
The Microbiota Architecture of the Chinchilla Gastrointestinal Tract



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Selection
2.2. Sample Collection
2.3. DNA Extraction and 16S rRNA Sequencing
2.4. Statistical Analysis
3. Results
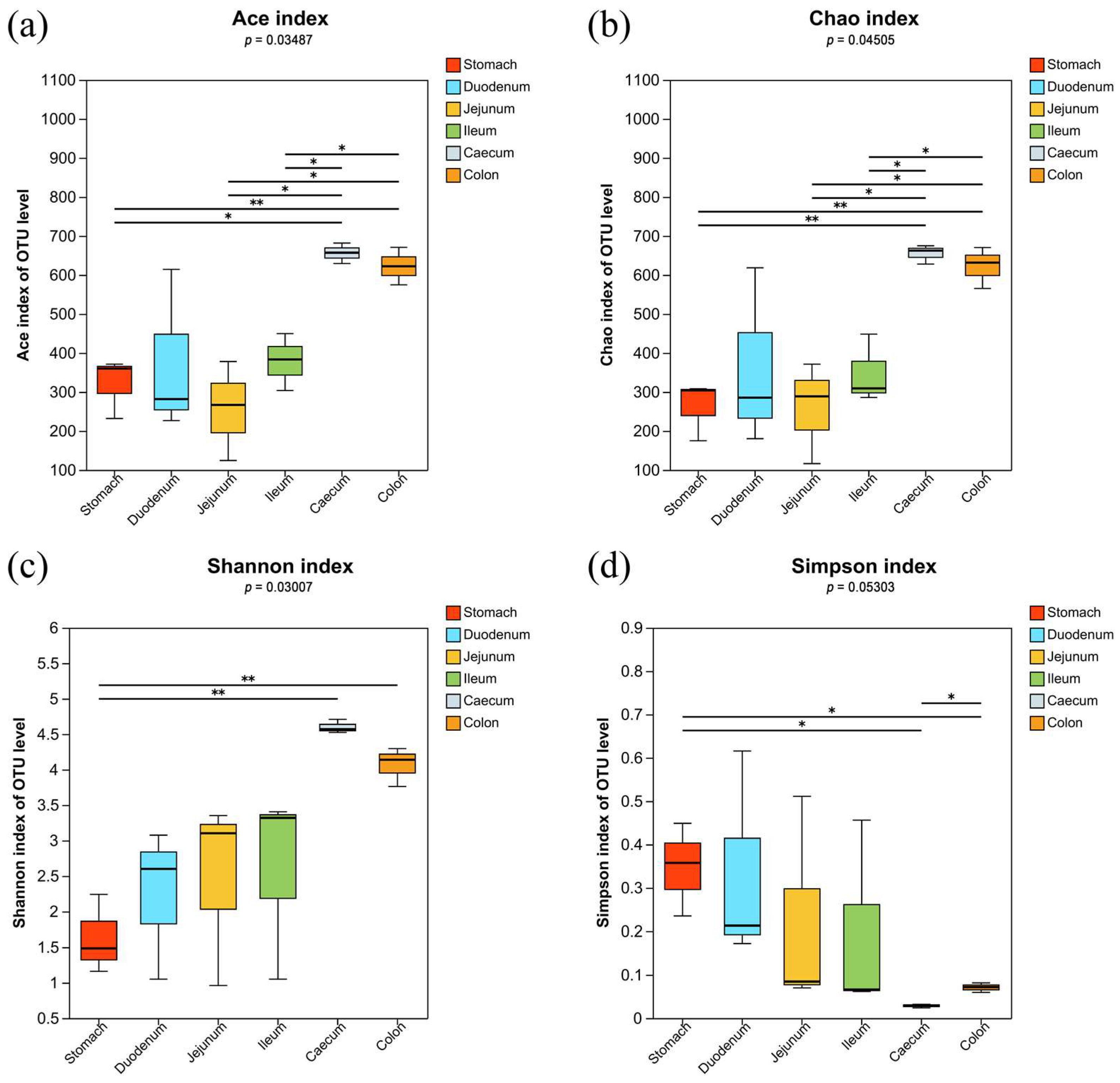
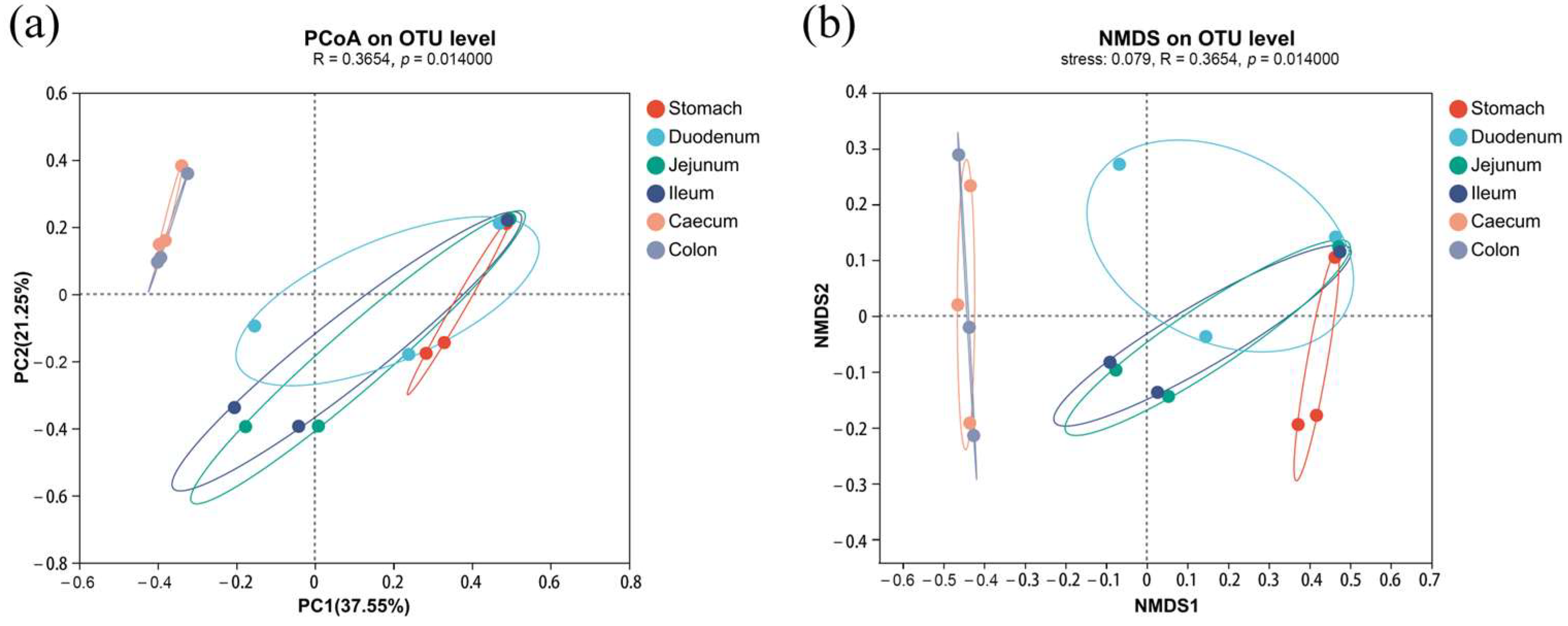
3.1. Microbiota Profile Analysis
3.2. Compositional Analysis
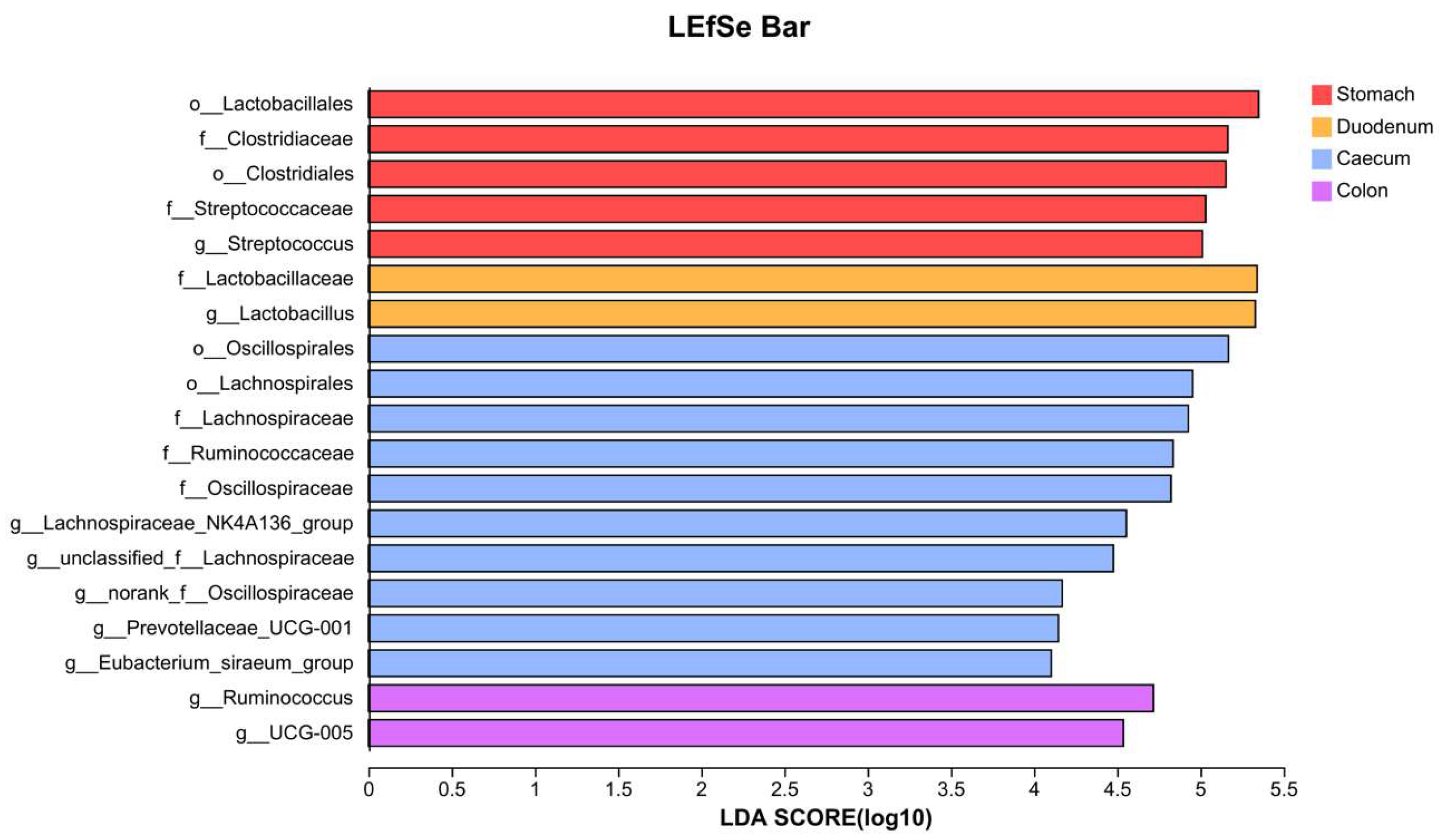
3.3. Gastrointestinal Signature Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Nageshwar Reddy, D. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef]
- Kohles, M. Gastrointestinal anatomy and physiology of select exotic companion mammals. Vet. Clin. North. Am. Exot. Anim. Pract. 2014, 17, 165–178. [Google Scholar] [CrossRef]
- Björnhag, G. Adaptations in the large intestine allowing small animals to eat fibrous foods. In The Digestive System in Mammals: Food Form and Function; Chivers, D.J., Langer, P., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 287–310. [Google Scholar]
- Holtenius, K.; Björnhag, G. The colonic separation mechanism in the guinea-pig (Cavia porcellus) and the chinchilla (Chinchilla laniger). Comp. Biochem. Physiol. A Comp. Physiol. 1985, 82, 537–542. [Google Scholar] [CrossRef]
- Lange, K.; Buerger, M.; Stallmach, A.; Bruns, T. Effects of Antibiotics on Gut Microbiota. Dig. Dis. 2016, 34, 260–268. [Google Scholar] [CrossRef]
- O’ Donnell, M.M.; Harris, H.M.B.; Ross, R.P.; O’Toole, P.W. Core fecal microbiota of domesticated herbivorous ruminant, hindgut fermenters, and monogastric animals. MicrobiologyOpen 2017, 6, e00509. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Li, J.; Yang, L.; Liu, H.; Zhu, Y.; Ren, H.; Yu, F.; Liu, B. Short Term Effect of Ivermectin on the Bacterial Microbiota from Fecal Samples in Chinchillas (Chinchilla lanigera). Vet. Sci. 2023, 10, 169. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.; Moore, R.J.; Wong, C.H.Y. An insight into intestinal mucosal microbiota disruption after stroke. Sci. Rep. 2018, 8, 568. [Google Scholar] [CrossRef] [PubMed]
- Cotozzolo, E.; Cremonesi, P.; Curone, G.; Menchetti, L.; Riva, F.; Biscarini, F.; Marongiu, M.L.; Castrica, M.; Castiglioni, B.; Miraglia, D.; et al. Characterization of Bacterial Microbiota Composition along the Gastrointestinal Tract in Rabbits. Animals 2020, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Morinaga, K.; Kusada, H.; Tamaki, H. Bile Salt Hydrolases with Extended Substrate Specificity Confer a High Level of Resistance to Bile Toxicity on Atopobiaceae Bacteria. Int. J. Mol. Sci. 2022, 23, 10980. [Google Scholar] [CrossRef] [PubMed]
- Jing, C.; Wang, J.; Xie, Y.; Zhang, J.; Guo, Y.; Tian, T.; Tang, J.; Ju, F.; Wang, C.; Liu, Y.; et al. Investigation of the growth performance, blood status, gut microbiome and metabolites of rabbit fed with low-nicotine tobacco. Front. Microbiol. 2022, 13, 1026680. [Google Scholar] [CrossRef] [PubMed]
- Leth, M.L.; Pichler, M.J.; Abou Hachem, M. Butyrate-producing colonic clostridia: Picky glycan utilization specialists. Essays Biochem. 2023, 67, 415–428. [Google Scholar] [CrossRef]
- Raimondi, S.; Musmeci, E.; Candeliere, F.; Amaretti, A.; Rossi, M. Identification of mucin degraders of the human gut microbiota. Sci. Rep. 2021, 11, 11094. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Zhang, Y.; Shi, B.; Wu, X.; Zhao, H.; Xin, Z.; Yang, J. Benzoic Acid Metabolism and Lipopolysaccharide Synthesis of Intestinal Microbiome Affects the Health of Ruminants under Free-Range and Captive Mode. Life 2022, 12, 1071. [Google Scholar] [CrossRef]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Liang, N.; Zhang, X.; Han, C.; Nan, X. Functional differentiation related to decomposing complex carbohydrates of intestinal microbes between two wild zokor species based on 16SrRNA sequences. BMC Vet. Res. 2021, 17, 216. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhao, Y.; Wang, Y.; Geng, R.; Fang, J.; Kang, S.G.; Huang, K.; Tong, T. Eugenol, A Major Component of Clove Oil, Attenuates Adiposity, and Modulates Gut Microbiota in High-Fat Diet-Fed Mice. Mol. Nutr. Food Res. 2022, 66, e2200387. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.Z.; Liang, Y.D.; Ma, Q.Y.; Hao, W.Z.; Li, X.J.; Wu, M.S.; Deng, L.J.; Li, Y.M.; Chen, J.X. Xiaoyaosan improves depressive-like behavior in rats with chronic immobilization stress through modulation of the gut microbiota. Biomed. Pharmacother. 2019, 112, 108621. [Google Scholar] [CrossRef]
- Zhang, H.; Lang, X.; Zhang, Y.; Wang, C. Distribution of bacteria in different regions of the small intestine with Zanthoxylum bungeanum essential oil supplement in small-tailed Han sheep. Front. Microbiol. 2022, 13, 1062077. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Mei, X.; Lei, C.; Li, C.; Gao, Y.; Kong, L.; Zhai, X.; Wang, H. Enrofloxacin Shifts Intestinal Microbiota and Metabolic Profiling and Hinders Recovery from Salmonella enterica subsp. enterica Serovar Typhimurium Infection in Neonatal Chickens. mSphere 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ma, L.; Li, Z.; Yin, J.; Tan, B.; Chen, J.; Jiang, Q.; Ma, X. Evolution of the Gut Microbiota and Its Fermentation Characteristics of Ningxiang Pigs at the Young Stage. Animals 2021, 11, 638. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, F.; Yin, Y.; Huang, P.; Jiang, Q.; Liu, Z.; Yin, Y.; Chen, J. Dietary Litsea cubeba essential oil supplementation improves growth performance and intestinal health of weaned piglets. Anim. Nutr. 2023, 13, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Song, Z.; Ji, R.; Liu, Y.; Zhao, H.; Liu, L.; Li, F. Chlorogenic acid improves growth performance of weaned rabbits via modulating the intestinal epithelium functions and intestinal microbiota. Front. Microbiol. 2022, 13, 1027101. [Google Scholar] [CrossRef] [PubMed]
- Nardone, G.; Compare, D. The human gastric microbiota: Is. it time to rethink the pathogenesis of stomach diseases? United Eur. Gastroenterol. J. 2015, 3, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Erawijantari, P.P.; Mizutani, S.; Shiroma, H.; Shiba, S.; Nakajima, T.; Sakamoto, T.; Saito, Y.; Fukuda, S.; Yachida, S.; Yamada, T. Influence of gastrectomy for gastric cancer treatment on faecal microbiome and metabolome profiles. Gut 2020, 69, 1404–1415. [Google Scholar] [CrossRef] [PubMed]
- Seekatz, A.M.; Schnizlein, M.K.; Koenigsknecht, M.J.; Baker, J.R.; Hasler, W.L.; Bleske, B.E.; Young, V.B.; Sun, D. Spatial and Temporal Analysis of the Stomach and Small-Intestinal Microbiota in Fasted Healthy Humans. mSphere 2019, 4, e00126-19. [Google Scholar] [CrossRef] [PubMed]
- Haenni, M.; Lupo, A.; Madec, J.Y. Antimicrobial Resistance in Streptococcus spp. Microbiol. Spectr. 2018, 6, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Yang, J.; Jin, M.; Zhou, B.; Shi, L.; Zhao, L.; Zhang, J.; Lin, Z.; Ren, J.; Liu, L.; et al. Fecal Streptococcus Alteration Is Associated with Gastric Cancer Occurrence and Liver Metastasis. mBio 2021, 12, e0299421. [Google Scholar] [CrossRef] [PubMed]
- Zoetendal, E.G.; Raes, J.; van den Bogert, B.; Arumugam, M.; Booijink, C.C.; Troost, F.J.; Bork, P.; Wels, M.; de Vos, W.M.; Kleerebezem, M. The human small intestinal microbiota is driven by rapid uptake and conversion of simple carbohydrates. ISME J. 2012, 6, 1415–1426. [Google Scholar] [CrossRef] [PubMed]
- Suksong, W.; Kongjan, P.; Prasertsan, P.S.O.T. Thermotolerant cellulolytic Clostridiaceae and Lachnospiraceae rich consortium enhanced biogas production from oil palm empty fruit bunches by solid-state anaerobic digestion. Bioresour. Technol. 2019, 291, 121851. [Google Scholar] [CrossRef]
- Ericsson, A.C.; Johnson, P.J.; Lopes, M.A.; Perry, S.C.; Lanter, H.R. A Microbiological Map of the Healthy Equine Gastrointestinal Tract. PLoS ONE 2016, 11, e0166523. [Google Scholar] [CrossRef]
- O’Callaghan, J.; O’Toole, P.W. Lactobacillus: Host-microbe relationships. Curr. Top. Microbiol. Immunol. 2013, 358, 119–154. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Liu, B.; Ma, X.; Yang, L.; Lu, X.; Wang, W.; Li, J. The Microbiota Architecture of the Chinchilla Gastrointestinal Tract. Vet. Sci. 2024, 11, 58. https://doi.org/10.3390/vetsci11020058
Wu Y, Liu B, Ma X, Yang L, Lu X, Wang W, Li J. The Microbiota Architecture of the Chinchilla Gastrointestinal Tract. Veterinary Sciences. 2024; 11(2):58. https://doi.org/10.3390/vetsci11020058
Chicago/Turabian StyleWu, Yuhong, Bo Liu, Xinyi Ma, Luo Yang, Xinyi Lu, Wei Wang, and Jing Li. 2024. "The Microbiota Architecture of the Chinchilla Gastrointestinal Tract" Veterinary Sciences 11, no. 2: 58. https://doi.org/10.3390/vetsci11020058
APA StyleWu, Y., Liu, B., Ma, X., Yang, L., Lu, X., Wang, W., & Li, J. (2024). The Microbiota Architecture of the Chinchilla Gastrointestinal Tract. Veterinary Sciences, 11(2), 58. https://doi.org/10.3390/vetsci11020058