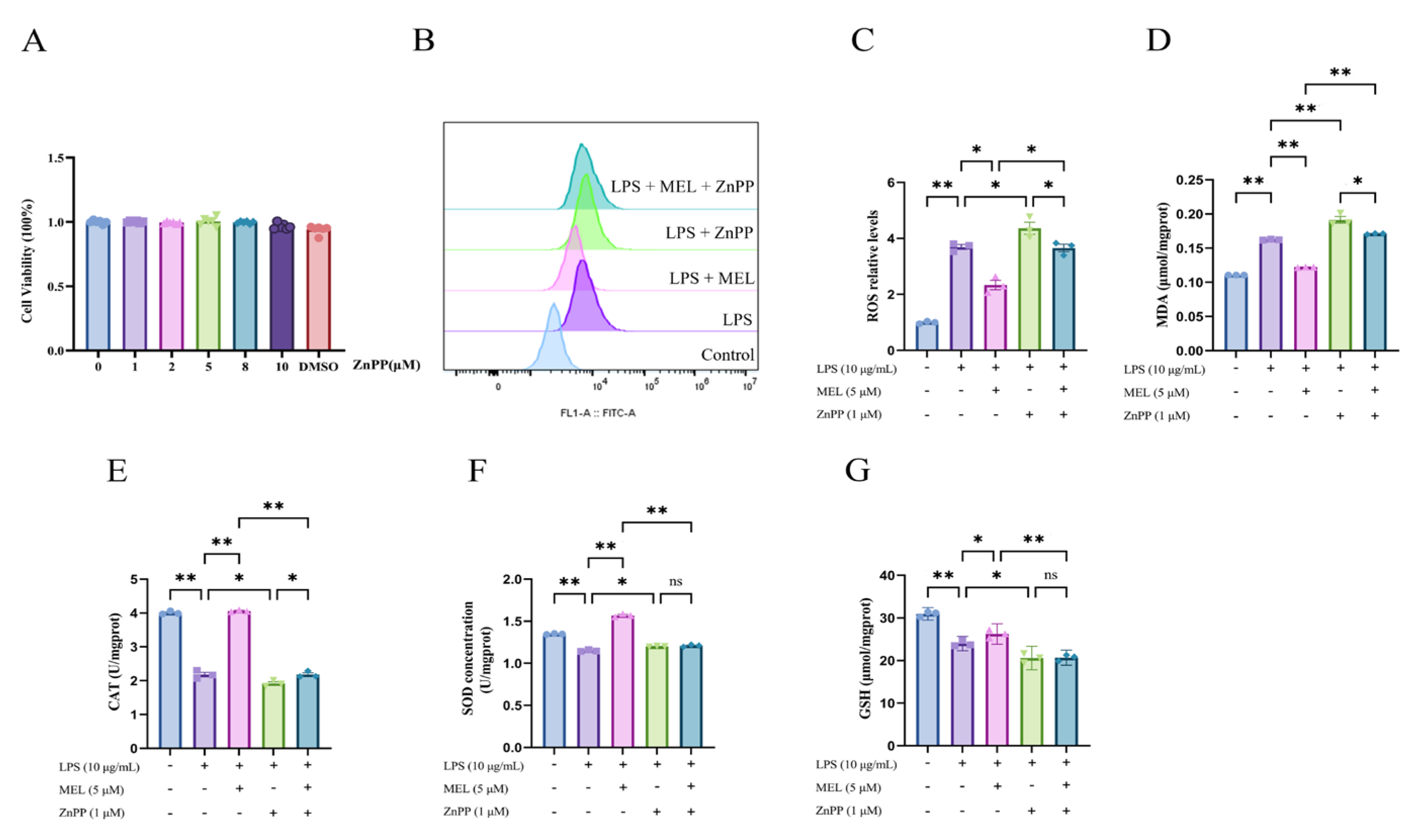
3.3. HO-1 Inhibitor Blunted the Antioxidant Effect of Meloxicam
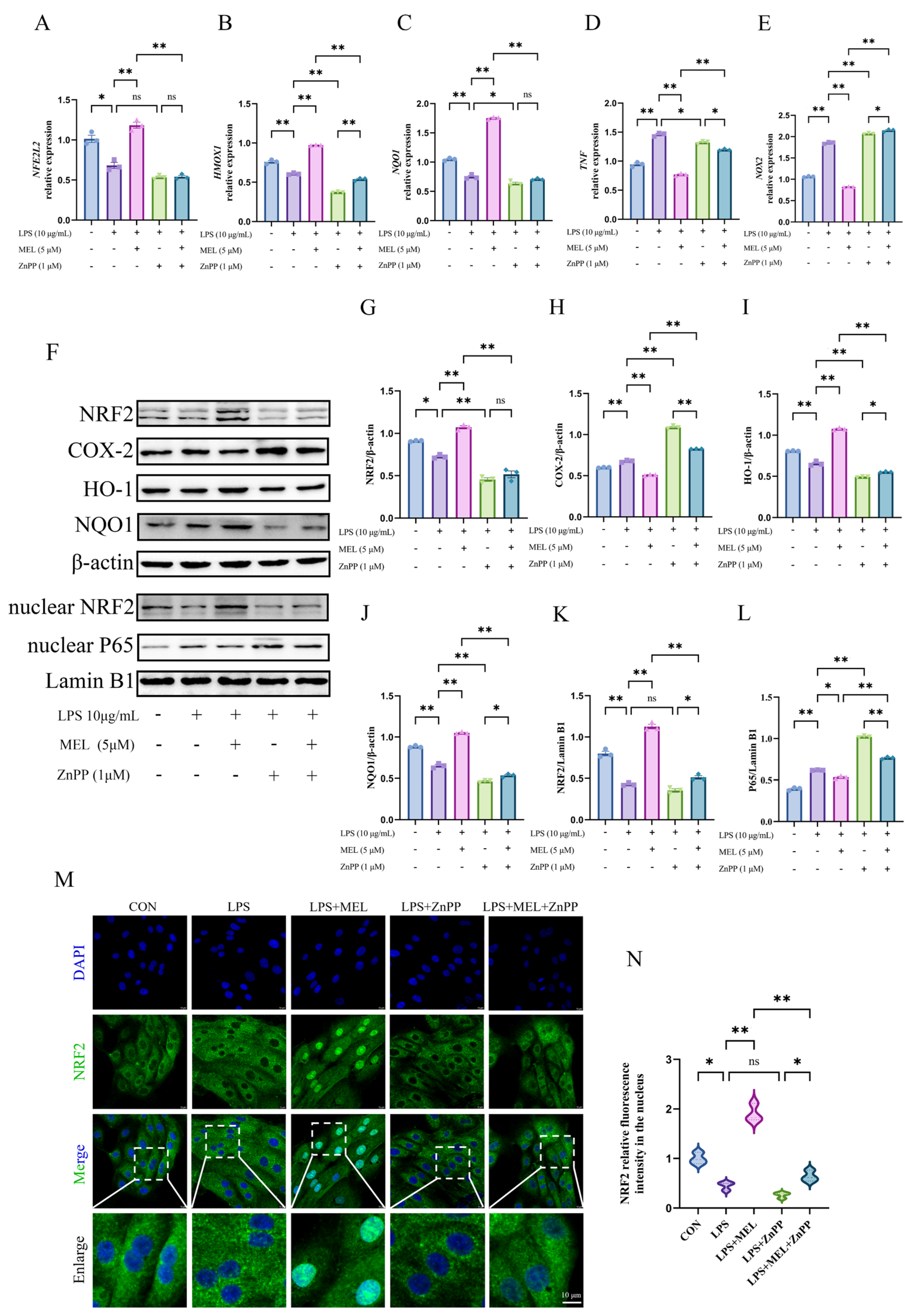
3.4. HO-1 Inhibitor Weakened Meloxicam-Induced Nrf2 Activation
3.5. Nrf2 Inhibitor Abolished Meloxicam Antioxidation
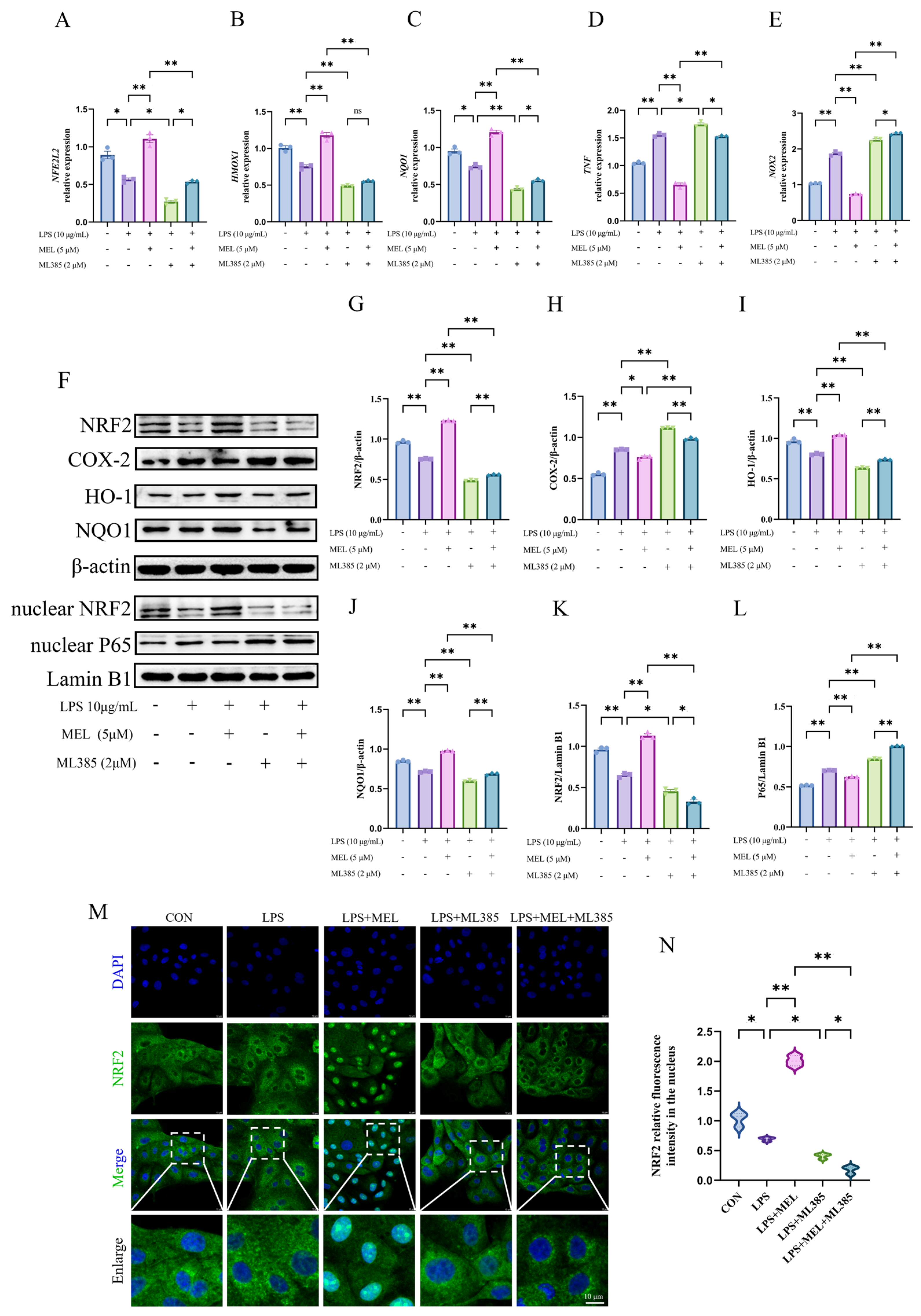
3.6. Nrf2 Inhibitor Reversed Meloxicam-Induced Nrf2 Activation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BEEC | Bovine endometrial epithelial cell |
| COX-2 | Cyclooxygenase-2 |
| CAT | Catalase |
| ERK | Extracellular regulated protein kinases |
| GSH | Glutathione |
| HMOX1/HO-1 | Bos taurus heme oxygenase 1 |
| LPS | Lipopolysaccharide |
| MDA | Malondialdehyde |
| NSAID | Non-steroidal anti-inflammatory drugs |
| NF-κB | Nuclear factor kappa-B |
| NFE2L2/Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| NQO1 | NAD(P)H quinone oxidoreductase 1 |
| NOX2 | NADPH oxidase 2 |
| NAC | N-acetylcysteine |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| TNF | Tumor necrosis factor |
| ZnPP | Zinc protoporphyrin |
References
- Druker, S.A.; Sicsic, R.; van Straten, M.; Goshen, T.; Kedmi, M.; Raz, T. Cytological endometritis diagnosis in primiparous versus multiparous dairy cows. J. Dairy Sci. 2022, 105, 665–683. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, I.M.; Dobson, H. Postpartum uterine health in cattle. Anim. Reprod. Sci. 2004, 82–83, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, I.M.; Noakes, D.E.; Rycroft, A.N.; Dobson, H. The effect of intrauterine administration of estradiol on postpartum uterine involution in cattle. Theriogenology 2003, 59, 1357–1371. [Google Scholar] [CrossRef] [PubMed]
- Bradford, B.J.; Yuan, K.; Farney, J.K.; Mamedova, L.K.; Carpenter, A.J. Invited review: Inflammation during the transition to lactation: New adventures with an old flame. J. Dairy Sci. 2015, 98, 6631–6650. [Google Scholar] [CrossRef]
- de la Puente, R.; Diez, R.; Diez, M.J.; Fernandez, N.; Sahagun, A.M.; Rodriguez, J.M.; Garcia, J.J.; Lopez, C. Pharmacokinetics of meloxicam in different animal species: A comprehensive review. Vet. Sci. 2024, 11, 519. [Google Scholar] [CrossRef]
- Offinger, J.; Herdtweck, S.; Rizk, A.; Starke, A.; Heppelmann, M.; Meyer, H.; Janssen, S.; Beyerbach, M.; Rehage, J. Postoperative analgesic efficacy of meloxicam in lame dairy cows undergoing resection of the distal interphalangeal joint. J. Dairy Sci. 2013, 96, 866–876. [Google Scholar] [CrossRef]
- Fitzpatrick, C.E.; Chapinal, N.; Petersson-Wolfe, C.S.; DeVries, T.J.; Kelton, D.F.; Duffield, T.F.; Leslie, K.E. The effect of meloxicam on pain sensitivity, rumination time, and clinical signs in dairy cows with endotoxin-induced clinical mastitis. J. Dairy Sci. 2013, 96, 2847–2856. [Google Scholar] [CrossRef]
- Mehra, U.S.; Arya, S.; Kadian, P.; Chaudhary, R.; Yadav, R.P.; Kumar, P. Evaluation of meloxicam as stress modulator in cases of assisted parturition in buffaloes. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1805–1812. [Google Scholar] [CrossRef]
- Lomb, J.; Neave, H.W.; Weary, D.M.; LeBlanc, S.J.; Huzzey, J.M.; von Keyserlingk, M.A.G. Changes in feeding, social, and lying behaviors in dairy cows with metritis following treatment with a nonsteroidal anti-inflammatory drug as adjunctive treatment to an antimicrobial. J. Dairy Sci. 2018, 101, 4400–4411. [Google Scholar] [CrossRef]
- Pascottini, O.B.; Van Schyndel, S.J.; Spricigo, J.F.W.; Carvalho, M.R.; Mion, B.; Ribeiro, E.S.; LeBlanc, S.J. Effect of anti-inflammatory treatment on systemic inflammation, immune function, and endometrial health in postpartum dairy cows. Sci. Rep. 2020, 10, 5236. [Google Scholar] [CrossRef]
- Ogino, K.; Saito, K.; Osugi, T.; Satoh, H. Meloxicam (Mobic): A review of its pharmacological and clinical profile. Nihon Yakurigaku Zasshi. Folia Pharmacol. Jpn. 2002, 120, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Caldeira, M.O.; Bruckmaier, R.M.; Wellnitz, O. Meloxicam affects the inflammatory responses of bovine mammary epithelial cells. J. Dairy Sci. 2019, 102, 10277–10290. [Google Scholar] [CrossRef]
- Cui, L.; Guo, J.; Wang, Z.; Zhang, J.; Li, W.; Dong, J.; Liu, K.; Guo, L.; Li, J.; Wang, H.; et al. Meloxicam inhibited oxidative stress and inflammatory response of LPS-stimulated bovine endometrial epithelial cells through Nrf2 and NF-κB pathways. Int. Immunopharmacol. 2023, 116, 109822. [Google Scholar] [CrossRef] [PubMed]
- Artimovič, P.; Badovská, Z.; Toporcerová, S.; Špaková, I.; Smolko, L.; Sabolová, G.; Kriváková, E.; Rabajdová, M. Oxidative stress and the Nrf2/PPARγ axis in the endometrium: Insights into female fertility. Cells 2024, 13, 1081. [Google Scholar] [CrossRef]
- Xue, D.; Zhou, X.; Qiu, J. Emerging role of NRF2 in ROS-mediated tumor chemoresistance. Biomed. Pharmacother. 2020, 131, 110676. [Google Scholar] [CrossRef]
- Kopacz, A.; Kloska, D.; Forman, H.J.; Jozkowicz, A.; Grochot-Przeczek, A. Beyond repression of Nrf2: An update on Keap1. Free. Radic. Biol. Med. 2020, 157, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.H.; Ghobara, M.M. Antifibrotic effect of meloxicam in rat liver: Role of nuclear factor kappa B, proinflammatory cytokines, and oxidative stress. Naunyn Schmiedebergs Arch. Pharmacol. 2016, 389, 971–983. [Google Scholar] [CrossRef]
- Yang, W.; Tiffany-Castiglioni, E.; Lee, M.Y.; Son, I.H. Paraquat induces cyclooxygenase-2 (COX-2) implicated toxicity in human neuroblastoma SH-SY5Y cells. Toxicol. Lett. 2010, 199, 239–246. [Google Scholar] [CrossRef]
- Arab, H.H.; Khames, A.; Mohammad, M.K.; Alsufyani, S.E.; Ashour, A.M.; El-Sheikh, A.A.K.; Darwish, H.W.; Gad, A.M. Meloxicam targets COX-2/NOX1/NOX4/Nrf2 axis to ameliorate the depression-like neuropathology induced by chronic restraint stress in rats. Pharmaceuticals 2023, 16, 848. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Lewis, G.S.; LeBlanc, S.; Gilbert, R.O. Defining postpartum uterine disease in cattle. Theriogenology 2006, 65, 1516–1530. [Google Scholar] [CrossRef]
- Dong, J.; Qu, Y.; Li, J.; Cui, L.; Wang, Y.; Lin, J.; Wang, H. Cortisol inhibits NF-κB and MAPK pathways in LPS activated bovine endometrial epithelial cells. Int. Immunopharmacol. 2018, 56, 71–77. [Google Scholar] [CrossRef]
- Cui, L.; Wang, H.; Lin, J.; Wang, Y.; Dong, J.; Li, J.; Li, J. Progesterone inhibits inflammatory response in E. coli- or LPS-Stimulated bovine endometrial epithelial cells by NF-κB and MAPK pathways. Dev. Comp. Immunol. 2020, 105, 103568. [Google Scholar] [CrossRef]
- Maślanka, T. Effect of dexamethasone and meloxicam on counts of selected T lymphocyte subpopulations and NK cells in cattle-In vivo investigations. Res. Vet. Sci. 2014, 96, 338–346. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Y.N.; Zhu, J.J.; Liu, X.X.; You, H.; Gong, M.Y.; Zou, M.; Cheng, W.H.; Zhu, J.H. N-acetylcysteine negatively regulates Notch3 and its malignant signaling. Oncotarget 2016, 7, 30855–30866. [Google Scholar] [CrossRef]
- Sun, W.S.; Jang, H.; Park, M.R.; Oh, K.B.; Lee, H.; Hwang, S.; Xu, L.J.; Hwang, I.S.; Lee, J.W. N-acetyl-L-cysteine improves the developmental competence of bovine oocytes and embryos cultured in vitro by attenuating oxidative damage and apoptosis. Antioxidants 2021, 10, 860. [Google Scholar] [CrossRef]
- Fu, K.; Feng, C.; Shao, L.; Mei, L.; Cao, R. Tanshinone IIA exhibits anti-inflammatory and antioxidative effects in LPS-stimulated bovine endometrial epithelial cells by activating the Nrf2 signaling pathway. Res. Vet. Sci. 2021, 136, 220–226. [Google Scholar] [CrossRef]
- Fu, K.; Chen, H.; Mei, L.; Wang, J.; Gong, B.; Li, Y.; Cao, R. Berberine enhances autophagic flux by activating the Nrf2 signaling pathway in bovine endometrial epithelial cells to resist LPS-induced apoptosis. Anim. Sci. J. 2023, 94, e13847. [Google Scholar] [CrossRef]
- Cui, L.; Zhong, J.; Duan, J.; Li, W.; Mao, P.; Dong, J.; Liu, K.; Guo, L.; Wang, H.; Li, J. The antioxidant effect of selenium is enhanced by cortisol through Nrf2 pathway in bovine endometrial epithelial cells. Animals 2025, 15, 1075. [Google Scholar] [CrossRef]
- Fu, K.; Wang, Z.; Cao, R. Berberine attenuates the inflammatory response by activating the Keap1/Nrf2 signaling pathway in bovine endometrial epithelial cells. Int. Immunopharmacol. 2021, 96, 107738. [Google Scholar] [CrossRef]
- Song, P.; Liu, C.; Sun, M.; Liu, J.; Lin, P.; Chen, H.; Zhou, D.; Tang, K.; Wang, A.; Jin, Y. Transcription factor Nrf2 modulates lipopolysaccharide-induced injury in bovine endometrial epithelial cells. Int. J. Mol. Sci. 2023, 24, 11221. [Google Scholar] [CrossRef]
- Moldéus, P.; Cotgreave, I.A.; Berggren, M. Lung protection by a thiol-containing antioxidant: N-acetylcysteine. Respiration 1986, 50 (Suppl. 1), 31–42. [Google Scholar] [CrossRef]
- Sahasrabudhe, S.A.; Terluk, M.R.; Kartha, R.V. N-acetylcysteine pharmacology and applications in rare diseases-repurposing an old antioxidant. Antioxidants 2023, 12, 1316. [Google Scholar] [CrossRef]
- Pedre, B.; Barayeu, U.; Ezeriņa, D.; Dick, T.P. The mechanism of action of N-acetylcysteine (NAC): The emerging role of H(2)S and sulfane sulfur species. Pharmacol. Ther. 2021, 228, 107916. [Google Scholar] [CrossRef]
- Cui, L.; Qu, Y.; Cai, H.; Wang, H.; Dong, J.; Li, J.; Qian, C.; Li, J. Meloxicam inhibited the proliferation of LPS-stimulated bovine endometrial epithelial cells through Wnt/β-Catenin and PI3K/AKT pathways. Front. Vet. Sci. 2021, 8, 637707. [Google Scholar] [CrossRef]
- Yang, Y.; He, Q.; Wang, H.; Hu, X.; Luo, Y.; Liang, G.; Kuang, S.; Mai, S.; Ma, J.; Tian, X.; et al. The protection of meloxicam against chronic aluminium overload-induced liver injury in rats. Oncotarget 2017, 8, 23448–23458. [Google Scholar] [CrossRef]
- Pyun, B.J.; Jo, K.; Lee, J.Y.; Lee, A.; Jung, M.A.; Hwang, Y.H.; Jung, D.H.; Ji, K.Y.; Choi, S.; Kim, Y.H.; et al. Caesalpinia sappan Linn. ameliorates allergic nasal inflammation by upregulating the Keap1/Nrf2/HO-1 pathway in an allergic rhinitis mouse model and nasal epithelial cells. Antioxidants 2022, 11, 2256. [Google Scholar] [CrossRef]
- Labbé, R.F.; Vreman, H.J.; Stevenson, D.K. Zinc protoporphyrin: A metabolite with a mission. Clin. Chem. 1999, 45, 2060–2072. [Google Scholar] [CrossRef]
- Wang, Y.; Qiu, J.; Yan, H.; Zhang, N.; Gao, S.; Xu, N.; Wang, C.; Lou, H. The Bach1/HO-1 pathway regulates oxidative stress and contributes to ferroptosis in doxorubicin-induced cardiomyopathy in H9c2 cells and mice. Arch. Toxicol. 2024, 98, 1781–1794. [Google Scholar] [CrossRef]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef]
- Qiu, W.; An, S.; Wang, T.; Li, J.; Yu, B.; Zeng, Z.; Chen, Z.; Lin, B.; Lin, X.; Gao, Y. Melatonin suppresses ferroptosis via activation of the Nrf2/HO-1 signaling pathway in the mouse model of sepsis-induced acute kidney injury. Int. Immunopharmacol. 2022, 112, 109162. [Google Scholar] [CrossRef]
- Liu, M.; Guan, G.; Wang, Y.; Lu, X.; Duan, X.; Xu, X. p-Hydroxy benzaldehyde, a phenolic compound from Nostoc commune, ameliorates DSS-induced colitis against oxidative stress via the Nrf2/HO-1/NQO-1/NF-κB/AP-1 pathway. Phytomedicine 2024, 133, 155941. [Google Scholar] [CrossRef]
- Alam, J.; Stewart, D.; Touchard, C.; Boinapally, S.; Choi, A.M.; Cook, J.L. Nrf2, a Cap’n’Collar transcription factor, regulates induction of the heme oxygenase-1 gene. J. Biol. Chem. 1999, 274, 26071–26078. [Google Scholar] [CrossRef]
- Pae, H.O.; Chung, H.T. Heme oxygenase-1: Its therapeutic roles in inflammatory diseases. Immune Netw. 2009, 9, 12–19. [Google Scholar] [CrossRef]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef]
- Wei, Z.; Jing, Z.; Pinfang, K.; Chao, S.; Shaohuan, Q. Quercetin inhibits pyroptosis in diabetic cardiomyopathy through the Nrf2 pathway. J. Diabetes Res. 2022, 2022, 9723632. [Google Scholar] [CrossRef]
- Meng, M.; Huo, R.; Wang, Y.; Ma, N.; Shi, X.; Shen, X.; Chang, G. Lentinan inhibits oxidative stress and alleviates LPS-induced inflammation and apoptosis of BMECs by activating the Nrf2 signaling pathway. Int. J. Biol. Macromol. 2022, 222, 2375–2391. [Google Scholar] [CrossRef]
- Li, Y.; Feng, Y.F.; Liu, X.T.; Li, Y.C.; Zhu, H.M.; Sun, M.R.; Li, P.; Liu, B.; Yang, H. Songorine promotes cardiac mitochondrial biogenesis via Nrf2 induction during sepsis. Redox Biol. 2021, 38, 101771. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (5′ → 3′) | Length (bp) | NCBI Number |
|---|---|---|---|
| ACTB | F: CATCACCATCGGCAATGAGC R: AGCACCGTGTTGGCGTAGAG | 156 | NM_173979.3 |
| NFE2L2 | F: CCCAGTCTTCACTGCTCCTC R: TCAGCCAGCTTGTCATTTTG | 165 | NM_001011678.2 |
| HMOX1 | F: GGCAGCAAGGTGCAAGA R: GAAGGAAGCCAGCCAAGAG | 221 | NM_001014912.1 |
| NQO1 | F: AACCAACAGACCAGCCAATC R: CACAGTGACCTCCCATCCTT | 154 | NM_001034535.1 |
| TNF | F: GGACACCCAGAATGTGAGGG R: GGAGAGTTGAAGTCCACGCA | 102 | NM_173966.3 |
| NOX2 | F: CTCAGCTACAACATCTGCCTCACT R: CTGTGATTACATCTTTCTCCTCGTCAT | 91 | NM_174035.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, L.; Duan, J.; Mao, P.; Zhong, J.; He, S.; Dong, J.; Liu, K.; Guo, L.; Li, J.; Wang, H. Meloxicam Alleviates Oxidative Stress Through Nrf2/HO-1 Activation in Bovine Endometrial Epithelial Cells. Vet. Sci. 2025, 12, 579. https://doi.org/10.3390/vetsci12060579
Cui L, Duan J, Mao P, Zhong J, He S, Dong J, Liu K, Guo L, Li J, Wang H. Meloxicam Alleviates Oxidative Stress Through Nrf2/HO-1 Activation in Bovine Endometrial Epithelial Cells. Veterinary Sciences. 2025; 12(6):579. https://doi.org/10.3390/vetsci12060579
Chicago/Turabian StyleCui, Luying, Jiangyao Duan, Peng Mao, Jingyi Zhong, Sasa He, Junsheng Dong, Kangjun Liu, Long Guo, Jianji Li, and Heng Wang. 2025. "Meloxicam Alleviates Oxidative Stress Through Nrf2/HO-1 Activation in Bovine Endometrial Epithelial Cells" Veterinary Sciences 12, no. 6: 579. https://doi.org/10.3390/vetsci12060579
APA StyleCui, L., Duan, J., Mao, P., Zhong, J., He, S., Dong, J., Liu, K., Guo, L., Li, J., & Wang, H. (2025). Meloxicam Alleviates Oxidative Stress Through Nrf2/HO-1 Activation in Bovine Endometrial Epithelial Cells. Veterinary Sciences, 12(6), 579. https://doi.org/10.3390/vetsci12060579