

Role of Microencapsulated Essential Oil and Pepper Resin in the Diet of Cows in the Third Lactation Phase on Immunological Pathways

 , , ,
, , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Additive
2.2. Experimental Condition
2.3. Experimental Design and Feed
2.4. Sample Collection
2.5. Hemogram
2.6. Serum Biochemistry
2.7. Cytokines—Interleukin 1β, Interleukin 6, and Interleukin 10
2.8. TBARS Levels
2.9. Reactive Oxygen Species (ROS)
2.10. Levels of Total Thiol (PSH) and Non-Protein Thiol (NPSH)
2.11. Glutathione S-Transferase (GST) Activity
2.12. Purinergic Signaling: Separation of Lymphocytes and Platelets
2.13. Enzymatic Energetic Metabolism
2.14. Nutrient Analysis
2.15. Productive Performance
2.16. Milk Analysis
2.17. Statistical Analyses
3. Results
3.1. Blood Count and Serum Biochemistry
3.2. Levels of Cytokines and Oxidative Status
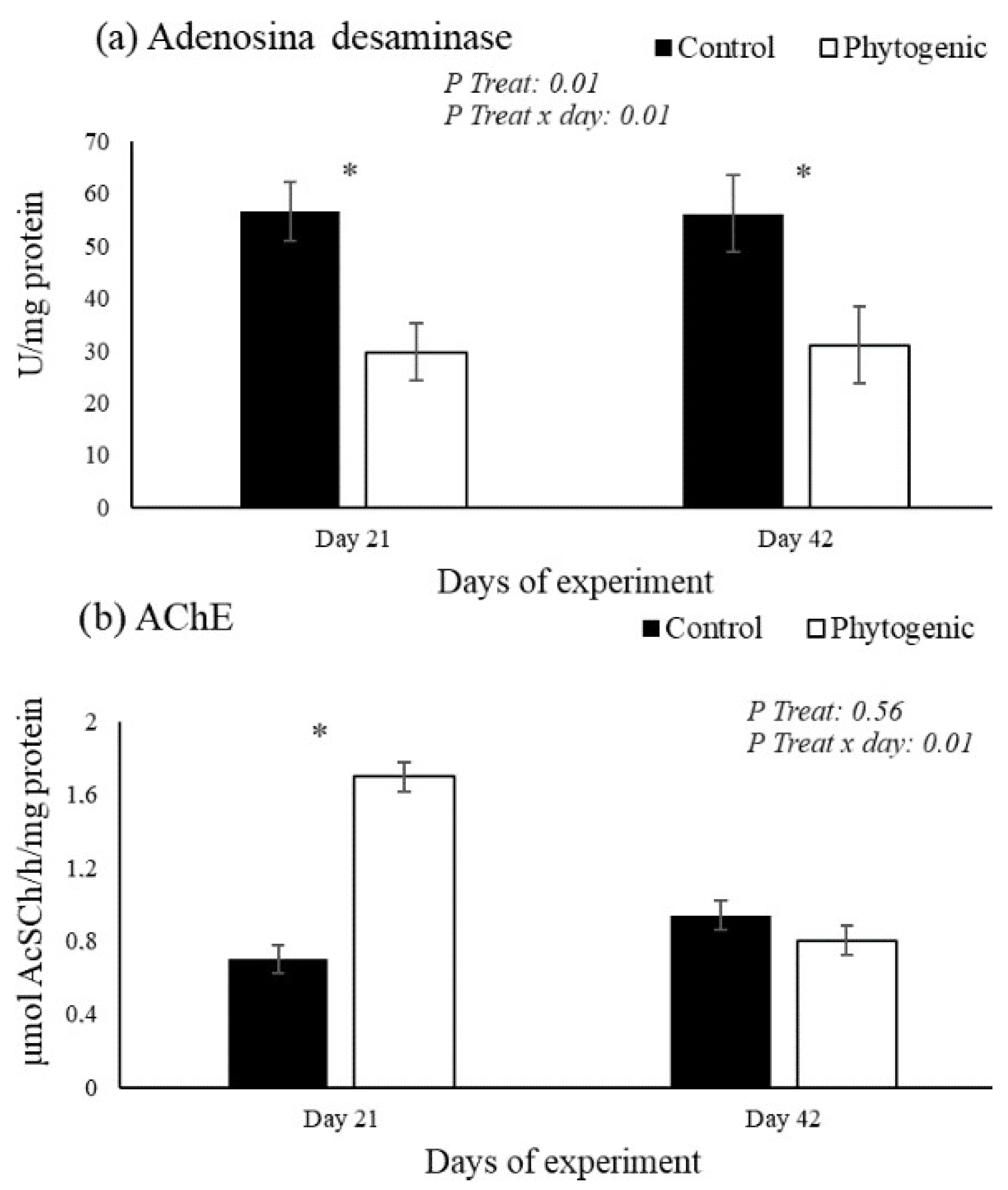
3.3. Purinergic Signaling
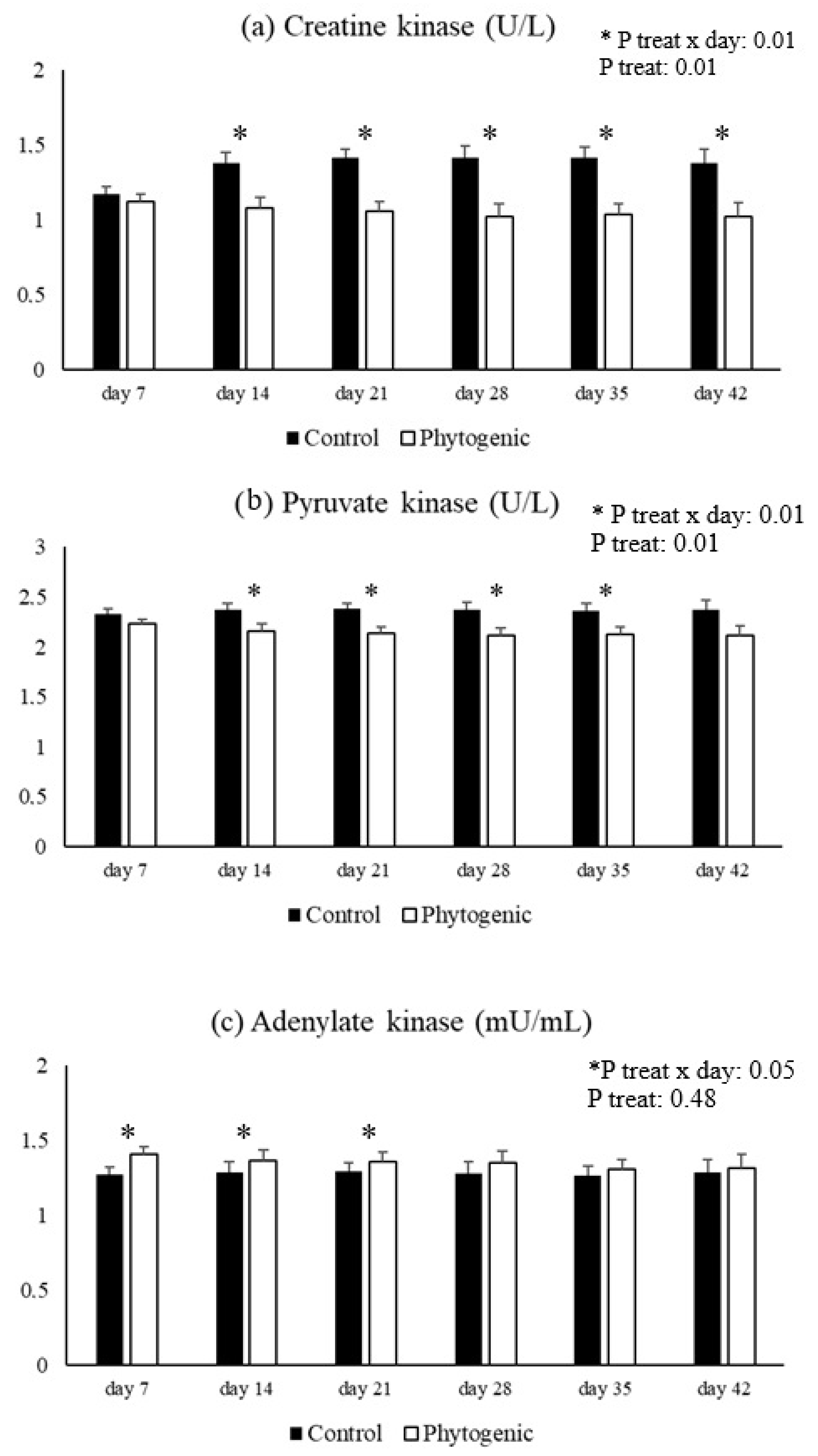
3.4. Energetic Metabolism
3.5. Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benchaar, C.; Calsamiglia, S.; Chaves, A.V.; Fraser, G.R.; Colombatto, D.; McAllister, T.A.; Beauchemin, K.A. A review of plant-derived essential oils in ruminant nutrition and production. Anim. Feed Sci. Technol. 2008, 145, 209–228. [Google Scholar] [CrossRef]
- Calsamiglia, S.; Busquet, M.; Cardozo, P.W.; Castillejos, L.; Ferret, A. Invited review: Essential oils as modifiers of rumen microbial fermentation. J. Dairy Sci. 2007, 90, 2580–2595. [Google Scholar] [CrossRef]
- Oh, J.; Wall, E.H.; Bravo, D.M.; Hristov, A.N. Host-mediated effects of phytonutrients in ruminants: A review. J. Dairy Sci. 2017, 100, 5974–5983. [Google Scholar] [CrossRef] [PubMed]
- Abuelo, A.; Hernández, J.; Benedito, J.L.; Castillo, C. Redox Biology in Transition Periods of Dairy Cattle: Role in the Health of Periparturient and Neonatal Animals. Antioxidants 2019, 8, 20. [Google Scholar] [CrossRef]
- Da Silva, R.B.; Pereira, M.N.; De Araujo, R.C.; De Rezende Silva, W.; Pereira, R.A.N. A blend of essential oils improved feed efficiency and affected ruminal and systemic variables of dairy cows. Transl. Anim. Sci. 2020, 4, 182–193. [Google Scholar] [CrossRef]
- Leal, K.W.; Leal, M.L.R.; Breancini, M.; Signor, M.H.; Vitt, M.G.; Silva, L.E.L.; Wagner, R.; Jung, C.T.K.; Kozloski, G.V.; de Araujo, R.C.; et al. Essential oils and capsaicin in the diet of Jersey cows at early lactation and their positive impact on anti-inflammatory, antioxidant and immunological responses. Trop. Anim. Health Prod. 2024, 56, 247. [Google Scholar] [CrossRef]
- Ai, Y.; Wang, H.; Liu, L.; Qi, Y.; Tang, S.; Tang, J.; Chen, N. Purine and purinergic receptors in health and disease. MedComm 2023, 4, e359. [Google Scholar] [CrossRef]
- Burnstock, G. Purinergic signalling. Br. J. Pharmacol. 2006, 147 (Suppl. S1), S172–S181. [Google Scholar] [CrossRef]
- Beckmann, J.; Lips, K.S. The non-neuronal cholinergic system in health and disease. Pharmacology 2014, 92, 286–302. [Google Scholar] [CrossRef]
- Grando, S.A.; Kawashima, K.; Kirkpatrick, C.J.; Kummer, W.; Wessler, I. Recent progress in revealing the biological and medical significance of the non-neuronal cholinergic system. Int. Immunopharmacol. 2015, 29, 1–7. [Google Scholar] [CrossRef]
- Kawashima, K.; Fujii, T. Basic and clinical aspects of non-neuronal acetylcholine: Overview of non-neuronal cholinergic systems and their biological. J. Pharmacol. Sci. 2008, 106, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H.; Botchkina, G.I.; Watkins, L.R.; Wang, H.; Abumrad, N.; Eaton, J.W.; Tracey, K.J. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature 2000, 405, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, V.A.; Wang, H.; Czura, C.; Friedman, S.G.; Tracey, K.J. The cholinergic anti-inflammatory pathway: A missing link in neuroimmunomodulation. Mol. Med. 2003, 9, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Braun, H.S.; Schrapers, K.T.; Mahlkow-Nerge, K.; Stumpff, F.; Rosendahl, J. Dietary supplementation of essential oils in dairy cows: Evidence for stimulatory effects on nutrient absorption. Animal 2019, 13, 518–523. [Google Scholar] [CrossRef]
- NRC (National Research Council). Nutrient Requirements of Dairy Cattle, 7th ed.; National Academy Press: Washington, DC, USA, 2001. [Google Scholar]
- Jentzsch, A.M.; Bachmann, H.; Fürst, P.; Biesalski, H.K. Improved analysis of malondialdehyde in human body fluids. Free Radic. Biol. Med. 1996, 20, 251–256. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Ali, S.F.; LeBel, C.P.; Bondy, S.C. Reactive oxygen species formation as a biomarker of methylmercury and trimethyltin neurotoxicity. Neurotoxicology 1992, 13, 637–648. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 5th ed.; Acta Crystallographica Section D: Structural Biology; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Maltez, L.C.; Barbas, L.A.L.; Nitz, L.F.; Pellegrin, L.; Okamoto, M.H.; Sampaio, L.A.; Monserrat, J.M.; Garcia, L. Oxidative stress and antioxidant responses in juvenile Brazilian flounder Paralichthys orbignyanus exposed to sublethal levels of nitrite. Fish Physiol. Biochem. 2018, 44, 1349–1362. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar]
- Bøyum, A. Isolation of lymphocytes, granulocytes and macrophages. Scand. J. Immunol. 1976, 5, 9–15. [Google Scholar]
- Pilla, C.; Emanuelli, T.; Frassetto, S.S.; Battastini, A.M.O.; Dias, R.D.; Sarkis, J.J.F. ATP diphosphohydrolase activity (apyrase, EC 3.6.1.5) in human blood platelets. Platelets 1996, 7, 225–230. [Google Scholar] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Giusti, G.; Galanti, B. 3.4. 2 Colorimetric Method. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Verlag Chemie: Weinheim, Germany, 1984; p. 315. [Google Scholar]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Lunkes, G.I.; Lunkes, D.; Stefanello, F.; Morsch, A.; Morsch, V.M.; Mazzanti, C.M.; Schetinger, M.R.C. Enzymes that hydrolyze adenine nucleotides in diabetes and associated pathologies. Thromb. Res. 2003, 109, 189–194. [Google Scholar] [CrossRef]
- Hughes, B.P. A method for the estimation of serum creatine kinase and its use in comparing creatine kinase and aldolase activity in normal and pathological sera. Clin. Chim. Acta 1962, 7, 597–603. [Google Scholar] [CrossRef]
- Dzeja, P.P.; Vitkevicius, K.T.; Redfield, M.M.; Burnett, J.C.; Terzic, A. Adenylate kinase–catalyzed phosphotransfer in the myocardium. Circ. Res. 1999, 84, 1137–1143. [Google Scholar] [CrossRef]
- Leong, S.F.; Lai, J.C.K.; Lim, L.; Clark, J.B. Energy-metabolising enzymes in brain regions of adult and aging rats. J. Neurochem. 1981, 37, 1548–1556. [Google Scholar] [CrossRef]
- Silva, D.J.; Queiroz, A.C. Análise de Alimentos: Métodos Químicos e Biológicos, 3rd ed.; UFV: Viçosa, Brazil, 2006; pp. 1–235. [Google Scholar]
- Komarek, A.R.; Manson, H.; Thiex, N. Crude Fiber Determinations Using the ANKOM System; Technology Corporation, Publication: Macedon, NY, USA, 1996; Volume 102. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Roland, L.; Drillich, M.; Iwersen, M. Hematology as a diagnostic tool in bovine medicine. J. Vet. Diagn. Investig. 2014, 26, 592–598. [Google Scholar] [CrossRef]
- González, F.H.D.; Silva, S.C. Introdução à Bioquímica Clínica Veterinária; UFRGS: Porto Alegre, Brazil, 2017. [Google Scholar]
- Kaneko, J.J.; Harvey, J.W.; Bruss, M.L. Clinical Biochemistry of Domestic Animals; Academic Press: San Diego, CA, USA, 2008. [Google Scholar] [CrossRef]
- Favaretto, J.A.; Alba, D.F.; Marchiori, M.S.; Marcon, H.J.; Souza, C.F.; Baldissera, M.D.; Bianchi, A.E.; Zanluchi, M.; Klein, B.; Wagner, R.; et al. Supplementation with a blend based on micro-encapsulated carvacrol, thymol, and cinnamaldehyde in lambs feed inhibits immune cells and improves growth performance. Livest. Sci. 2020, 240, 104144. [Google Scholar] [CrossRef]
- Wallace, R.J. Antimicrobial properties of plant secondary metabolites. Proc. Nutr. Soc. 2004, 63, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Demirbilek, S.; Ersoy, M.O.; Demirbilek, S.; Karaman, A.; Gürbüz, N.; Bayraktar, N.; Bayraktar, M. Small-dose capsaicin reduces systemic inflammatory responses in septic rats. Anesth. Analg. 2004, 99, 1501–1507. [Google Scholar] [CrossRef]
- Lima, M.D.S.; Quintans-Júnior, L.J.; De Santana, W.A.; Martins Kaneto, C.; Pereira Soares, M.B.; Villarreal, C.F. Anti-inflammatory effects of carvacrol: Evidence for a key role of interleukin-10. Eur. J. Pharmacol. 2013, 699, 112–117. [Google Scholar] [CrossRef]
- Pandey, V.K.; Srivastava, S.; Ashish; Dash, K.K.; Singh, R.; Dar, A.H.; Singh, T.; Farooqui, A.; Shaikh, A.M.; Kovacs, B. Bioactive properties of clove (Syzygium aromaticum) essential oil nanoemulsion: A comprehensive review. Heliyon 2024, 10, e22437. [Google Scholar] [CrossRef]
- Schink, A.; Naumoska, K.; Kitanovski, Z.; Kampf, C.J.; Fröhlich-Nowoisky, J.; Thines, E.; Pöschl, U.; Schuppan, D.; Lucas, K. Anti-inflammatory effects of cinnamon extract and identification of active compounds influencing the TLR2 and TLR4 signaling pathways. Food Funct. 2018, 9, 5950–5964. [Google Scholar] [CrossRef]
- Brunetto, A.L.; Giacomelli, C.M.; Favero, J.F.; Bissacotti, B.F.; Copeti, P.M.; Morsch, V.M.; Oliveira, F.d.C.d.; Wagner, R.; Alves, R.; Pereira, W.A.; et al. Phytogenic blend in the diet of growing Holstein steers: Effects on performance, digestibility, rumen volatile fatty acid profile, and immune and antioxidant responses. Anim. Feed. Sci. Technol. 2023, 297, 115595. [Google Scholar] [CrossRef]
- Fachini-Queiroz, F.C.; Kummer, R.; Estevão-Silva, C.F.; Carvalho, M.D.D.B.; Cunha, J.M.; Grespan, R.; Bersani-Amado, C.A.; Cuman, R.K.N. Effects of thymol and carvacrol, constituents of Thymus vulgaris L. essential oil, on the inflammatory response. Evid.-Based Complement. Altern. Med. 2012, 2012, 657026. [Google Scholar] [CrossRef]
- Miettinen, P.V.A.; Juvonen, R.O. Diurnal variations of serum and milk urea levels in dairy cows. Acta Agric. Scand. 1990, 40, 289–296. [Google Scholar] [CrossRef]
- Otto-Ślusarczyk, D.; Graboń, W.; Mielczarek-Puta, M. Aspartate aminotransferase—Key enzyme in the human systemic metabolism. Adv. Hyg. Exp. Med. 2016, 70, 219–230. [Google Scholar] [CrossRef]
- Aristatile, B.; Al-Numair, K.S.; Veeramani, C.; Pugalendi, V. Effect of carvacrol on hepatic marker enzymes and antioxidant status in D-galactosamine-induced hepatotoxicity in rats. Fundam. Clin. Pharmacol. 2009, 23, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Siddiqi, N.J.; Alrashood, S.T.; Khan, H.A.; Dubey, A.; Sharma, B. Protective effect of eugenol on hepatic inflammation and oxidative stress induced by cadmium in male rats. Biomed. Pharmacother. 2021, 139, 111588. [Google Scholar] [CrossRef]
- Eidi, A.; Mortazavi, P.; Bazargan, M.; Zaringhalam, J. Hepatoprotective activity of cinnamon ethanolic extract against CCI4-induced liver injury in rats. Excli J. 2012, 11, 495. [Google Scholar] [CrossRef]
- Ghorbanpour, A.; Salari, S.; Baluchnejadmojarad, T.; Roghani, M. Capsaicin protects against septic acute liver injury by attenuation of apoptosis and mitochondrial dysfunction. Heliyon 2023, 9, e14205. [Google Scholar] [CrossRef]
- Amorati, R.; Foti, M.C.; Valgimigli, L. Antioxidant activity of essential oils. J. Agric. Food Chem. 2013, 61, 10835–10847. [Google Scholar] [CrossRef]
- Bhavaniramya, S.; Vishnupriya, S.; Al-Aboody, M.S.; Vijayakumar, R.; Baskaran, D. Role of essential oils in food safety: Antimicrobial and antioxidant applications. Grain Oil Sci. Technol. 2019, 2, 49–55. [Google Scholar] [CrossRef]
- Dickinson, D.A.; Forman, H.J. Cellular glutathione and thiols metabolism. Biochem. Pharmacol. 2002, 64, 1019–1026. [Google Scholar] [CrossRef]
- Abdel-Salam, O.M.E.; Abdel-Rahman, R.F.; Sleem, A.A.; Farrag, A.R. Modulation of lipopolysaccharide-induced oxidative stress by capsaicin. Inflammopharmacology 2012, 20, 207–217. [Google Scholar] [CrossRef]
- Wang, H.; Yu, M.; Ochani, M.; Amella, C.A.; Tanovic, M.; Susarla, S.; Li, J.H.; Wang, H.; Yang, N.; Ulloa, L.; et al. Nicotinic acetylcholine receptor α7 subunit is an essential regulator of inflammation. Nature 2003, 421, 384–388. [Google Scholar] [CrossRef]
- Mazzanti, C.M.; dos Anjos Lopes, S.T.; Da Silva, A.S.; França, R.T.; Costa, M.M.; Paim, C.B.; Paim, F.C.; Schmatz, R.; Jaques, J.A.; Schetinger, M.R.; et al. Cholinesterase activity in serum, whole blood and lymphocytes of dogs experimentally infected with Rangelia Vitalii. Acta Sci. Vet. 2013, 41, 1–7. Available online: http://www.ufrgs.br/actavet/41/PUB%201155.pdf (accessed on 2 November 2024).
- Kawashima, K.; Fujii, T. The lymphocytic cholinergic system and its contribution to the regulation of immune activity. Life Sci. 2003, 74, 675–696. [Google Scholar] [CrossRef] [PubMed]
- Kimura, R.; Ushiyama, N.; Fujii, T.; Kawashima, K. Nicotine-induced Ca2+ signaling and down-regulation of nicotinic acetylcholine receptor subunit expression in the CEM human leukemic T-cell line. Life Sci. 2003, 72, 2155–2158. [Google Scholar] [CrossRef] [PubMed]
- Tayebati, S.K.; El-Assouad, D.; Ricci, A.; Amenta, F. Immunochemical and immunocytochemical characterization of cholinergic markers in human peripheral blood lymphocytes. J. Neuroimmunol. 2002, 132, 147–155. [Google Scholar] [CrossRef]
- Pavlov, V.A.; Parrish, W.R.; Rosas-Ballina, M.; Ochani, M.; Puerta, M.; Ochani, K.; Chavan, S.; Al-Abed, Y.; Tracey, K.J. Brain acetylcholinesterase activity controls systemic cytokine levels through the cholinergic anti-inflammatory pathway. Brain Behav Immun. 2009, 23, 41–45. [Google Scholar] [CrossRef]
- Tracey, K.J. Physiology and immunology of the cholinergic antiinflammatory pathway. J. Clin. Investig. 2007, 117, 289–296. [Google Scholar] [CrossRef]
- Da Silva, A.S.; Monteiro, S.G.; Gonçalves, J.F.; Spanevello, R.; Schmatz, R.; Oliveira, C.B.; Costa, M.M.; França, R.T.; Jaques, J.A.; Schetinger, M.R.C.; et al. Trypanosoma evansi: Immune response and acetylcholinesterase activity in lymphocytes from infected rats. Exp. Parasitol. 2011, 127, 475–480. [Google Scholar] [CrossRef]
- Tonin, A.A.; Da Silva, A.S.; Schafer, A.S.; Aires, A.R.; Oliveira, C.B.; Zanini, D.; Schetinger, M.R.; Morsch, V.M.; Lopes, S.T.; Monteiro, S.G.; et al. Influence of experimental infection by Haemonchus contortus on acetylcholinesterase activity in lymphocytes of lambs. Exp. Parasitol. 2014, 139, 19–23. [Google Scholar] [CrossRef]
- Burnstock, G. Pathophysiology and therapeutic potential of purinergic signaling. Pharmacol Rev. 2006, 58, 58–86. [Google Scholar] [CrossRef]
- Zimmermann, H. Ectonucleoside triphosphate diphosphohydrolases and ecto-5′-nucleotidase in purinergic signaling: How the field developed and where we are now. Purinergic Signal. 2021, 17, 117–125. [Google Scholar] [CrossRef]
- Mullen, N.J.; Singh, P.K. Nucleotide metabolism: A pan-cancer metabolic dependency. Nat. Rev. Cancer 2023, 23, 275–294. [Google Scholar] [CrossRef]
- Forrester, T.; Williams, C.A. Release of adenosine triphosphate from isolated adult heart cells in response to hypoxia. J. Physiol. 1977, 268, 371. [Google Scholar] [CrossRef] [PubMed]
- Linden, J.; Koch-Nolte, F.; Dahl, G. Purine release, metabolism, and signaling in the inflammatory response. Annu. Rev. Immunol. 2019, 7, 325–347. [Google Scholar] [CrossRef] [PubMed]
- da Rocha Lapa, F.; da Silva, M.D.; de Almeida Cabrini, D.; Santos, A.R.S. Anti-inflammatory effects of purine nucleosides, adenosine and inosine, in a mouse model of pleurisy: Evidence for the role of adenosine A2 receptors. Purinergic Signal. 2012, 8, 693–704. [Google Scholar] [CrossRef]
- Welihinda, A.A.; Kaur, M.; Raveendran, K.S.; Amento, E.P. Enhancement of inosine-mediated A2AR signaling through positive allosteric modulation. Cell Signal. 2018, 42, 227–235. [Google Scholar] [CrossRef]
- Rocha, C.A.; Félix, L.M.; Monteiro, S.M.; Venâncio, C. Antinociceptive Analysis of Natural Monoterpenes Eugenol, Menthol, Carvacrol and Thymol in a Zebrafish Larval Model. Pharmaceuticals 2024, 17, 457. [Google Scholar] [CrossRef]
- Dzeja, P.P.; Terzic, A. Phosphotransfer networks and cellular energetics. J. Exp. Biol. 2003, 206, 2039–2047. [Google Scholar] [CrossRef]
- Schlattner, U.; Tokarska-Schlattner, M.; Wallimann, T. Mitochondrial creatine kinase in human health and disease. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2006, 1762, 164–180. [Google Scholar] [CrossRef]
- Dzeja, P.; Terzic, A. Adenylate kinase and AMP signaling networks: Metabolic monitoring, signal communication and body energy sensing. Int. J. Mol. Sci. 2009, 10, 1729–1772. [Google Scholar] [CrossRef]
- Baldissera, M.D.; Souza, C.F.; Descovi, S.N.; Verdi, C.M.; Zeppenfeld, C.C.; da Silva, A.S.; Santos, R.C.; Baldisserotto, B. Grape pomace flour ameliorates Pseudomonas aeruginosa-induced bioenergetic dysfunction in gills of grass carp. Aquaculture 2019, 506, 359–366. [Google Scholar] [CrossRef]
- Dzeja, P.P.; Zeleznikar, R.J.; Goldberg, N.D. Suppression of creatine kinase-catalyzed phosphotransfer results in increased phosphoryl transfer by adenylate kinase in intact skeletal muscle. J. Biol. Chem. 1996, 271, 12847–12851. [Google Scholar] [CrossRef]
- Cardoso, G.M.F.; Pletsch, J.T.; Parmeggiani, B.; Grings, M.; Glanzel, N.M.; Bobermin, L.D.; Amaral, A.U.; Wajner, M.; Leipnitz, G. Bioenergetics dysfunction, mitochondrial permeability transition pore opening and lipid peroxidation induced by hydrogen sulfide as relevant pathomechanisms underlying the neurological dysfunction characteristic of ethylmalonic encephalopathy. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2017, 1863, 2192–2201. [Google Scholar] [CrossRef] [PubMed]
- Alvorcem, L.d.M.; Britto, R.; Parmeggiani, B.; Glanzel, N.M.; da Rosa-Junior, N.T.; Cecatto, C.; Bobermin, L.D.; Amaral, A.U.; Wajner, M.; Leipnitz, G. Evidence that thiol group modification and reactive oxygen species are involved in hydrogen sulfide-induced mitochondrial permeability transition pore opening in rat cerebellum. Mitochondrion 2019, 47, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Grummer, R.R.; Mashek, D.G.; Hayirli, A. Dry matter intake and energy balance in the transition period. Vet. Clin. North Am.—Food Anim. Pract. 2004, 20, 447–470. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Variables, % | TMR: Supplied in Individual Feeders | * TMR: Provided in Intergado® | Pelleted Feed for Robot |
|---|---|---|---|
| Dry matter | 38.2 | 33.7 | 86.7 |
| Crude protein | 19.3 | 14.4 | 20.8 |
| Ether extract | 3.01 | 2.67 | 2.77 |
| Ash | 7.08 | 6.62 | 5.93 |
| Neutral detergent fiber (NDF) | 42.2 | 51.2 | 27.6 |
| Acid detergent fiber (ADF) | 16.6 | 22.7 | 8.98 |
| Gross energy (Kcal) | 271 | 282 | 379 |
| Variables | Control | Phytogenic | SEM | P: Treat | P: Treat × Day |
|---|---|---|---|---|---|
| Hemogram | |||||
| Leukocytes (×103 µL) | 0.58 | 0.05 | |||
| d1 | 5.62 | 5.58 | 0.15 | ||
| d7 | 5.89 a | 5.03 b | 0.16 | ||
| d14 | 5.76 a | 4.87 b | 0.15 | ||
| d21 | 5.56 | 5.09 | 0.15 | ||
| d28 | 5.62 a | 4.71 b | 0.15 | ||
| d35 | 5.61 | 5.21 | 0.14 | ||
| d42 | 5.48 | 5.33 | 0.14 | ||
| Lymphocytes (×103 µL) | 0.41 | 0.03 | |||
| d1 | 3.15 | 3.01 | 0.11 | ||
| d7 | 3.25 a | 2.67 b | 0.11 | ||
| d14 | 3.43 a | 2.83 b | 0.10 | ||
| d21 | 3.25 | 3.91 | 0.12 | ||
| d28 | 3.37 | 3.89 | 0.12 | ||
| d35 | 3.35 a | 2.74 b | 0.10 | ||
| d42 | 3.12 | 3.05 | 0.11 | ||
| Granulocytes (×103 µL) | 1.34 | 1.40 | 0.13 | 0.84 | 0.91 |
| Monocytes (×103 µL) | 0.94 | 0.87 | 0.26 | 0.82 | 0.63 |
| Erythrocytes (×106 µL) | 5.32 | 5.33 | 0.03 | 0.95 | 0.97 |
| Hemoglobin | 10.0 | 9.95 | 0.09 | 0.93 | 0.95 |
| Hematocrit, % | 26.9 | 27.2 | 0.45 | 0.92 | 0.87 |
| Platelets (×103 µL) | 302 | 264 | 14.6 | 0.32 | 0.15 |
| Serum biochemistry | |||||
| Total protein (g/dL) | 7.57 | 7.62 | 0.11 | 0.76 | 0.81 |
| Globulin (g/dL) | 4.05 b | 4.30 a | 0.10 | 0.07 | 0.11 |
| Albumin (g/dL) | 3.51 | 3.32 | 0.08 | 0.65 | 0.52 |
| Glucose (mg/dL) | 43.6 | 46.7 | 0.54 | 0.34 | 0.28 |
| Cholesterol (mg/dL) | 160 | 147 | 4.89 | 0.12 | 0.21 |
| Triglycerides (mg/dL) | 8.80 | 10.1 | 0.59 | 0.64 | 0.72 |
| Urea (mg/dL) | 0.16 | 0.05 | |||
| d1 | 35.6 | 39.7 | 2.13 | ||
| d7 | 36.7 | 37.4 | 2.09 | ||
| d14 | 45.1 | 47.3 | 2.06 | ||
| d21 | 48.6 | 44.3 | 2.07 | ||
| d28 | 51.8 a | 41.7 b | 2.01 | ||
| d35 | 48.3 | 42.6 | 2.04 | ||
| d42 | 42.6 | 38.4 | 2.01 | ||
| GGT (U/L) | 14.6 | 17.2 | 1.25 | 0.41 | 0.57 |
| AST (U/L) | 0.02 | 0.01 | |||
| d1 | 91.8 | 89.7 | 4.65 | ||
| d7 | 88.3 | 90.3 | 5.03 | ||
| d14 | 96.2 | 84.7 | 5.01 | ||
| d21 | 92.1 | 76.1 | 5.21 | ||
| d28 | 123 a | 91.4 b | 6.14 | ||
| d35 | 93.7 a | 71.2 b | 5.85 | ||
| d42 | 80.3 | 67.3 | 5.12 | ||
| Cholinesterase (×10 U/L) | 0.67 | 0.01 | |||
| d1 | 114 | 118 | 3.52 | ||
| d7 | 121 | 124 | 3.45 | ||
| d14 | 132 | 128 | 3.51 | ||
| d21 | 117 | 102 | 3.42 | ||
| d28 | 115 a | 84.9 b | 3.49 | ||
| d35 | 138 a | 122 b | 3.45 | ||
| d42 | 113 b | 137 a | 3.52 | ||
| BHB (mmol/L) | 1.18 | 1.15 | 0.03 | 0.95 | 0.93 |
| Variables | Control | Phytogenic | SEM 2 | Treat 1 | Treat × Day 1 |
|---|---|---|---|---|---|
| CytokinesInterleukin 1β (pg/mL) | 26.9 a | 17.8 b | 1.68 | 0.01 | 0.01 |
| d14 | 28.3 a | 19.4 b | 1.71 | ||
| d28 | 27.0 a | 16.9 b | 1.70 | ||
| d42 | 25.4 a | 17.3 b | 1.70 | ||
| Interleukin 6 (pg/mL) | 17.5 a | 10.2 b | 0.91 | 0.02 | 0.01 |
| d14 | 18.2 a | 10.2 b | 0.90 | ||
| d28 | 17.9 a | 9.98 b | 0.93 | ||
| d42 | 16.5 a | 10.4 b | 0.95 | ||
| Interleukin 10 (pg/mL) | 36.5 b | 48.9 a | 3.31 | 0.01 | 0.01 |
| d14 | 35.7 b | 41.6 a | 3.34 | ||
| d28 | 36.4 b | 48.9 a | 3.30 | ||
| d42 | 37.6 b | 56.4 a | 3.32 | ||
| Oxidative statusTBARS (nmol/mL) | 12.4 a | 9.74 b | 1.25 | 0.05 | 0.12 |
| ROS (% of fluorescence intensity) | 0.04 | 0.01 | |||
| d14 | 89.1 | 87.2 | 2.35 | ||
| d28 | 83.4 a | 69.7 b | 6.04 | ||
| d42 | 81.0 a | 62.2 b | 4.50 | ||
| PSH (µmol/L) | 3.52 | 3.68 | 0.56 | 0.85 | 0.72 |
| NPSH (µmol/L) | 0.82 b | 0.91 a | 0.06 | 0.01 | 0.01 |
| d14 | 0.85 b | 0.97 a | 0.05 | ||
| d28 | 0.82 b | 0.99 a | 0.05 | ||
| d42 | 0.86 b | 1.05 a | 0.06 | ||
| GST (U GST/mg protein) | 0.01 | 0.01 | |||
| d14 | 249 | 247 | 7.44 | ||
| d28 | 241 b | 222 a | 7.89 | ||
| d42 | 210 b | 246 a | 9.85 |
| Variables | Control | Phytogenic | SEM | P: Treat 1 | P: Treat × Day 1 |
|---|---|---|---|---|---|
| Milk production (kg/d) | 15.6 | 15.2 | 0.30 | 0.92 | 0.95 |
| FCM-4% (kg/day) 2 | 17.3 | 16.2 | 0.26 | 0.35 | NE |
| ECM (lbs) | 212 | 199 | 3.29 | 0.12 | NE |
| Feed intake, kg DM | 14.0 | 13.8 | 0.18 | 0.94 | 0.89 |
| Feed efficiency | 1.11 | 1.10 | 0.05 | 0.97 | NE |
| Milk fat, % | 4.75 | 4.45 | 0.06 | 0.45 | 0.29 |
| Milk protein, % | 3.89 | 3.67 | 0.04 | 0.52 | 0.44 |
| Milk lactose, % | 4.64 | 4.62 | 0.02 | 0.97 | 0.94 |
| Milk total solids, % | 14.3 | 13.7 | 0.36 | 0.50 | 0.43 |
| SCC (×103/mL) | 169 | 229 | 20.4 | 0.62 | 0.49 |
| Body score | 3.37 | 3.41 | 0.03 | 0.38 | 0.73 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leal, K.W.; do Rego Leal, M.L.; Klein, G.S.; Brunetto, A.L.R.; Deolindo, G.L.; Justen, C.E.; Baldissera, M.D.; Santos, T.L.; Zanini, D.; de Araujo, R.C.; et al. Role of Microencapsulated Essential Oil and Pepper Resin in the Diet of Cows in the Third Lactation Phase on Immunological Pathways. Vet. Sci. 2025, 12, 344. https://doi.org/10.3390/vetsci12040344
Leal KW, do Rego Leal ML, Klein GS, Brunetto ALR, Deolindo GL, Justen CE, Baldissera MD, Santos TL, Zanini D, de Araujo RC, et al. Role of Microencapsulated Essential Oil and Pepper Resin in the Diet of Cows in the Third Lactation Phase on Immunological Pathways. Veterinary Sciences. 2025; 12(4):344. https://doi.org/10.3390/vetsci12040344
Chicago/Turabian StyleLeal, Karoline Wagner, Marta Lizandra do Rego Leal, Gabriel S. Klein, Andrei Lucas R. Brunetto, Guilherme Luiz Deolindo, Camila Eduarda Justen, Matheus Dellaméa Baldissera, Tainara L. Santos, Daniela Zanini, Rafael C. de Araujo, and et al. 2025. "Role of Microencapsulated Essential Oil and Pepper Resin in the Diet of Cows in the Third Lactation Phase on Immunological Pathways" Veterinary Sciences 12, no. 4: 344. https://doi.org/10.3390/vetsci12040344
APA StyleLeal, K. W., do Rego Leal, M. L., Klein, G. S., Brunetto, A. L. R., Deolindo, G. L., Justen, C. E., Baldissera, M. D., Santos, T. L., Zanini, D., de Araujo, R. C., & da Silva, A. S. (2025). Role of Microencapsulated Essential Oil and Pepper Resin in the Diet of Cows in the Third Lactation Phase on Immunological Pathways. Veterinary Sciences, 12(4), 344. https://doi.org/10.3390/vetsci12040344