


Use of Monoglycerides and Diglycerides to Mitigate Poultry Production Losses: A Review

Abstract
Simple Summary
Abstract
1. Introduction
2. Nutrient Utilization, Metabolism, Growth, and Carcass Composition
3. Antimicrobial Activity of Monoglycerides and Diglycerides
4. Antiviral Activity
5. Effects on Intestinal Morphology, Gut Immunity, and the Microbiome
5.1. Bacterial Patterns with MDG
5.2. Gut Integrity and Function with MDG
5.3. Gut Immunity
5.4. Free Fatty Acid Receptors (FFAR)
6. Practical Applications
7. Summary
- Most in vivo responses are observed when the diet contained between 0.05% and 0.5% MDG and inclusion rates as high as 1% showed no adverse effects;
- Nutrient utilization, carcass composition, and serum biochemistry results tend to point toward glyceride-dependent reductions in fat deposition with concurrent increases in muscle deposition;
- Antimicrobial activity is seen against many of the problematic organisms found in poultry production and meat processing including C. perfringens, E. coli, Salmonella spp., Campylobacter spp., Mycoplasma spp., and some enveloped viruses as illustrated in Table 2;
- There tends to be an increase in the abundance of specific beneficial bacteria (i.e., SCFA producers) and a general increase in intestinal community diversity;
- Increases in overall enteric health and function were commonly reported through a variety of mechanisms including improved tight junction integrity, increased villus height, and reduced inflammation.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dohlman, E. USDA Agricultural Projections to 2032. 2023. Available online: https://www.ers.usda.gov/webdocs/outlooks/105853/oce-2023-01.pdf?v=5101.4 (accessed on 12 December 2023).
- FAO. Contribution of Terrestrial Animal Source Food to Healthy Diets for Improved Nutrition and Health Outcomes: An Evidence and Policy Overview on the State of Knowledge and Gaps; FAO: Rome, Italy, 2023; ISBN 978-92-5-137536-5. [Google Scholar]
- Jackman, J.A.; Lavergne, T.A.; Elrod, C.C. Antimicrobial Monoglycerides for Swine and Poultry Applications. Front. Anim. Sci. 2022, 3, 1019320. [Google Scholar] [CrossRef]
- Zhang, Q.; Wu, W.; Zhang, J.; Xia, X. Antimicrobial Lipids in Nano-Carriers for Antibacterial Delivery. J. Drug Target. 2020, 28, 271–281. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jackman, J.A.; Valle-González, E.R.; Cho, N.-J. Antibacterial Free Fatty Acids and Monoglycerides: Biological Activities, Experimental Testing, and Therapeutic Applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef]
- 21 CFR 184.1505—Mono- and Diglycerides. Available online: https://www.ecfr.gov/current/title-21/part-184/section-184.1505 (accessed on 12 December 2023).
- EFSA Panel on Food Additives and Nutrient Sources added to Food (ANS); Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Dusemund, B.; Filipič, M.; Frutos, M.J.; Galtier, P.; Gott, D.; et al. Re-Evaluation of Mono- and Di-Glycerides of Fatty Acids (E 471) as Food Additives. EFSA J. 2017, 15, e05045. [Google Scholar] [CrossRef]
- Singh, I.R.; Pulikkal, A.K. Preparation, Stability and Biological Activity of Essential Oil-Based Nano Emulsions: A Comprehensive Review. OpenNano 2022, 8, 100066. [Google Scholar] [CrossRef]
- Eduardo, M.; Svanberg, U.; Ahrné, L. Effect of Hydrocolloids and Emulsifiers on the Shelf-Life of Composite Cassava-Maize-Wheat Bread after Storage. Food Sci. Nutr. 2016, 4, 636–644. [Google Scholar] [CrossRef]
- Amer, S.A.; A-Nasser, A.; Al-Khalaifah, H.S.; AlSadek, D.M.M.; Abdel Fattah, D.M.; Roushdy, E.M.; Sherief, W.R.I.A.; Farag, M.F.M.; Altohamy, D.E.; Abdel-Wareth, A.A.A.; et al. Effect of Dietary Medium-Chain α-Monoglycerides on the Growth Performance, Intestinal Histomorphology, Amino Acid Digestibility, and Broiler Chickens’ Blood Biochemical Parameters. Animals 2021, 11, 57. [Google Scholar] [CrossRef]
- Bedford, A.; Yu, H.; Hernandez, M.; Squires, E.J.; Leeson, S.; Gong, J. Effects of Fatty Acid Glyceride Product SILOhealth 104 on the Growth Performance and Carcass Composition of Broiler Chickens. Poult. Sci. 2018, 97, 1315–1323. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Ruan, S.; Mo, Q.; Zhao, M.; Wang, J.; Ye, Z.; Chen, L.; Feng, F. Evaluation of Dynamic Effects of Dietary Medium-Chain Monoglycerides on Performance, Intestinal Development and Gut Microbiota of Broilers in Large-Scale Production. Anim. Nutr. 2023, 14, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Osorio, L.-M.; Yepes-Medina, V.; Ballou, A.; Parini, M.; Angel, R. Short and Medium Chain Fatty Acids and Their Derivatives as a Natural Strategy in the Control of Necrotic Enteritis and Microbial Homeostasis in Broiler Chickens. Front. Vet. Sci. 2021, 8, 773372. [Google Scholar] [CrossRef] [PubMed]
- Thormar, H.; Hilmarsson, H.; Bergsson, G. Stable Concentrated Emulsions of the 1-Monoglyceride of Capric Acid (Monocaprin) with Microbicidal Activities against the Food-Borne Bacteria Campylobacter jejuni, Salmonella spp., and Escherichia coli. Appl. Environ. Microbiol. 2006, 72, 522–526. [Google Scholar] [CrossRef] [PubMed]
- Dobson, D.E.; Kambe, A.; Block, E.; Dion, T.; Lu, H.; Castellot, J.J.; Spiegelman, B.M. 1-Butyryl-Glycerol: A Novel Angiogenesis Factor Secreted by Differentiating Adipocytes. Cell 1990, 61, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.U.; Naz, S.; Raziq, F.; Qudratullah, Q.; Khan, N.A.; Laudadio, V.; Tufarelli, V.; Ragni, M. Prospects of Organic Acids as Safe Alternative to Antibiotics in Broiler Chickens Diet. Environ. Sci. Pollut. Res. 2022, 29, 32594–32604. [Google Scholar] [CrossRef] [PubMed]
- Haetinger, V.S.; Dalmoro, Y.K.; Godoy, G.L.; Lang, M.B.; de Souza, O.F.; Aristimunha, P.; Stefanello, C. Optimizing Cost, Growth Performance, and Nutrient Absorption with a Bio-Emulsifier Based on Lysophospholipids for Broiler Chickens. Poult. Sci. 2021, 100, 101025. [Google Scholar] [CrossRef]
- Roy, A.; Haldar, S.; Mondal, S.; Ghosh, T.K. Effects of Supplemental Exogenous Emulsifier on Performance, Nutrient Metabolism, and Serum Lipid Profile in Broiler Chickens. Vet. Med. Int. 2010, 2010, 262604. [Google Scholar] [CrossRef]
- Siyal, F.A.; Babazadeh, D.; Wang, C.; Arain, M.A.; Saeed, M.; Ayasan, T.; Zhang, L.; Wang, T. Emulsifiers in the Poultry Industry. Worlds Poult. Sci. J. 2017, 73, 611–620. [Google Scholar] [CrossRef]
- Hu, X.Q.; Wang, W.B.; Liu, L.; Wang, C.; Feng, W.; Luo, Q.P.; Han, R.; Wang, X.D. Effects of Fat Type and Emulsifier in Feed on Growth Performance, Slaughter Traits, and Lipid Metabolism of Cherry Valley Ducks. Poult. Sci. 2019, 98, 5759–5766. [Google Scholar] [CrossRef]
- Feye, K.M.; Dittoe, D.K.; Jendza, J.A.; Caldas-Cueva, J.P.; Mallmann, B.A.; Booher, B.; Tellez-Isaias, G.; Owens, C.M.; Kidd, M.T.; Ricke, S.C. A Comparison of Formic Acid or Monoglycerides to Formaldehyde on Production Efficiency, Nutrient Absorption, and Meat Yield and Quality of Cobb 700 Broilers. Poult. Sci. 2021, 100, 101476. [Google Scholar] [CrossRef]
- Liu, T.; Mo, Q.; Wei, J.; Zhao, M.; Tang, J.; Feng, F. Mass Spectrometry-Based Metabolomics to Reveal Chicken Meat Improvements by Medium-Chain Monoglycerides Supplementation: Taste, Fresh Meat Quality, and Composition. Food Chem. 2021, 365, 130303. [Google Scholar] [CrossRef]
- Leeson, S.; Namkung, H.; Antongiovanni, M.; Lee, E.H. Effect of Butyric Acid on the Performance and Carcass Yield of Broiler Chickens. Poult. Sci. 2005, 84, 1418–1422. [Google Scholar] [CrossRef]
- Antongiovanni, M.; Buccioni, A.; Petacchi, F.; Leeson, S.; Minieri, S.; Martini, A.; Cecchi, R. Butyric Acid Glycerides in the Diet of Broiler Chickens: Effects on Gut Histology and Carcass Composition. Ital. J. Anim. Sci. 2007, 6, 19–25. [Google Scholar] [CrossRef]
- Yin, F.; Yu, H.; Lepp, D.; Shi, X.; Yang, X.; Hu, J.; Leeson, S.; Yang, C.; Nie, S.; Hou, Y.; et al. Transcriptome Analysis Reveals Regulation of Gene Expression for Lipid Catabolism in Young Broilers by Butyrate Glycerides. PloS ONE 2016, 11, e0160751. [Google Scholar] [CrossRef]
- Bedford, A.; Yu, H.; Squires, E.J.; Leeson, S.; Gong, J. Effects of Supplementation Level and Feeding Schedule of Butyrate Glycerides on the Growth Performance and Carcass Composition of Broiler Chickens. Poult. Sci. 2017, 96, 3221–3228. [Google Scholar] [CrossRef] [PubMed]
- Jackman, J.A.; Yoon, B.K.; Li, D.; Cho, N.-J. Nanotechnology Formulations for Antibacterial Free Fatty Acids and Monoglycerides. Molecules 2016, 21, 305. [Google Scholar] [CrossRef] [PubMed]
- Batovska, D.I.; Todorova, I.T.; Tsvetkova, I.V.; Najdenski, H.M. Antibacterial Study of the Medium Chain Fatty Acids and Their 1-Monoglycerides: Individual Effects and Synergistic Relationships. Pol. J. Microbiol. 2009, 58, 43–47. [Google Scholar] [PubMed]
- Davies, J.; Davies, D. Origins and Evolution of Antibiotic Resistance. Microbiol. Mol. Biol. Rev. MMBR 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Kabara, J.J.; Vrable, R.; Lie Ken Jie, M.S.F. Antimicrobial Lipids: Natural and Synthetic Fatty Acids and Monoglycerides. Lipids 1977, 12, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Hussain, M.M. Intestinal Lipid Absorption. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E1183–E1194. [Google Scholar] [CrossRef] [PubMed]
- Projan, S.J.; Brown-Skrobot, S.; Schlievert, P.M.; Vandenesch, F.; Novick, R.P. Glycerol Monolaurate Inhibits the Production of Beta-Lactamase, Toxic Shock Toxin-1, and Other Staphylococcal Exoproteins by Interfering with Signal Transduction. J. Bacteriol. 1994, 176, 4204–4209. [Google Scholar] [CrossRef] [PubMed]
- Schlievert, P.M.; Peterson, M.L. Glycerol Monolaurate Antibacterial Activity in Broth and Biofilm Cultures. PloS ONE 2012, 7, e40350. [Google Scholar] [CrossRef]
- Schlievert, P.M.; Deringer, J.R.; Kim, M.H.; Projan, S.J.; Novick, R.P. Effect of Glycerol Monolaurate on Bacterial Growth and Toxin Production. Antimicrob. Agents Chemother. 1992, 36, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Fierer, J.; Guiney, D.G. Diverse Virulence Traits Underlying Different Clinical Outcomes of Salmonella Infection. J. Clin. Investig. 2001, 107, 775–780. [Google Scholar] [CrossRef]
- Vetter, S.M.; Schlievert, P.M. Glycerol Monolaurate Inhibits Virulence Factor Production in Bacillus Anthracis. Antimicrob. Agents Chemother. 2005, 49, 1302–1305. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, R.; Zhang, G.; Chen, F.; Xu, B. In Vitro Antibacterial Activities and Mechanisms of Action of Fatty Acid Monoglycerides Against Four Foodborne Bacteria. J. Food Prot. 2020, 83, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Zhao, J.; Zeng, Z.; Wan, D.; Yu, P.; Cheng, D.; Gong, D.; Deng, S. Antibacterial Activity and Membrane-Disrupting Mechanism of Monocaprin against Escherichia Coli and Its Application in Apple and Carrot Juices. LWT 2020, 131, 109794. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jackman, J.A.; Kim, M.C.; Sut, T.N.; Cho, N.-J. Correlating Membrane Morphological Responses with Micellar Aggregation Behavior of Capric Acid and Monocaprin. Langmuir 2017, 33, 2750–2759. [Google Scholar] [CrossRef]
- Namkung, H.; Yu, H.; Gong, J.; Leeson, S. Antimicrobial Activity of Butyrate Glycerides toward Salmonella Typhimurium and Clostridium Perfringens. Poult. Sci. 2011, 90, 2217–2222. [Google Scholar] [CrossRef]
- Kovanda, L.L.; Hejna, M.; Liu, Y. 124 Butyric Acid and Derivatives: In Vitro Effects on Barrier Integrity of Porcine Intestinal Epithelial Cells Quantified by Transepithelial Electrical Resistance. J. Anim. Sci. 2020, 98, 109–110. [Google Scholar] [CrossRef]
- Asadullah, K.; Sterry, W.; Volk, H.D. Interleukin-10 Therapy—Review of a New Approach. Pharmacol. Rev. 2003, 55, 241–269. [Google Scholar] [CrossRef]
- Wang, H.; Chen, H.; Lin, Y.; Wang, G.; Luo, Y.; Li, X.; Wang, M.; Huai, M.; Li, L.; Barri, A. Butyrate Glycerides Protect against Intestinal Inflammation and Barrier Dysfunction in Mice. Nutrients 2022, 14, 3991. [Google Scholar] [CrossRef]
- Nguyen, T.D.; Prykhodko, O.; Hållenius, F.F.; Nyman, M. Monobutyrin Reduces Liver Cholesterol and Improves Intestinal Barrier Function in Rats Fed High-Fat Diets. Nutrients 2019, 11, 308. [Google Scholar] [CrossRef]
- Kabara, J.J. Lipids as Host-Resistance Factors of Human Milk. Nutr. Rev. 1980, 38, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Kabara, J.J. Symposium on the Pharmacological Effect of Lipids; The American Oil Chemists Society: Champaign, IL, USA, 1978; ISBN 978-99918-1-769-9. [Google Scholar]
- Isaacs, C.E. Human Milk Inactivates Pathogens Individually, Additively, and Synergistically. J. Nutr. 2005, 135, 1286–1288. [Google Scholar] [CrossRef] [PubMed]
- Anacarso, I.; Quartieri, A.; De Leo, R.; Pulvirenti, A. Evaluation of the Antimicrobial Activity of a Blend of Monoglycerides against Escherichia Coli and Enterococci with Multiple Drug Resistance. Arch. Microbiol. 2018, 200, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Xu, E.; Chen, C.; Fu, J.; Zhu, L.; Shu, J.; Jin, M.; Wang, Y.; Zong, X. Dietary Fatty Acids in Gut Health: Absorption, Metabolism and Function. Anim. Nutr. 2021, 7, 1337–1344. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.K.; Arsenakis, M.; Coelen, R.J.; May, J.T. Effect of Antiviral Lipids, Heat, and Freezing on the Activity of Viruses in Human Milk. J. Infect. Dis. 1979, 140, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.K.; Skurrie, I.J.; May, J.T. Use of Semliki Forest Virus to Identify Lipid-Mediated Antiviral Activity and Anti-Alphavirus Immunoglobulin A in Human Milk. Infect. Immun. 1978, 19, 395–401. [Google Scholar] [CrossRef]
- Thormar, H.; Isaacs, C.E.; Brown, H.R.; Barshatzky, M.R.; Pessolano, T. Inactivation of Enveloped Viruses and Killing of Cells by Fatty Acids and Monoglycerides. Antimicrob. Agents Chemother. 1987, 31, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Thormar, H.; Isaacs, C.E.; Soo Kim, K.; Brown, H.R. Inactivation of Visna Virus and Other Enveloped Viruses by Free Fatty Acids and Monoglycerides. Ann. N. Y. Acad. Sci. 1994, 724, 465–471. [Google Scholar] [CrossRef]
- Jackman, J.A.; Hakobyan, A.; Zakaryan, H.; Elrod, C.C. Inhibition of African Swine Fever Virus in Liquid and Feed by Medium-Chain Fatty Acids and Glycerol Monolaurate. J. Anim. Sci. Biotechnol. 2020, 11, 114. [Google Scholar] [CrossRef]
- Phillips, F.C.; Rubach, J.K.; Poss, M.J.; Anam, S.; Goyal, S.M.; Dee, S.A. Monoglyceride Reduces Viability of Porcine Epidemic Diarrhoea Virus in Feed and Prevents Disease Transmission to Post-Weaned Piglets. Transbound. Emerg. Dis. 2022, 69, 121–127. [Google Scholar] [CrossRef]
- Jackman, J.A.; Boyd, R.D.; Elrod, C.C. Medium-Chain Fatty Acids and Monoglycerides as Feed Additives for Pig Production: Towards Gut Health Improvement and Feed Pathogen Mitigation. J. Anim. Sci. Biotechnol. 2020, 11, 44. [Google Scholar] [CrossRef] [PubMed]
- Hariastuti, N.I. Avian Influenza Virus Inactivation by Caprylic Acid, Sodium Caprylate, and Monocaprylin. Health Sci. J. Indones. 2012, 2, 41–45. [Google Scholar]
- Mellouk, A.; Goossens, T.; Quinteiro-Filho, W.; Michel, V.; Vieco, N.; Lemâle, O.; Yakout, H.; Consuegra, J. Glycerides of Lauric Acid Supplementation in the Chicken Diet Enhances the Humoral and Cellular Immune Response to Infectious Bronchitis Virus. In Proceedings of the Poultry Science Association Annual Meeting, Philadelphia, PA, USA, 10–13 July 2023; Volume 102 (E-Supplement 1), p. 84. [Google Scholar]
- Zhang, J.; Feng, F.; Zhao, M. Glycerol Monocaprylate Modulates Gut Microbiota and Increases Short-Chain Fatty Acids Production without Adverse Effects on Metabolism and Inflammation. Nutrients 2021, 13, 1427. [Google Scholar] [CrossRef]
- Liu, T.; Guo, L.; Zhangying, Y.; Ruan, S.; Liu, W.; Zhang, X.; Feng, F. Dietary Medium-Chain 1-Monoglycerides Modulates the Community and Function of Cecal Microbiota of Broilers. J. Sci. Food Agric. 2022, 102, 2242–2252. [Google Scholar] [CrossRef]
- Liu, T.; Tang, J.; Feng, F. Medium-Chain α-Monoglycerides Improves Productive Performance and Egg Quality in Aged Hens Associated with Gut Microbiota Modulation. Poult. Sci. 2020, 99, 7122–7132. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Jiang, Z.; Cai, H.; Li, Y.; Mo, Q.; Deng, L.; Zhong, H.; Liu, T.; Zhang, H.; Kang, J.X.; et al. Modulation of the Gut Microbiota during High-Dose Glycerol Monolaurate-Mediated Amelioration of Obesity in Mice Fed a High-Fat Diet. mBio 2020, 11, e00190-20. [Google Scholar] [CrossRef]
- Kong, L.; Wang, Z.; Xiao, C.; Zhu, Q.; Song, Z. Glycerol Monolaurate Ameliorated Intestinal Barrier and Immunity in Broilers by Regulating Intestinal Inflammation, Antioxidant Balance, and Intestinal Microbiota. Front. Immunol. 2021, 12, 713485. [Google Scholar] [CrossRef]
- Sacakli, P.; Çınar, Ö.Ö.; Ceylan, A.; Ramay, M.S.; Harijaona, J.A.; Bayraktaroglu, A.G.; Shastak, Y.; Calik, A. Performance and Gut Health Status of Broilers Fed Diets Supplemented with Two Graded Levels of a Monoglyceride Blend. Poult. Sci. 2023, 102, 102359. [Google Scholar] [CrossRef]
- Kong, L.; Wang, Z.; Xiao, C.; Zhu, Q.; Song, Z. Glycerol Monolaurate Attenuated Immunological Stress and Intestinal Mucosal Injury by Regulating the Gut Microbiota and Activating AMPK/Nrf2 Signaling Pathway in Lipopolysaccharide-Challenged Broilers. Anim. Nutr. 2022, 10, 347–359. [Google Scholar] [CrossRef]
- Grochowska, M.; Laskus, T.; Paciorek, M.; Pollak, A.; Lechowicz, U.; Makowiecki, M.; Horban, A.; Radkowski, M.; Perlejewski, K. Patients with Infections of The Central Nervous System Have Lowered Gut Microbiota Alpha Diversity. Curr. Issues Mol. Biol. 2022, 44, 2903–2914. [Google Scholar] [CrossRef]
- Zhong, X.; Harrington, J.M.; Millar, S.R.; Perry, I.J.; O’Toole, P.W.; Phillips, C.M. Gut Microbiota Associations with Metabolic Health and Obesity Status in Older Adults. Nutrients 2020, 12, 2364. [Google Scholar] [CrossRef]
- Hagerty, S.L.; Hutchison, K.E.; Lowry, C.A.; Bryan, A.D. An Empirically Derived Method for Measuring Human Gut Microbiome Alpha Diversity: Demonstrated Utility in Predicting Health-Related Outcomes among a Human Clinical Sample. PLoS ONE 2020, 15, e0229204. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, J.; Liang, H.; Ye, L.; Lan, L.; Lu, F.; Wang, Q.; Lei, T.; Yang, X.; Cui, P.; et al. Differences in Alpha Diversity of Gut Microbiota in Neurological Diseases. Front. Neurosci. 2022, 16, 879318. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kheravii, S.K.; Li, L.; Wu, S.-B. Monoglyceride Blend Reduces Mortality, Improves Nutrient Digestibility, and Intestinal Health in Broilers Subjected to Clinical Necrotic Enteritis Challenge. Animals 2021, 11, 1432. [Google Scholar] [CrossRef] [PubMed]
- Meslin, C.; Desert, C.; Callebaut, I.; Djari, A.; Klopp, C.; Pitel, F.; Leroux, S.; Martin, P.; Froment, P.; Guilbert, E.; et al. Expanding Duplication of Free Fatty Acid Receptor-2 (GPR43) Genes in the Chicken Genome. Genome Biol. Evol. 2015, 7, 1332–1348. [Google Scholar] [CrossRef]
- Mielenz, M. Invited Review: Nutrient-Sensing Receptors for Free Fatty Acids and Hydroxycarboxylic Acids in Farm Animals. Animal 2017, 11, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Suh, H.N.; Huong, H.T.; Song, C.H.; Lee, J.H.; Han, H.J. Linoleic Acid Stimulates Gluconeogenesis via Ca2+/PLC, cPLA2, and PPAR Pathways through GPR40 in Primary Cultured Chicken Hepatocytes. Am. J. Physiol. Cell Physiol. 2008, 295, C1518–C1527. [Google Scholar] [CrossRef] [PubMed]
- Cordero, P.; Díaz-Avilés, F.; Torres, P.; Guzmán, M.; Niknafs, S.; Roura, E.; Guzmán-Pino, S.A. The Expression of Amino Acid and Fatty Acid Receptors Show an Age-Dependent Pattern Involving Oral Cavity, Jejunum and Lower Gut Sensing in Broiler Chickens. Animals 2023, 13, 3120. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.K.; Gu, M.J.; Ko, K.H.; Bae, S.; Kim, G.; Jin, G.-D.; Kim, E.B.; Kong, Y.-Y.; Park, T.S.; Park, B.-C.; et al. Regulation of CD4+CD8−CD25+ and CD4+CD8+CD25+ T Cells by Gut Microbiota in Chicken. Sci. Rep. 2018, 8, 8627. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, V.V.; Han, J.; Fasina, Y.; Harrison, S.H. Connecting Gut Microbiomes and Short Chain Fatty Acids with the Serotonergic System and Behavior in Gallus Gallus and Other Avian Species. Front. Physiol. 2022, 13, 1035538. [Google Scholar] [CrossRef] [PubMed]
- Kaemmerer, E.; Plum, P.; Klaus, C.; Weiskirchen, R.; Liedtke, C.; Adolf, M.; Schippers, A.; Wagner, N.; Reinartz, A.; Gassler, N. Fatty Acid Binding Receptors in Intestinal Physiology and Pathophysiology. World J. Gastrointest. Pathophysiol. 2010, 1, 147–153. [Google Scholar] [CrossRef] [PubMed]
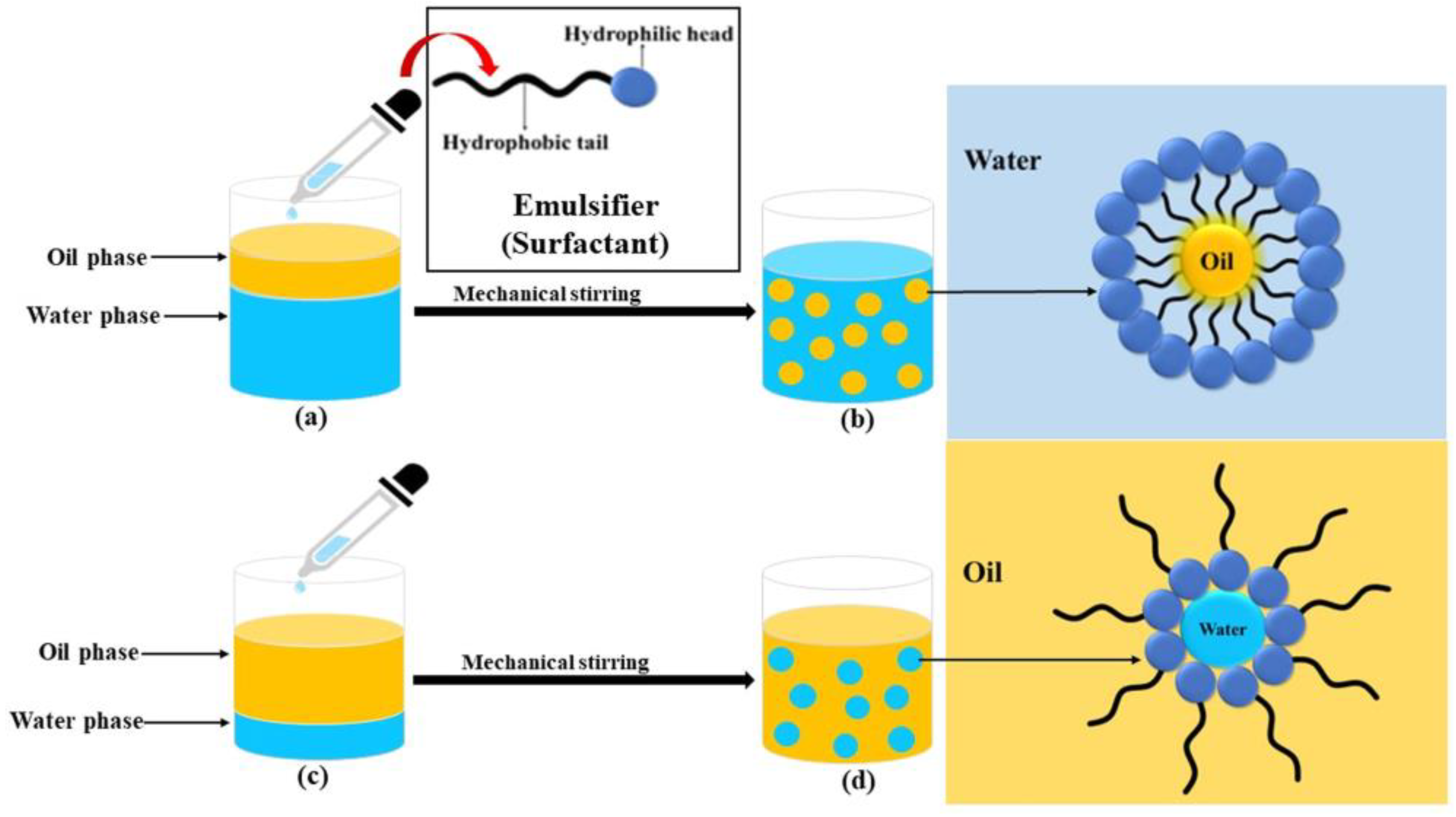

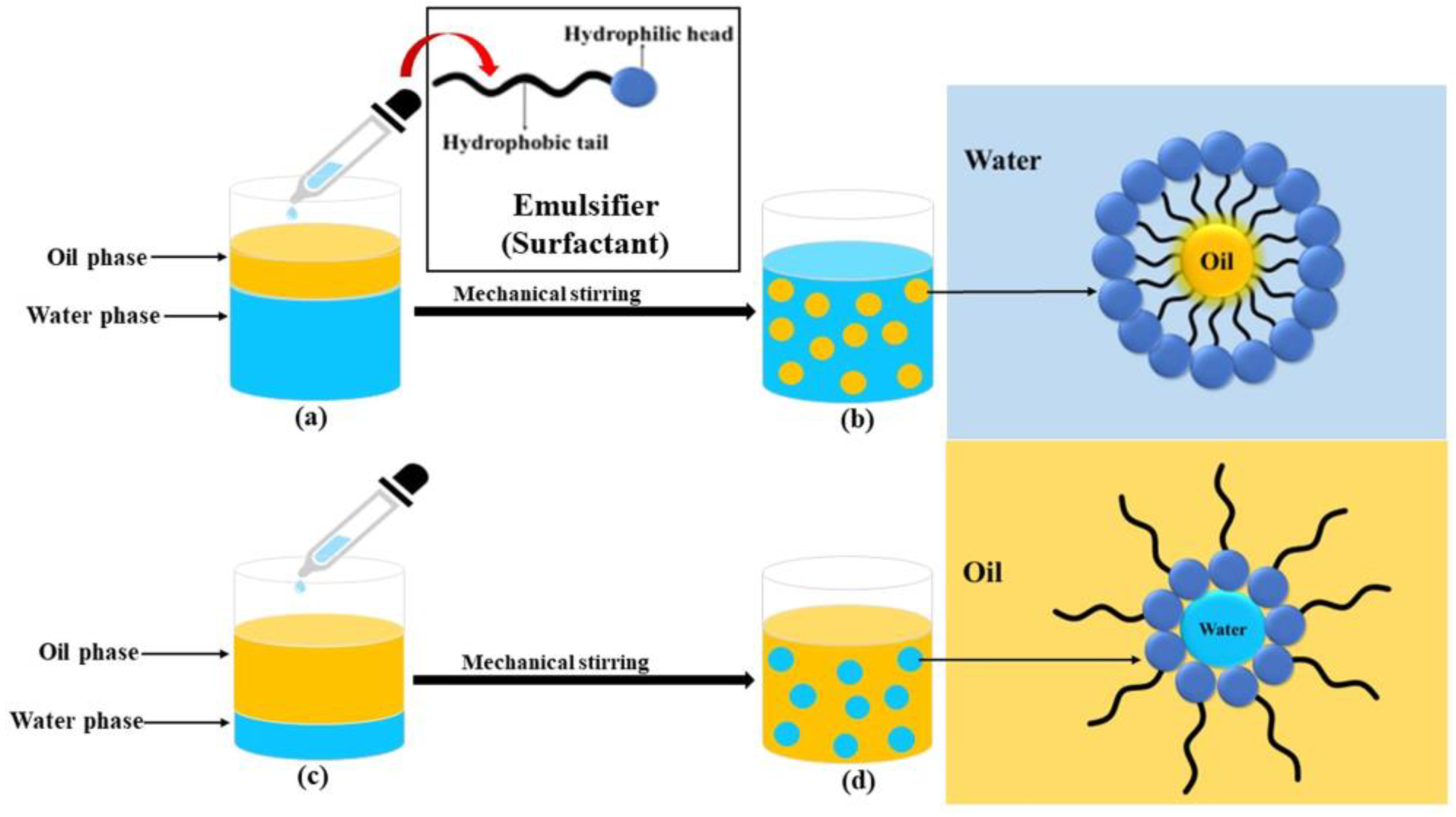{kind=link}
| Product Description | Animal Age, d | Inclusion Rates, % | Response Criteria in Broilers Except Where Noted | Literature Reference |
|---|---|---|---|---|
| Butyric, caprylic and capric acids | 0–42 | 0.03, 0.05 | gut structure and growth performance | [64] |
| Butyrate monoglycerides | 0–28 | 0.015 | intestinal barrier function and inflammation responses in mice | [43] |
| Butyrate mono-, di- and tri-glycerides | 0–42 | 0.2, 0.4 | growth performance, and carcass yield | [23] |
| 0.1, 0.2 | ||||
| 0–27 | 0.4 | |||
| 0–42 | 0.2, 0.35, 0.5, 1.0 | growth performance, intestinal morphology, and carcass yield | [24] | |
| 0–7 | 0.6 | growth performance, carcass composition, serum biochemistry, and gene expression | [25] | |
| 8–20 | 0.3 | |||
| Glycerol monocaprylate | 28–154 | 0.015, 0.16 | gut microbiota, short-chain fatty acid production, metabolism, and inflammation in mice | [59] |
| Glycerol monolaurate | 0–112 | 0.16 | gut microbiota and regulating glycerophospholipid metabolism in mice | [62] |
| 0–14 | 0.03, 0.06, 0.09, 0.12 | growth performance, immunity, intestinal barrier, and cecal microbiota | [63] | |
| 0–21 | 0.12 | immunological stress and intestinal mucosal injury | [65] | |
| Mixture of butyric, propionic, caprylic, capric, and lauric acid mono- and diglycerides | 0–35 | 0.05, 0.1, 0.2, 0.3 | growth performance, carcass composition, fat, and muscle gene expression | [11] |
| Mixture of medium chain alpha-monoglycerides | 0–42 | 0.03, 0.045, 0.06 | community and function of gut microbiota | [60] |
| 280–448 | 0.03 | productive performance, egg quality, serum biochemical indices, and gut microbiota in laying hens | [61] | |
| Mixture of monoglycerides | 1–14 | 0.5 | growth performance, nutrient utilization, and meat yield | [21] |
| 14–28 | 0.4 | |||
| 28–42 | 0.2 | |||
| 42–49 | 0.15, 0.25, 0.35 | viral disease transmission to piglets | [56] | |
| Mixture of mono- and di-glycerides | 0–43 | 0.5 | growth performance, nutrient digestibility, and energy utilization | [17] |
| Monobutyrin | 0–28 | 0.25, 0.75, 1.25 | lipid metabolism and gut barrier function | [44] |
| Monobutyrin and/or tributyrin | 1–35 | 0.05 tributyrin + 0.05 monobutyrin | growth performance, carcass composition, and blood biochemistry | [26] |
| 0.05 tributyrin + 0.2 monobutyrin | ||||
| 1–7 | 0.05 tributyrin | |||
| 8–14 | 0.05 or 0.2 monobutyrin | |||
| 15–35 | no glycerides | |||
| Monolaurate | 3–35 | 0.1, 0.3, 0.5 | growth performance, nutrient utilization, and blood biochemistry | [10] |
| Monolaurate and Monodecanoate | 0–70 | 0.03 | growth performance, intestinal microbiota, intestinal morphology, and blood chemistry | [12] |
| carcass composition | [22] | |||
| Monostearate | 18–42 | 0.02 | growth performance, carcass composition, serum biochemistry, and liver biochemistry | [20] |
| Ricinoleate | 0–21 | 0.035, 0.07 | growth performance, nutrient metabolism, and blood chemistry | [18] |
| 22–39 | 0.028, 0.056 |
| Product Description | Inclusion Rates, % | Response Criteria | Literature Reference |
|---|---|---|---|
| Butyrate glycerides | 0.025, 0.7 | antibacterial activity against Salmonella typhimurium and Clostridium perfringens | [40] |
| Monobutyrin | 20 pg | angiogenesis in chick chorioallantoic membrane | [15] |
| Mixture of monoglycerides | 0.05–2 | antibacterial activity against E. coli and Enterococcus | [48] |
| Monocaprylin | 0.2, 0.4 | anti-viral activity | [55] |
| Monocaprin, monolaurin, monomyristin, monopalmitin, and monostearin | 0.016–1.0 | antimicrobial activity | [37] |
| Monocaprylate, monocaprin, monolaurin, and monomyristin | 0.5 | antimicrobial activity | [28] |
| Monolaurate | 2 | antimicrobial activity | [32] |
| 0.05–0.1 | antimicrobial activity | [33] | |
| 0.1–0.3 | antimicrobial activity | [34] | |
| 0.01 | antimicrobial activity | [36] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Appleton, S.R.; Ballou, A.; Watkins, K.L. Use of Monoglycerides and Diglycerides to Mitigate Poultry Production Losses: A Review. Vet. Sci. 2024, 11, 101. https://doi.org/10.3390/vetsci11030101
Appleton SR, Ballou A, Watkins KL. Use of Monoglycerides and Diglycerides to Mitigate Poultry Production Losses: A Review. Veterinary Sciences. 2024; 11(3):101. https://doi.org/10.3390/vetsci11030101
Chicago/Turabian StyleAppleton, Stacie R., Anne Ballou, and Kevin L. Watkins. 2024. "Use of Monoglycerides and Diglycerides to Mitigate Poultry Production Losses: A Review" Veterinary Sciences 11, no. 3: 101. https://doi.org/10.3390/vetsci11030101
APA StyleAppleton, S. R., Ballou, A., & Watkins, K. L. (2024). Use of Monoglycerides and Diglycerides to Mitigate Poultry Production Losses: A Review. Veterinary Sciences, 11(3), 101. https://doi.org/10.3390/vetsci11030101