
Deciphering the Dual Role of Heligmosomoides polygyrus Antigens in Macrophage Modulation and Breast Cancer Cell Growth



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. H. polygyrus Antigens
2.2. Cell Culture
2.3. MTT Assay
2.4. RT PCR
2.5. Flow Cytometry Analysis
2.6. Ethics
2.7. Statistics
3. Results
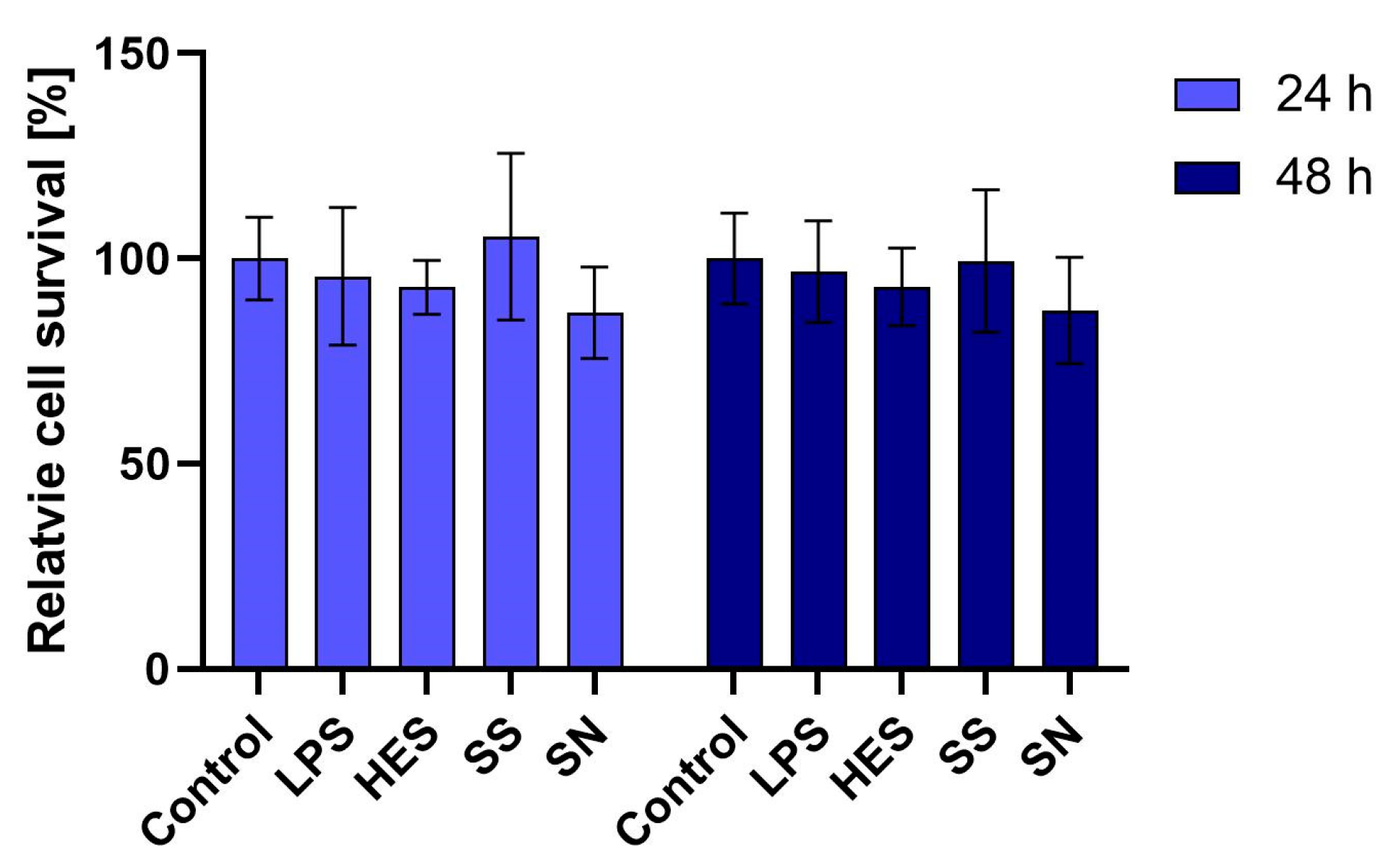
3.1. Impact on Macrophage Viability
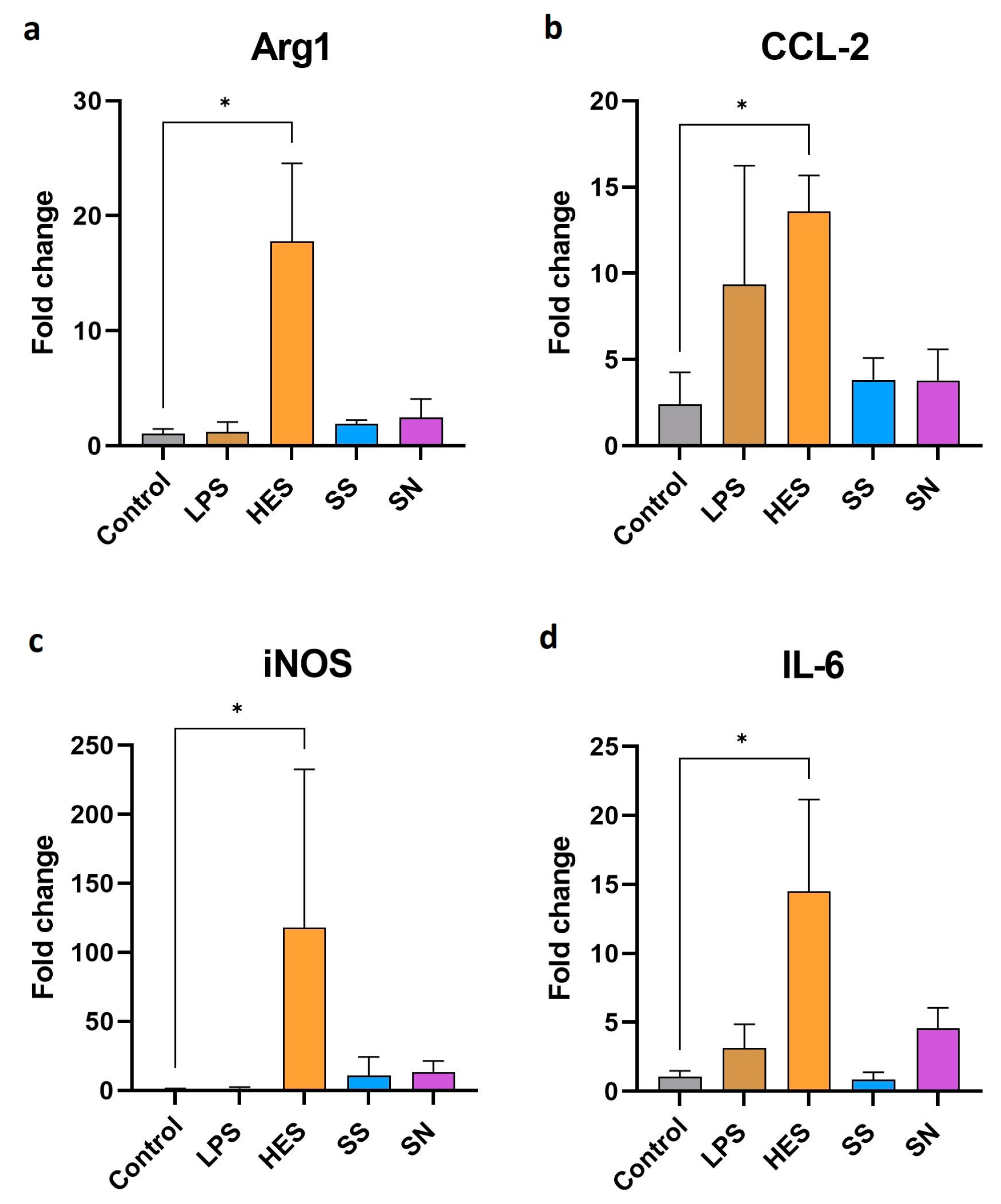
3.2. Gene Expression Changes in Macrophages
3.3. Macrophage Surface Receptor Modulation
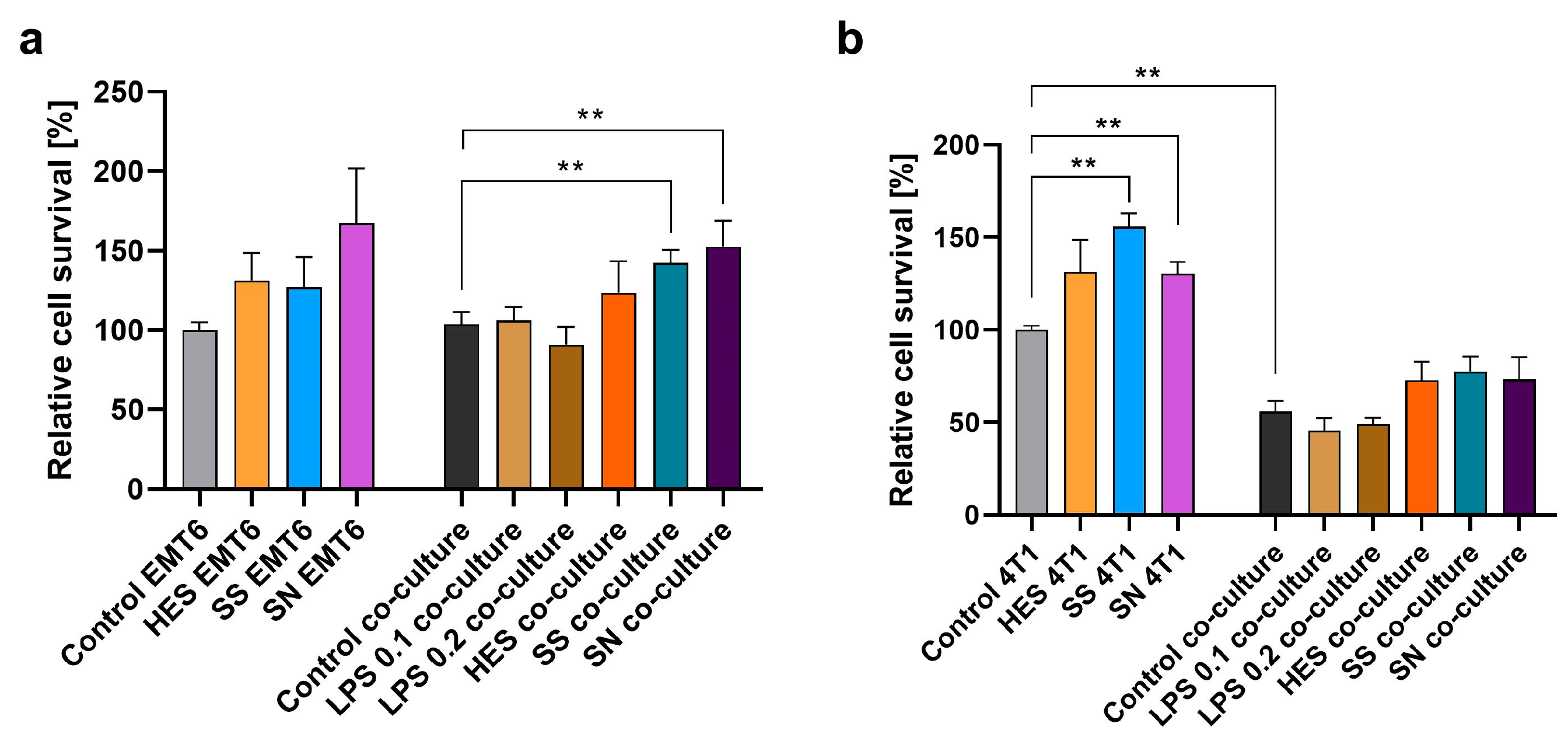
3.4. Effect on Breast Cancer Cell Proliferation
3.5. Overview
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Behnke, J.M.; Wahid, F.N. Immunological Relationships during Primary Infection with Heligmosomoides polygyrus (Nematospiroides dubius): H-2 Linked Genes Determine Worm Survival. Parasitology 1991, 103, 157–164. [Google Scholar] [CrossRef]
- Gouÿ De Bellocq, J.; Ferté, H.; Depaquit, J.; Justine, J.-L.; Tillier, A.; Durette-Desset, M.-C. Phylogeny of the Trichostrongylina (Nematoda) Inferred from 28S rDNA Sequences. Mol. Phylogenet. Evol. 2001, 19, 430–442. [Google Scholar] [CrossRef]
- Ludwig-Portugall, I.; Layland, L.E. TLRs, Treg, and B Cells, an Interplay of Regulation during Helminth Infection. Front. Immun. 2012, 3, 8. [Google Scholar] [CrossRef]
- Maizels, R.M.; Smith, K.A. Regulatory T Cells in Infection. In Advances in Immunology; Elsevier: Amsterdam, The Netherlands, 2011; Volume 112, pp. 73–136. ISBN 978-0-12-387827-4. [Google Scholar]
- Medzhitov, R.; Schneider, D.S.; Soares, M.P. Disease Tolerance as a Defense Strategy. Science 2012, 335, 936–941. [Google Scholar] [CrossRef]
- Bobardt, S.D.; Dillman, A.R.; Nair, M.G. The Two Faces of Nematode Infection: Virulence and Immunomodulatory Molecules From Nematode Parasites of Mammals, Insects and Plants. Front. Microbiol. 2020, 11, 577846. [Google Scholar] [CrossRef]
- Van Dyken, S.J.; Locksley, R.M. Interleukin-4- and Interleukin-13-Mediated Alternatively Activated Macrophages: Roles in Homeostasis and Disease. Annu. Rev. Immunol. 2013, 31, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Tarique, A.A.; Logan, J.; Thomas, E.; Holt, P.G.; Sly, P.D.; Fantino, E. Phenotypic, Functional, and Plasticity Features of Classical and Alternatively Activated Human Macrophages. Am. J. Respir. Cell Mol. Biol. 2015, 53, 676–688. [Google Scholar] [CrossRef] [PubMed]
- Rolot, M.; Dewals, B.G. Macrophage Activation and Functions during Helminth Infection: Recent Advances from the Laboratory Mouse. J. Immunol. Res. 2018, 2018, 2790627. [Google Scholar] [CrossRef] [PubMed]
- Coakley, G.; McCaskill, J.L.; Borger, J.G.; Simbari, F.; Robertson, E.; Millar, M.; Harcus, Y.; McSorley, H.J.; Maizels, R.M.; Buck, A.H. Extracellular Vesicles from a Helminth Parasite Suppress Macrophage Activation and Constitute an Effective Vaccine for Protective Immunity. Cell Rep. 2017, 19, 1545–1557. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J. Macrophage Polarization. Annu. Rev. Physiol. 2017, 79, 541–566. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Werb, Z. Inflammation and Cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Qian, B.-Z.; Pollard, J.W. Macrophage Diversity Enhances Tumor Progression and Metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Sica, A. Macrophages, Innate Immunity and Cancer: Balance, Tolerance, and Diversity. Curr. Opin. Immunol. 2010, 22, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Ojalvo, L.S.; Whittaker, C.A.; Condeelis, J.S.; Pollard, J.W. Gene Expression Analysis of Macrophages That Facilitate Tumor Invasion Supports a Role for Wnt-Signaling in Mediating Their Activity in Primary Mammary Tumors. J. Immunol. 2010, 184, 702–712. [Google Scholar] [CrossRef]
- Bingle, L.; Brown, N.J.; Lewis, C.E. The Role of Tumour-Associated Macrophages in Tumour Progression: Implications for New Anticancer Therapies. J. Pathol. 2002, 196, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Lin, E.Y.; Nguyen, A.V.; Russell, R.G.; Pollard, J.W. Colony-Stimulating Factor 1 Promotes Progression of Mammary Tumors to Malignancy. J. Exp. Med. 2001, 193, 727–740. [Google Scholar] [CrossRef]
- Pollard, J.W. Tumour-Educated Macrophages Promote Tumour Progression and Metastasis. Nat. Rev. Cancer 2004, 4, 71–78. [Google Scholar] [CrossRef]
- Jacobs, B.-A.; Prince, S.; Smith, K.A. Gastrointestinal Nematode-Derived Antigens Alter Colorectal Cancer Cell Proliferation and Migration through Regulation of Cell Cycle and Epithelial-Mesenchymal Transition Proteins. Int. J. Mol. Sci. 2020, 21, 7845. [Google Scholar] [CrossRef]
- Callejas, B.E.; Martínez-Saucedo, D.; Terrazas, L.I. Parasites as Negative Regulators of Cancer. Biosci. Rep. 2018, 38, BSR20180935. [Google Scholar] [CrossRef]
- Sripa, B.; Brindley, P.J.; Mulvenna, J.; Laha, T.; Smout, M.J.; Mairiang, E.; Bethony, J.M.; Loukas, A. The Tumorigenic Liver Fluke Opisthorchis Viverrini—Multiple Pathways to Cancer. Trends Parasitol. 2012, 28, 395–407. [Google Scholar] [CrossRef]
- Arora, N.; Kaur, R.; Anjum, F.; Tripathi, S.; Mishra, A.; Kumar, R.; Prasad, A. Neglected Agent Eminent Disease: Linking Human Helminthic Infection, Inflammation, and Malignancy. Front. Cell. Infect. Microbiol. 2019, 9, 402. [Google Scholar] [CrossRef]
- Ehata, S.; Hanyu, A.; Hayashi, M.; Aburatani, H.; Kato, Y.; Fujime, M.; Saitoh, M.; Miyazawa, K.; Imamura, T.; Miyazono, K. Transforming Growth Factor-β Promotes Survival of Mammary Carcinoma Cells through Induction of Antiapoptotic Transcription Factor DEC1. Cancer Res. 2007, 67, 9694–9703. [Google Scholar] [CrossRef]
- Ehata, S.; Hanyu, A.; Fujime, M.; Katsuno, Y.; Fukunaga, E.; Goto, K.; Ishikawa, Y.; Nomura, K.; Yokoo, H.; Shimizu, T.; et al. Ki26894, a Novel Transforming Growth Factor-? Type I Receptor Kinase Inhibitor, Inhibits in Vitro Invasion and in Vivo Bone Metastasis of a Human Breast Cancer Cell Line. Cancer Sci. 2007, 98, 127–133. [Google Scholar] [CrossRef]
- Johnston, C.J.C.; Smyth, D.J.; Kodali, R.B.; White, M.P.J.; Harcus, Y.; Filbey, K.J.; Hewitson, J.P.; Hinck, C.S.; Ivens, A.; Kemter, A.M.; et al. A Structurally Distinct TGF-β Mimic from an Intestinal Helminth Parasite Potently Induces Regulatory T Cells. Nat. Commun. 2017, 8, 1741. [Google Scholar] [CrossRef]
- Tao, K.; Fang, M.; Alroy, J.; Sahagian, G.G. Imagable 4T1 Model for the Study of Late Stage Breast Cancer. BMC Cancer 2008, 8, 228. [Google Scholar] [CrossRef]
- White, M.P.J.; Smyth, D.J.; Cook, L.; Ziegler, S.F.; Levings, M.K.; Maizels, R.M. The Parasite Cytokine Mimic Hp -TGM Potently Replicates the Regulatory Effects of TGF-β on Murine CD4+ T Cells. Immunol. Cell Biol. 2021, 99, 848–864. [Google Scholar] [CrossRef] [PubMed]
- Gorczynski, R.M.; Chen, Z.; Erin, N.; Khatri, I.; Podnos, A. Comparison of Immunity in Mice Cured of Primary/Metastatic Growth of EMT6 or 4THM Breast Cancer by Chemotherapy or Immunotherapy. PLoS ONE 2014, 9, e113597. [Google Scholar] [CrossRef] [PubMed]
- Ouzounova, M.; Lee, E.; Piranlioglu, R.; El Andaloussi, A.; Kolhe, R.; Demirci, M.F.; Marasco, D.; Asm, I.; Chadli, A.; Hassan, K.A.; et al. Monocytic and Granulocytic Myeloid Derived Suppressor Cells Differentially Regulate Spatiotemporal Tumour Plasticity during Metastatic Cascade. Nat. Commun. 2017, 8, 14979. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Jung, J.I.; Lim, D.Y.; Kwon, G.T.; Her, S.; Park, J.H.; Park, J.H.Y. Bone Marrow-Derived, Alternatively Activated Macrophages Enhance Solid Tumor Growth and Lung Metastasis of Mammary Carcinoma Cells in a Balb/C Mouse Orthotopic Model. Breast Cancer Res. 2012, 14, R81. [Google Scholar] [CrossRef] [PubMed]
- Anthony, R.M.; Urban, J.F.; Alem, F.; Hamed, H.A.; Rozo, C.T.; Boucher, J.-L.; Van Rooijen, N.; Gause, W.C. Memory TH2 Cells Induce Alternatively Activated Macrophages to Mediate Protection against Nematode Parasites. Nat. Med. 2006, 12, 955–960. [Google Scholar] [CrossRef] [PubMed]
- Rzepecka, J.; Lucius, R.; Doligalska, M.; Beck, S.; Rausch, S.; Hartmann, S. Screening for Immunomodulatory Proteins of the Intestinal Parasitic Nematode Heligmosomoides polygyrus. Parasite Immunol. 2006, 28, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Doligalska, M.; Brodaczewska, K.; Donskow-Łysoniewska, K. The Antiapoptotic Activity of Heligmosomoides Polygyrus Antigen Fractions. Parasite Immunol. 2012, 34, 589–603. [Google Scholar] [CrossRef] [PubMed]
- Xue, Q.; Yan, Y.; Zhang, R.; Xiong, H. Regulation of iNOS on Immune Cells and Its Role in Diseases. Int. J. Mol. Sci. 2018, 19, 3805. [Google Scholar] [CrossRef] [PubMed]
- Franco, L.H.; Fleuri, A.K.A.; Pellison, N.C.; Quirino, G.F.S.; Horta, C.V.; De Carvalho, R.V.H.; Oliveira, S.C.; Zamboni, D.S. Autophagy Downstream of Endosomal Toll-like Receptor Signaling in Macrophages Is a Key Mechanism for Resistance to Leishmania Major Infection. J. Biol. Chem. 2017, 292, 13087–13096. [Google Scholar] [CrossRef]
- Van Der Kleij, D.; Latz, E.; Brouwers, J.F.H.M.; Kruize, Y.C.M.; Schmitz, M.; Kurt-Jones, E.A.; Espevik, T.; De Jong, E.C.; Kapsenberg, M.L.; Golenbock, D.T.; et al. A Novel Host-Parasite Lipid Cross-Talk. J. Biol. Chem. 2002, 277, 48122–48129. [Google Scholar] [CrossRef]
- Lin, Z.; Shi, J.-L.; Chen, M.; Zheng, Z.-M.; Li, M.-Q.; Shao, J. CCL2: An Important Cytokine in Normal and Pathological Pregnancies: A Review. Front. Immunol. 2023, 13, 1053457. [Google Scholar] [CrossRef]
- Avraham, R.; Haseley, N.; Brown, D.; Penaranda, C.; Jijon, H.B.; Trombetta, J.J.; Satija, R.; Shalek, A.K.; Xavier, R.J.; Regev, A.; et al. Pathogen Cell-to-Cell Variability Drives Heterogeneity in Host Immune Responses. Cell 2015, 162, 1309–1321. [Google Scholar] [CrossRef]
- Maizels, R.M.; Hewitson, J.P.; Murray, J.; Harcus, Y.M.; Dayer, B.; Filbey, K.J.; Grainger, J.R.; McSorley, H.J.; Reynolds, L.A.; Smith, K.A. Immune Modulation and Modulators in Heligmosomoides Polygyrus Infection. Exp. Parasitol. 2012, 132, 76–89. [Google Scholar] [CrossRef]
- Vandenberg, T.; Kraal, G. A Function for the Macrophage F4/80 Molecule in Tolerance Induction. Trends Immunol. 2005, 26, 506–509. [Google Scholar] [CrossRef]
- Rodríguez-Sosa, M.; Satoskar, A.R.; Calderón, R.; Gomez-Garcia, L.; Saavedra, R.; Bojalil, R.; Terrazas, L.I. Chronic Helminth Infection Induces Alternatively Activated Macrophages Expressing High Levels of CCR5 with Low Interleukin-12 Production and Th2-Biasing Ability. Infect. Immun. 2002, 70, 3656–3664. [Google Scholar] [CrossRef]
- Schick, J.; Altunay, M.; Lacorcia, M.; Marschner, N.; Westermann, S.; Schluckebier, J.; Schubart, C.; Bodendorfer, B.; Christensen, D.; Alexander, C.; et al. IL-4 and Helminth Infection Downregulate MINCLE-Dependent Macrophage Response to Mycobacteria and Th17 Adjuvanticity. eLife 2023, 12, e72923. [Google Scholar] [CrossRef] [PubMed]
- Weng, M.; Huntley, D.; Huang, I.-F.; Foye-Jackson, O.; Wang, L.; Sarkissian, A.; Zhou, Q.; Walker, W.A.; Cherayil, B.J.; Shi, H.N. Alternatively Activated Macrophages in Intestinal Helminth Infection: Effects on Concurrent Bacterial Colitis. J. Immunol. 2007, 179, 4721–4731. [Google Scholar] [CrossRef] [PubMed]
- Xaus, J.; Comalada, M.; Valledor, A.F.; Lloberas, J.; López-Soriano, F.; Argilés, J.M.; Bogdan, C.; Celada, A. LPS Induces Apoptosis in Macrophages Mostly through the Autocrine Production of TNF-Alpha. Blood 2000, 95, 3823–3831. [Google Scholar] [CrossRef] [PubMed]
- Osbourn, M.; Soares, D.C.; Vacca, F.; Cohen, E.S.; Scott, I.C.; Gregory, W.F.; Smyth, D.J.; Toivakka, M.; Kemter, A.M.; Le Bihan, T.; et al. HpARI Protein Secreted by a Helminth Parasite Suppresses Interleukin-33. Immunity 2017, 47, 739–751.e5. [Google Scholar] [CrossRef]
- Ramanan, D.; Bowcutt, R.; Lee, S.C.; Tang, M.S.; Kurtz, Z.D.; Ding, Y.; Honda, K.; Gause, W.C.; Blaser, M.J.; Bonneau, R.A.; et al. Helminth Infection Promotes Colonization Resistance via Type 2 Immunity. Science 2016, 352, 608–612. [Google Scholar] [CrossRef] [PubMed]
- Harnett, M.M.; Harnett, W. Can Parasitic Worms Cure the Modern World’s Ills? Trends Parasitol. 2017, 33, 694–705. [Google Scholar] [CrossRef][Green Version]
- Wammes, L.J.; Mpairwe, H.; Elliott, A.M.; Yazdanbakhsh, M. Helminth Therapy or Elimination: Epidemiological, Immunological, and Clinical Considerations. Lancet Infect. Dis. 2014, 14, 1150–1162. [Google Scholar] [CrossRef]
- Shi, X.; Yang, J.; Deng, S.; Xu, H.; Wu, D.; Zeng, Q.; Wang, S.; Hu, T.; Wu, F.; Zhou, H. TGF-β Signaling in the Tumor Metabolic Microenvironment and Targeted Therapies. J. Hematol. Oncol. 2022, 15, 135. [Google Scholar] [CrossRef]
- Zhang, Y.; Alexander, P.B.; Wang, X.-F. TGF-β Family Signaling in the Control of Cell Proliferation and Survival. Cold Spring Harb. Perspect. Biol. 2017, 9, a022145. [Google Scholar] [CrossRef]
- Maruszewska-Cheruiyot, M.; Krawczak-Wójcik, K.; Michniowska, M.; Stear, M.J.; Machcińska, M.; Doligalska, M.; Donskow-Łysoniewska, K. Nematode-Induced Growth Factors Related to Angiogenesis in Autoimmune Disease Attenuation. Life 2023, 13, 321. [Google Scholar] [CrossRef]
- Wang, H.; Tian, T.; Zhang, J. Tumor-Associated Macrophages (TAMs) in Colorectal Cancer (CRC): From Mechanism to Therapy and Prognosis. Int. J. Mol. Sci. 2021, 22, 8470. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Firmanty, P.; Doligalska, M.; Krol, M.; Taciak, B. Deciphering the Dual Role of Heligmosomoides polygyrus Antigens in Macrophage Modulation and Breast Cancer Cell Growth. Vet. Sci. 2024, 11, 69. https://doi.org/10.3390/vetsci11020069
Firmanty P, Doligalska M, Krol M, Taciak B. Deciphering the Dual Role of Heligmosomoides polygyrus Antigens in Macrophage Modulation and Breast Cancer Cell Growth. Veterinary Sciences. 2024; 11(2):69. https://doi.org/10.3390/vetsci11020069
Chicago/Turabian StyleFirmanty, Patryk, Maria Doligalska, Magdalena Krol, and Bartlomiej Taciak. 2024. "Deciphering the Dual Role of Heligmosomoides polygyrus Antigens in Macrophage Modulation and Breast Cancer Cell Growth" Veterinary Sciences 11, no. 2: 69. https://doi.org/10.3390/vetsci11020069
APA StyleFirmanty, P., Doligalska, M., Krol, M., & Taciak, B. (2024). Deciphering the Dual Role of Heligmosomoides polygyrus Antigens in Macrophage Modulation and Breast Cancer Cell Growth. Veterinary Sciences, 11(2), 69. https://doi.org/10.3390/vetsci11020069