
Fosfomycin Resistance in Bacteria Isolated from Companion Animals (Dogs and Cats)





Abstract
Simple Summary
Abstract
1. Introduction
1.1. Properties and Mode of Action of Fosfomycin
1.2. Usage in Human and Veterinary Medicine
1.3. Fosfomycin Resistance
1.4. Objectives of the Review
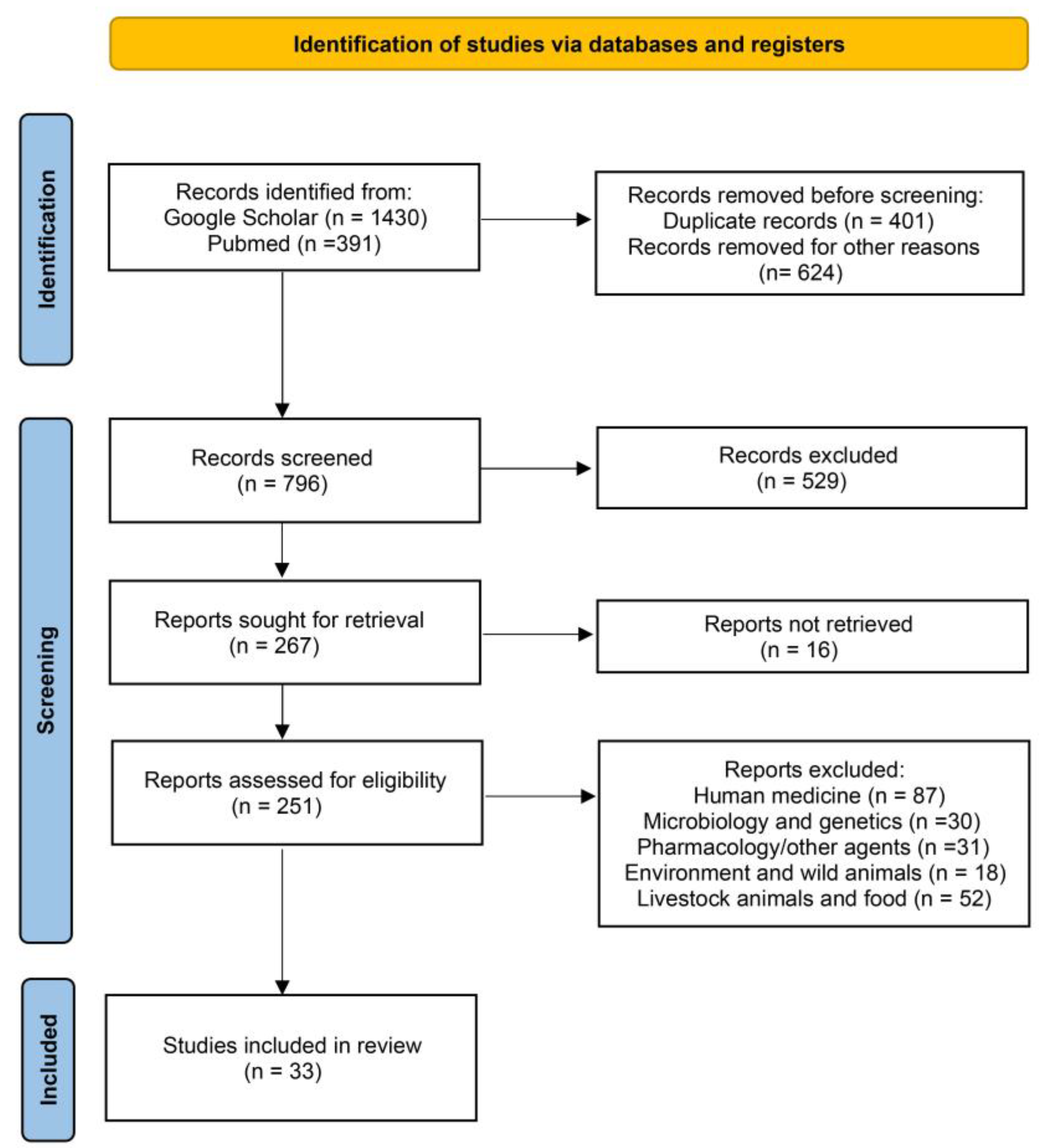
2. Materials and Methods
3. Results
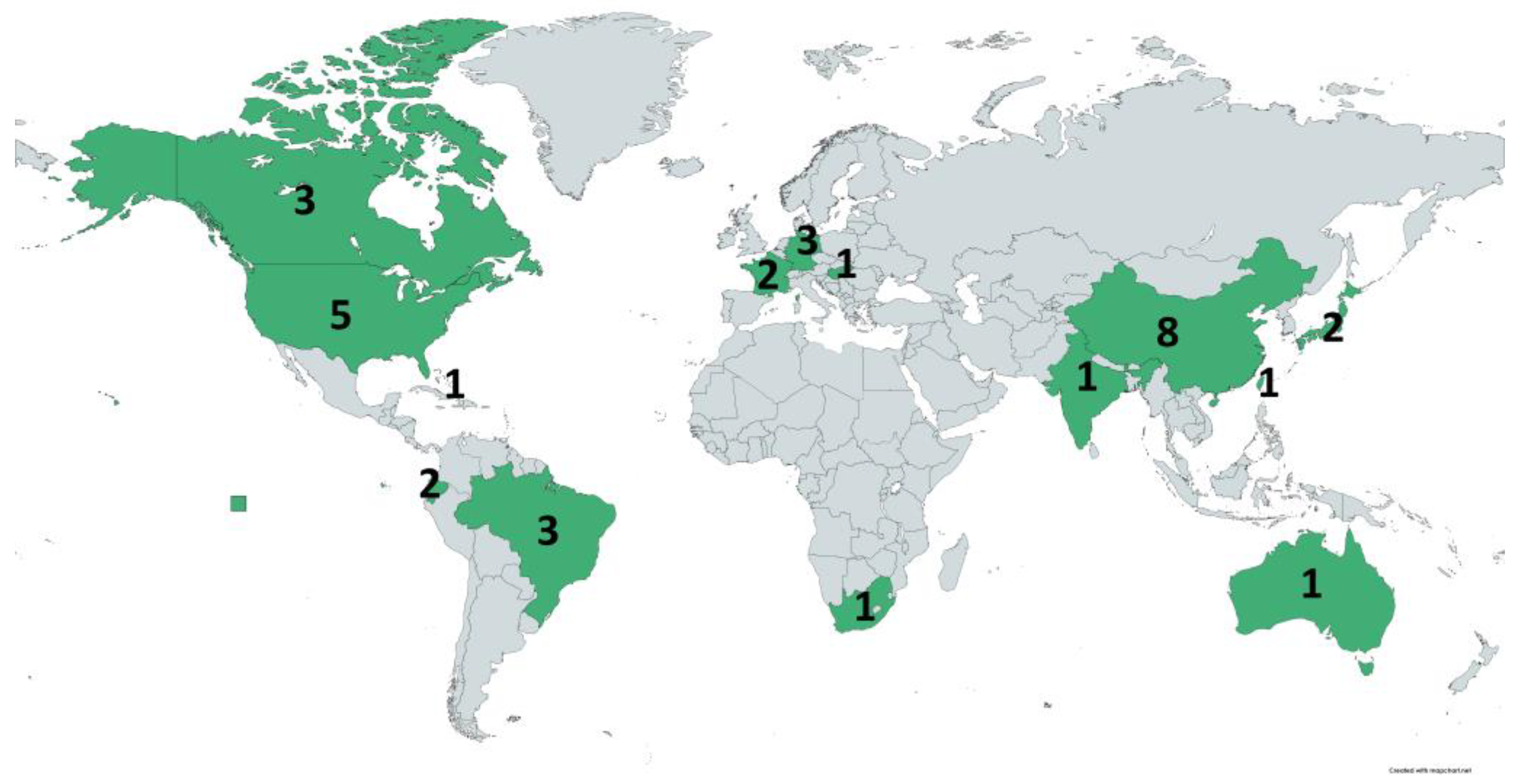
3.1. Geographical Distribution
3.2. Relevant Findings of the Studies Per Country of Isolates Origin
3.2.1. China
3.2.2. USA
3.2.3. Canada
3.2.4. USA and Canada
3.2.5. Germany
3.2.6. Brazil
3.2.7. France
3.2.8. Japan
3.2.9. Ecuador
3.2.10. India
3.2.11. Australia
3.2.12. Taiwan
3.2.13. Hungary
3.2.14. Caribbean
3.2.15. South Africa
3.3. Microorganisms and ARGs of Interest
3.4. Resistome and Phenotypic Resistance against Other Antibacterial Agents in Fosfomycin-Resistant Strains
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hendlin, D.; Stapley, E.O.; Jackson, M.; Wallick, H.; Miller, A.K.; Wolf, F.J.; Miller, T.W.; Chaiet, L.; Kahan, F.M.; Foltz, E.L.; et al. Phosphonomycin, a New Antibiotic Produced by Strains of Streptomyces. Science 1969, 166, 122–123. [Google Scholar] [CrossRef] [PubMed]
- Zhanel, G.G.; Walkty, A.J.; Karlowsky, J.A. Fosfomycin: A First-Line Oral Therapy for Acute Uncomplicated Cystitis. Can. J. Infect. Dis. Med. Microbiol. 2016, 2016, 2082693. [Google Scholar] [CrossRef]
- Falagas, M.E.; Vouloumanou, E.K.; Samonis, G.; Vardakas, K.Z. Fosfomycin. Clin. Microbiol. Rev. 2016, 29, 321–347. [Google Scholar] [CrossRef] [PubMed]
- Díez-Aguilar, M.; Cantón, R. New microbiological aspects of fosfomycin. Rev. Esp. Quimioter. 2019, 32 (Suppl. S1), 8–18. [Google Scholar] [PubMed]
- Silver, L.L. Fosfomycin: Mechanism and Resistance. Cold Spring Harb. Perspect. Med. 2017, 7, a025262. [Google Scholar] [CrossRef]
- Dijkmans, A.C.; Zacarías, N.V.O.; Burggraaf, J.; Mouton, J.W.; Wilms, E.; van Nieuwkoop, C.; Touw, D.J.; Stevens, J.; Kamerling, I.M.C. Fosfomycin: Pharmacological, Clinical and Future Perspectives. Antibiotics 2017, 6, 24. [Google Scholar] [CrossRef]
- Parker, S.; Lipman, J.; Koulenti, D.; Dimopoulos, G.; Roberts, J.A. What Is the Relevance of Fosfomycin Pharmacokinetics in the Treatment of Serious Infections in Critically Ill Patients? A Systematic Review. Int. J. Antimicrob. Agents 2013, 42, 289–293. [Google Scholar] [CrossRef]
- Bassetti, M.; Graziano, E.; Berruti, M.; Giacobbe, D.R. The Role of Fosfomycin for Multidrug-Resistant Gram-Negative Infections. Curr. Opin. Infect. Dis. 2019, 32, 617–625. [Google Scholar] [CrossRef]
- Candel, F.J.; Cantón, R. Uso Actual de La Fosfomicina: Del Laboratorio a La Práctica Clínica. Enfermedades Infecc. Microbiol. Clínica 2019, 37, 1–3. [Google Scholar] [CrossRef]
- Michalopoulos, A.S.; Livaditis, I.G.; Gougoutas, V. The Revival of Fosfomycin. Int. J. Infect. Dis. 2011, 15, e732–e739. [Google Scholar] [CrossRef]
- Karaiskos, I.; Giamarellou, H. Multidrug-Resistant and Extensively Drug-Resistant Gram-Negative Pathogens: Current and Emerging Therapeutic Approaches. Expert Opin. Pharmacother. 2014, 15, 1351–1370. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Bulman, Z.P.; Lenhard, J.R.; Satlin, M.J.; Kreiswirth, B.N.; Walsh, T.J.; Marrocco, A.; Bergen, P.J.; Nation, R.L.; Li, J.; et al. Pharmacodynamics of Colistin and Fosfomycin: A ‘Treasure Trove’ Combination Combats KPC-Producing Klebsiella Pneumoniae. J. Antimicrob. Chemother. 2017, 72, 1985–1990. [Google Scholar] [CrossRef] [PubMed]
- Ku, N.S.; Lee, S.-H.; Lim, Y.-S.; Choi, H.; Ahn, J.Y.; Jeong, S.J.; Shin, S.J.; Choi, J.Y.; Choi, Y.H.; Yeom, J.-S.; et al. In Vivo Efficacy of Combination of Colistin with Fosfomycin or Minocycline in a Mouse Model of Multidrug-Resistant Acinetobacter Baumannii Pneumonia. Sci. Rep. 2019, 9, 17127. [Google Scholar] [CrossRef] [PubMed]
- Saelim, W.; Changpradub, D.; Thunyaharn, S.; Juntanawiwat, P.; Nulsopapon, P.; Santimaleeworagun, W. Colistin plus Sulbactam or Fosfomycin against Carbapenem-Resistant Acinetobacter Baumannii: Improved Efficacy or Decreased Risk of Nephrotoxicity? Infect. Chemother. 2021, 53, 128. [Google Scholar] [CrossRef]
- Singkham-in, U.; Chatsuwan, T. In Vitro Activities of Carbapenems in Combination with Amikacin, Colistin, or Fosfomycin against Carbapenem-Resistant Acinetobacter Baumannii Clinical Isolates. Diagn. Microbiol. Infect. Dis. 2018, 91, 169–174. [Google Scholar] [CrossRef]
- Hall Snyder, A.D.; Werth, B.J.; Nonejuie, P.; McRoberts, J.P.; Pogliano, J.; Sakoulas, G.; Yim, J.; Singh, N.; Rybak, M.J. Fosfomycin Enhances the Activity of Daptomycin against Vancomycin-Resistant Enterococci in an In Vitro Pharmacokinetic-Pharmacodynamic Model. Antimicrob. Agents Chemother. 2016, 60, 5716–5723. [Google Scholar] [CrossRef]
- Yan, Y.; Yang, G.; Li, Y.; Mao, J.; Wang, S.; Zhang, N.; Liu, H.; Huang, X. Factorial Design and Post-Antibiotic Sub-MIC Effects of Linezolid Combined with Fosfomycin against Vancomycin-Resistant Enterococci. Ann. Transl. Med. 2022, 10, 148. [Google Scholar] [CrossRef]
- Pérez, D.; Tapia, M.; Soraci, A. Fosfomycin: Uses and Potentialities in Veterinary Medicine. Open Vet. J. 2014, 5, 26. [Google Scholar] [CrossRef]
- Harada, K.; Shimizu, T.; Kawaguchi, K.; Furuhashi, T.; Ishihara, G. Urinary Pharmacokinetic and Pharmacodynamic Profiles of Fosfomycin against Extended-Spectrum β-Lactamase-Producing Escherichia coli with Canine Ex Vivo Modeling: A Pilot Study. Antibiotics 2020, 9, 230. [Google Scholar] [CrossRef]
- European Medicines Committee. Categorisation of Antibiotics Used in Animals Promotes Responsible Use to Protect Public and Animal Health; European Medicines Agency: Amsterdam, The Netherlands, 2020. [Google Scholar]
- World Health Organization. Critically Important Antimicrobials for Human Medicine, 6th ed.; World Health Organization: Geneva, Switzerland, 2019; ISBN 978-92-4-151552-8. [Google Scholar]
- Aramayona, J.J.; Bregante, M.A.; Solans, C.; Rueda, S.; Fraile, L.J.; Garcia, M.A. Pharmacokinetics of fosfomycin in chickens after a single intravenous dose and tissue levels following chronic oral administration. Vet Res. 1997, 28, 581–588. [Google Scholar]
- Soraci, A.L.; Pérez, D.S.; Tapia, M.O.; Martinez, G.; Dieguez, S.N.; Buronfosse-Roque, F.; Colusi, A.; Romano, O. Pharmacocinétique et biodisponibilité de fosfomycine chez le poulet de chair. Ec. Natl. Vet. Toulouse Rev. Med. Vet. 2011, 162, 358–363. [Google Scholar]
- Gutierrez, O.L.; Ocampo, C.L.; Aguilera, J.R.; Luna, J.; Sumano, L.H. Pharmacokinetics of Disodium-Fosfomycin in Mongrel Dogs. Res. Vet. Sci. 2008, 85, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Soraci, A.L.; Pérez, D.S.; Martínez, G.; Amanto, F.; Tapia, M.O.; Dieguez, S.; Fernández Paggi, M.B. Fosfomycin Concentrations in Epithelial Lining Fluid in Weaning Piglets: Fosfomycin Concentrations in ELF. J. Vet. Pharmacol. Ther. 2012, 35, 406–409. [Google Scholar] [CrossRef] [PubMed]
- Prez, D.; Soraci, A.; Tapia, M. Pharmacokinetics and Bioavailability of Calcium Fosfomycin in Post Weaning Piglets after Oral Administration. Int. J. Agro Vet. Med. Sci. 2012, 6, 424. [Google Scholar] [CrossRef]
- Sumano, L.H.; Ocampo, C.L.; Gutierrez, O.L. Intravenous and Intramuscular Pharmacokinetics of a Single-Daily Dose of Disodium Fosfomycin in Cattle, Administered for 3 days. J. Vet. Pharmacol. Ther. 2007, 30, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Zozaya, D.H.; Gutirrez, O.L.; Ocampo, C.L.; Sumano, L.H. Pharmacokinetics of a Single Bolus Intravenous, Intramuscular and Subcutaneous Dose of Disodium Fosfomycin in Horses. J. Vet. Pharmacol. Ther. 2008, 31, 321–327. [Google Scholar] [CrossRef]
- Fukata, T.; Imai, N.; Shibata, S. Acute Renal Insufficiency in Cats after Fosfomycin Administration. Vet. Rec. 2008, 163, 337–338. [Google Scholar] [CrossRef]
- Sakai, Y.; Yoshida, T.; Shibutani, M.; Ohmori, K. Acute Renal Failure in an Adult Cat Following Oral Administration of Fosfomycin. J. Feline Med. Surg. Open Rep. 2018, 4, 205511691878660. [Google Scholar] [CrossRef]
- Castañeda-García, A.; Blázquez, J.; Rodríguez-Rojas, A. Molecular Mechanisms and Clinical Impact of Acquired and Intrinsic Fosfomycin Resistance. Antibiotics 2013, 2, 217–236. [Google Scholar] [CrossRef]
- Couce, A.; Briales, A.; Rodríguez-Rojas, A.; Costas, C.; Pascual, Á.; Blázquez, J. Genomewide Overexpression Screen for Fosfomycin Resistance in Escherichia Coli: MurA Confers Clinical Resistance at Low Fitness Cost. Antimicrob. Agents Chemother. 2012, 56, 2767–2769. [Google Scholar] [CrossRef]
- Xu, S.; Fu, Z.; Zhou, Y.; Liu, Y.; Xu, X.; Wang, M. Mutations of the Transporter Proteins GlpT and UhpT Confer Fosfomycin Resistance in Staphylococcus Aureus. Front. Microbiol. 2017, 8, 914. [Google Scholar] [CrossRef] [PubMed]
- Leite, G.C.; Perdigão-Neto, L.V.; Ruedas Martins, R.C.; Rizek, C.; Levin, A.S.; Costa, S.F. Genetic Factors Involved in Fosfomycin Resistance of Multidrug-Resistant Acinetobacter Baumannii. Infect. Genet. Evol. 2021, 93, 104943. [Google Scholar] [CrossRef] [PubMed]
- Ito, R.; Mustapha, M.M.; Tomich, A.D.; Callaghan, J.D.; McElheny, C.L.; Mettus, R.T.; Shanks, R.M.Q.; Sluis-Cremer, N.; Doi, Y. Widespread Fosfomycin Resistance in Gram-Negative Bacteria Attributable to the Chromosomal FosA Gene. mBio 2017, 8, e00749-17. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Athanasaki, F.; Voulgaris, G.L.; Triarides, N.A.; Vardakas, K.Z. Resistance to Fosfomycin: Mechanisms, Frequency and Clinical Consequences. Int. J. Antimicrob. Agents 2019, 53, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Vuillemin, X.; Kieffer, N.; Mueller, L.; Descombes, M.-C.; Nordmann, P. Identification of FosA8, a Plasmid-Encoded Fosfomycin Resistance Determinant from Escherichia Coli, and Its Origin in Leclercia Adecarboxylata. Antimicrob. Agents Chemother. 2019, 63, e01403-19. [Google Scholar] [CrossRef]
- ten Doesschate, T.; Abbott, I.J.; Willems, R.J.L.; Top, J.; Rogers, M.R.C.; Bonten, M.M.; Paganelli, F.L. In Vivo Acquisition of Fosfomycin Resistance in Escherichia Coli by FosA Transmission from Commensal Flora. J. Antimicrob. Chemother. 2019, 74, 3630–3632. [Google Scholar] [CrossRef]
- Huang, Y.; Lin, Q.; Zhou, Q.; Lv, L.; Wan, M.; Gao, X.; Wang, C.; Liu, J.-H. Identification of FosA10, a Novel Plasmid-Mediated Fosfomycin Resistance Gene of Klebsiella Pneumoniae Origin, in Escherichia Coli. Infect. Drug Resist. 2020, 13, 1273–1279. [Google Scholar] [CrossRef]
- Gaballa, A.; Newton, G.L.; Antelmann, H.; Parsonage, D.; Upton, H.; Rawat, M.; Claiborne, A.; Fahey, R.C.; Helmann, J.D. Biosynthesis and Functions of Bacillithiol, a Major Low-Molecular-Weight Thiol in Bacilli. Proc. Natl. Acad. Sci. USA 2010, 107, 6482–6486. [Google Scholar] [CrossRef]
- Cao, M.; Bernat, B.A.; Wang, Z.; Armstrong, R.N.; Helmann, J.D. FosB, a Cysteine-Dependent Fosfomycin Resistance Protein under the Control of ς W, an Extracytoplasmic-Function ς Factor in Bacillus Subtilis. J. Bacteriol. 2001, 183, 2380–2383. [Google Scholar] [CrossRef]
- Fillgrove, K.L.; Pakhomova, S.; Schaab, M.R.; Newcomer, M.E.; Armstrong, R.N. Structure and Mechanism of the Genomically Encoded Fosfomycin Resistance Protein, FosX, from Listeria Monocytogenes. Biochemistry 2007, 46, 8110–8120. [Google Scholar] [CrossRef]
- Kobayashi, S.; Kuzuyama, T.; Seto, H. Characterization of the FomA and FomB Gene Products from Streptomyces Wedmorensis, Which Confer Fosfomycin Resistance on Escherichia Coli. Antimicrob. Agents Chemother. 2000, 44, 647–650. [Google Scholar] [CrossRef] [PubMed]
- García, P.; Arca, P.; Evaristo Suárez, J. Product of FosC, a Gene from Pseudomonas Syringae, Mediates Fosfomycin Resistance by Using ATP as Cosubstrate. Antimicrob. Agents Chemother. 1995, 39, 1569–1573. [Google Scholar] [CrossRef] [PubMed]
- Truong-Bolduc, Q.C.; Wang, Y.; Hooper, D.C. Tet38 Efflux Pump Contributes to Fosfomycin Resistance in Staphylococcus Aureus. Antimicrob. Agents Chemother. 2018, 62, e00927-18. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sharma, R.; Bhattacharyya, T.; Bhando, T.; Pathania, R. Fosfomycin Resistance in Acinetobacter Baumannii Is Mediated by Efflux through a Major Facilitator Superfamily (MFS) Transporter—AbaF. J. Antimicrob. Chemother. 2017, 72, 68–74. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Hubka, P.; Boothe, D.M. In Vitro Susceptibility of Canine and Feline Escherichia Coli to Fosfomycin. Vet. Microbiol. 2011, 149, 277–282. [Google Scholar] [CrossRef]
- Haenni, M.; Saras, E. A USA300 Variant and Other Human-Related Methicillin-Resistant Staphylococcus Aureus Strains Infecting Cats and Dogs in France. J. Antimicrob. Chemother. 2012, 67, 326–329. [Google Scholar] [CrossRef]
- Hou, J.; Huang, X.; Deng, Y.; He, L.; Yang, T.; Zeng, Z.; Chen, Z.; Liu, J.-H. Dissemination of the Fosfomycin Resistance Gene FosA3 with CTX-M β-Lactamase Genes and RmtB Carried on IncFII Plasmids among Escherichia Coli Isolates from Pets in China. Antimicrob. Agents Chemother. 2012, 56, 2135–2138. [Google Scholar] [CrossRef]
- He, L.; Partridge, S.R.; Yang, X.; Hou, J.; Deng, Y.; Yao, Q.; Zeng, Z.; Chen, Z.; Liu, J.-H. Complete Nucleotide Sequence of PHN7A8, an F33:A-:B-Type Epidemic Plasmid Carrying BlaCTX-M-65, FosA3 and RmtB from China. J. Antimicrob. Chemother. 2013, 68, 46–50. [Google Scholar] [CrossRef]
- Ho, P.L.; Chan, J.; Lo, W.U.; Law, P.Y.; Li, Z.; Lai, E.L.; Chow, K.H. Dissemination of Plasmid-Mediated Fosfomycin Resistance FosA3 among Multidrug-Resistant Escherichia Coli from Livestock and Other Animals. J. Appl. Microbiol. 2013, 114, 695–702. [Google Scholar] [CrossRef]
- DiCicco, M.; Weese, S.; Neethirajan, S.; Rousseau, J.; Singh, A. Fosfomycin Susceptibility of Canine Methicillin-Resistant Staphylococcus Pseudintermedius Isolates. Res. Vet. Sci. 2014, 96, 251–253. [Google Scholar] [CrossRef] [PubMed]
- Haenni, M.; Hocquet, D.; Ponsin, C.; Cholley, P.; Guyeux, C.; Madec, J.-Y.; Bertrand, X. Population Structure and Antimicrobial Susceptibility of Pseudomonas Aeruginosa from Animal Infections in France. BMC Vet. Res. 2015, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Ewers, C.; Klotz, P.; Leidner, U.; Stamm, I.; Prenger-Berninghoff, E.; Göttig, S.; Semmler, T.; Scheufen, S. OXA-23 and IS Aba1 –OXA-66 Class D β-Lactamases in Acinetobacter Baumannii Isolates from Companion Animals. Int. J. Antimicrob. Agents 2017, 49, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Wu, D.; Lei, L.; Shen, Z.; Wang, Y.; Liao, K. The Detection of Fosfomycin Resistance Genes in Enterobacteriaceae from Pets and Their Owners. Vet. Microbiol. 2016, 193, 67–71. [Google Scholar] [CrossRef]
- Yukawa, S.; Tsuyuki, Y.; Sato, T.; Fukuda, A.; Usui, M.; Tamura, Y. Antimicrobial Resistance of Pseudomonas Aeruginosa Isolated from Dogs and Cats in Primary Veterinary Hospitals in Japan. Jpn. J. Infect. Dis. 2017, 70, 461–463. [Google Scholar] [CrossRef]
- Maeyama, Y.; Taniguchi, Y.; Hayashi, W.; Ohsaki, Y.; Osaka, S.; Koide, S.; Tamai, K.; Nagano, Y.; Arakawa, Y.; Nagano, N. Prevalence of ESBL/AmpC Genes and Specific Clones among the Third-Generation Cephalosporin-Resistant Enterobacteriaceae from Canine and Feline Clinical Specimens in Japan. Vet. Microbiol. 2018, 216, 183–189. [Google Scholar] [CrossRef]
- Zhang, P.L.C.; Shen, X.; Chalmers, G.; Reid-Smith, R.J.; Slavic, D.; Dick, H.; Boerlin, P. Prevalence and Mechanisms of Extended-Spectrum Cephalosporin Resistance in Clinical and Fecal Enterobacteriaceae Isolates from Dogs in Ontario, Canada. Vet. Microbiol. 2018, 213, 82–88. [Google Scholar] [CrossRef]
- Kaspar, U.; von Lützau, A.; Schlattmann, A.; Roesler, U.; Köck, R.; Becker, K. Zoonotic Multidrug-Resistant Microorganisms among Small Companion Animals in Germany. PLoS ONE 2018, 13, e0208364. [Google Scholar] [CrossRef]
- Pruthvishree, B.S.; Kumar, O.R.V.; Sivakumar, M.; Tamta, S.; Sunitha, R.; Sinha, D.K.; Singh, B.R. Molecular Characterization of Extensively Drug Resistant (XDR), Extended Spectrum Beta-Lactamases (ESBL) and New Delhi Metallo Beta-Lactamase-1 (BlaNDM1) Producing Escherichia Coli Isolated from a Male Dog—A Case Report. Escherichia Coli. Vet. Arhiv. 2018, 88, 139–148. [Google Scholar] [CrossRef]
- Fernandes, M.R.; Sellera, F.P.; Moura, Q.; Carvalho, M.P.N.; Rosato, P.N.; Cerdeira, L.; Lincopan, N. Zooanthroponotic Transmission of Drug-Resistant Pseudomonas Aeruginosa, Brazil. Emerg. Infect. Dis. 2018, 24, 1160–1162. [Google Scholar] [CrossRef]
- Cui, L.; Lei, L.; Lv, Y.; Zhang, R.; Liu, X.; Li, M.; Zhang, F.; Wang, Y. Bla NDM-1 -Producing Multidrug-Resistant Escherichia Coli Isolated from a Companion Dog in China. J. Glob. Antimicrob. Resist. 2018, 13, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Paredes, D.; Haro, M.; Leoro-Garzón, P.; Barba, P.; Loaiza, K.; Mora, F.; Fors, M.; Vinueza-Burgos, C.; Fernández-Moreira, E. Multidrug-Resistant Escherichia Coli Isolated from Canine Faeces in a Public Park in Quito, Ecuador. J. Glob. Antimicrob. Resist. 2019, 18, 263–268. [Google Scholar] [CrossRef]
- Courtise, R.M. Characterisation of Antimicrobial Resistance among Canine Urinary Isolates in Western Canada. Master’s Thesis, Department of Veterinary Microbiology, University of Saskatchewan, Saskatoon, SK, Canada, 2018; pp. 110–136. [Google Scholar]
- Kamathewatta, K.I.; Bushell, R.N.; Young, N.D.; Stevenson, M.A.; Billman-Jacobe, H.; Browning, G.F.; Marenda, M.S. Exploration of Antibiotic Resistance Risks in a Veterinary Teaching Hospital with Oxford Nanopore Long Read Sequencing. PLoS ONE 2019, 14, e0217600. [Google Scholar] [CrossRef] [PubMed]
- Sartori, L.; Sellera, F.P.; Moura, Q.; Cardoso, B.; Cerdeira, L.; Lincopan, N. Multidrug-Resistant CTX-M-15-Positive Klebsiella Pneumoniae ST307 Causing Urinary Tract Infection in a Dog in Brazil. J. Glob. Antimicrob. Resist. 2019, 19, 96–97. [Google Scholar] [CrossRef] [PubMed]
- Sellera, F.P.; Lopes, R.; Monte, D.F.M.; Cardoso, B.; Esposito, F.; dos Anjos, C.; da Silva, L.C.B.A.; Lincopan, N. Genomic Analysis of Multidrug-Resistant CTX-M-15-Positive Klebsiella Pneumoniae Belonging to the Highly Successful ST15 Clone Isolated from a Dog with Chronic Otitis. J. Glob. Antimicrob. Resist. 2020, 22, 659–661. [Google Scholar] [CrossRef]
- Chen, L.; Ou, B.; Zhang, M.; Chou, C.-H.; Chang, S.-K.; Zhu, G. Coexistence of Fosfomycin Resistance Determinant FosA and FosA 3 in Enterobacter Cloacae Isolated from Pets with Urinary Tract Infection in Taiwan. Microb. Drug Resist. 2021, 27, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Loayza-Villa, F.; Salinas, L.; Tijet, N.; Villavicencio, F.; Tamayo, R.; Salas, S.; Rivera, R.; Villacis, J.; Satan, C.; Ushiña, L.; et al. Diverse Escherichia Coli Lineages from Domestic Animals Carrying Colistin Resistance Gene Mcr-1 in an Ecuadorian Household. J. Glob. Antimicrob. Resist. 2020, 22, 63–67. [Google Scholar] [CrossRef]
- Cummings, K.J.; Mitchell, P.K.; Rodriguez-Rivera, L.D.; Goodman, L.B. Sequence Analysis of Salmonella Enterica Isolates Obtained from Shelter Dogs throughout Texas. Vet. Med. Sci. 2020, 6, 975–979. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Lei, L.; Zhang, H.; Dai, H.; Song, Y.; Li, L.; Wang, Y.; Xia, Z. Molecular Investigation of Klebsiella Pneumoniae from Clinical Companion Animals in Beijing, China, 2017–2019. Pathogens 2021, 10, 271. [Google Scholar] [CrossRef]
- Qiu, Y.; Nambiar, R.B.; Xu, X.; Weng, S.; Pan, H.; Zheng, K.; Yue, M. Global Genomic Characterization of Salmonella Enterica Serovar Telelkebir. Front. Microbiol. 2021, 12, 704152. [Google Scholar] [CrossRef]
- Sahin-Tóth, J.; Kovács, E.; Tóthpál, A.; Juhász, J.; Forró, B.; Bányai, K.; Havril, K.; Horváth, A.; Ghidán, Á.; Dobay, O. Whole Genome Sequencing of Coagulase Positive Staphylococci from a Dog-and-Owner Screening Survey. PLoS ONE 2021, 16, e0245351. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sun, Z.-Z.; Rong, J.-C.; Xie, B.-B. Comparative Genomics Reveals Broad Genetic Diversity, Extensive Recombination and Nascent Ecological Adaptation in Micrococcus Luteus. BMC Genom. 2021, 22, 124. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Chen, W.; Zhou, R.; Yang, J.; Wu, Y.; Zheng, J.; Fei, S.; Wu, G.; Sun, Z.; Li, J.; et al. Characteristics of the Plasmid-Mediated Colistin-Resistance Gene Mcr-1 in Escherichia Coli Isolated from a Veterinary Hospital in Shanghai. Front. Microbiol. 2022, 13, 1002827. [Google Scholar] [CrossRef] [PubMed]
- Butaye, P.; Stegger, M.; Moodley, A.; Damborg, P.; Williams, A.; Halliday-Simmonds, I.; Guardabassi, L. One Health Genomic Study of Human and Animal Klebsiella Pneumoniae Isolated at Diagnostic Laboratories on a Small Caribbean Island. Antibiotics 2021, 11, 42. [Google Scholar] [CrossRef]
- Jonker, A.; Gouws, J.; Kapp, E.R.; Henning, A. Isolation of a Multidrug-Resistant Escherichia Coli Pathotype Stx2:Cnf1:Cnf2:Eae as a Potential Cause of Hemorrhagic Diarrhea and Secondary Septicemia in a Dog. J. Vet. Diagn. Investig. 2022, 34, 339–342. [Google Scholar] [CrossRef]
- Bruce, S.A.; Smith, J.T.; Mydosh, J.L.; Ball, J.; Needle, D.B.; Gibson, R.; Andam, C.P. Shared Antibiotic Resistance and Virulence Genes in Staphylococcus Aureus from Diverse Animal Hosts. Sci. Rep. 2022, 12, 4413. [Google Scholar] [CrossRef]
- Teng, L.; Liao, S.; Zhou, X.; Jia, C.; Feng, M.; Pan, H.; Ma, Z.; Yue, M. Prevalence and Genomic Investigation of Multidrug-Resistant Salmonella Isolates from Companion Animals in Hangzhou, China. Antibiotics 2022, 11, 625. [Google Scholar] [CrossRef]
- Kim, D.-W.; Cha, C.-J. Antibiotic Resistome from the One-Health Perspective: Understanding and Controlling Antimicrobial Resistance Transmission. Exp. Mol. Med. 2021, 53, 301–309. [Google Scholar] [CrossRef]
- Jiang, Y.; Shen, P.; Wei, Z.; Liu, L.; He, F.; Shi, K.; Wang, Y.; Wang, H.; Yu, Y. Dissemination of a Clone Carrying a FosA3-Harbouring Plasmid Mediates High Fosfomycin Resistance Rate of KPC-Producing Klebsiella Pneumoniae in China. Int. J. Antimicrob. Agents 2015, 45, 66–70. [Google Scholar] [CrossRef]
- Mendes, A.C.; Rodrigues, C.; Pires, J.; Amorim, J.; Ramos, M.H.; Novais, Â.; Peixe, L. Importation of Fosfomycin Resistance FosA3 Gene to Europe. Emerg. Infect. Dis. 2016, 22, 346–348. [Google Scholar] [CrossRef]
- Stull, J.W.; Weese, J.S. Hospital-Associated Infections in Small Animal Practice. Vet. Clin. N. Am. Small Anim. Pract. 2015, 45, 217–233. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibillity Testing. Supplement M100, 32nd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022; ISBN 978-1-68440-134-5. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals. Supplement VET01S, 6th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2023; ISBN 978-1-68440-167-3. [Google Scholar]
- Schmidt, V.M.; Pinchbeck, G.; McIntyre, K.M.; Nuttall, T.; McEwan, N.; Dawson, S.; Williams, N.J. Routine Antibiotic Therapy in Dogs Increases the Detection of Antimicrobial-Resistant Faecal Escherichia Coli. J. Antimicrob. Chemother. 2018, 73, 3305–3316. [Google Scholar] [CrossRef] [PubMed]
- Menard, J.; Goggs, R.; Mitchell, P.; Yang, Y.; Robbins, S.; Franklin-Guild, R.J.; Thachil, A.J.; Altier, C.; Anderson, R.; Putzel, G.G.; et al. Effect of Antimicrobial Administration on Fecal Microbiota of Critically Ill Dogs: Dynamics of Antimicrobial Resistance over Time. Anim. Microbiome 2022, 4, 36. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Mechanism of Resistance | Description | Related Genes | References |
|---|---|---|---|
| Modification or overexpression of MurA gene | A modification of MurA gene, could alter amino acid sequence in Fosfomycin’s binding site, conferring resistance [28,29]. Furthermore, a resistant phenotype can also be achieved by increased synthesis of the enzyme through overexpression of the MurA gene. | MurA | [31,32] |
| Reduced permeability |
| Variable related genes | [31,33,34] |
| Fosfomycin modifying enzymes | FosA enzyme: a glutathione S-transferase that inactivates Fosfomycin by the addition of glutathione. Mn+2 and K+ are used as cofactors. Mostly found in Gram-negative bacteria such as Enterobacteriaceae and Pseudomonas. Several subtypes of fosA have been identified: fosA3, fosA4, fosA5, fosA6, fosA8, fosA9, fosA10, fosAEC and fosASH 1. | fosA, fosA3, fosA4, fosA5, fosA6, fosA8, fosA9, fosA10, fosAEC, fosASH | [35,36,37,38,39] |
| FosB enzyme: differs from fosA in being a Mg+2 dependent enzyme and using l-cysteine or possibly bacillithiol, as the physiologic thiol donor. Additionally, an extracytoplasmic sigma factor SigW, seems to be essential for its expression. It is routinely detected in Gram-positive bacteria, (Staphylococcus spp., Enterococcus spp., Bacillus subtilis). | fosB | [36,40,41] | |
| FosX enzyme: a Mn2+-dependent epoxide hydrolase, which catalyzes the addition of a water molecule to C1 position of Fosfomycin’s oxirane ring and as a result breaks it and inactivates its antibacterial properties. It can be regularly found in specific species, such as Listeria monocytogenes, Clostridium botulinum, and Brucella melitensis. | fosX | [31,42] | |
| FomA and fomB enzymes: kinases that modify Fosfomycin by phosphorylation and thus detoxify it intracellularly. They are encountered in Fosfomycin-producing bacteria, such as Streptomyces spp. | fomA, fomB | [43] | |
| FosC enzyme: a kinase that converts Fosfomycin to Fosfomycin monophosphate, conferring intrinsic resistance in Pseudomonas syringae, through the expression of a chromosomally encoded gene. | fosC | [44] | |
| Efflux pumps | Tet38 (when overexpressed) and AbaF pumps in Staphylococcus aureus and Acinetobacter baumannii, respectively, contribute to Fosfomycin resistance, possibly by acting as efflux transporters of the agent. | Tet38, abaF | [45,46] |
| Country/Area | Bacterial Species | ARGs | Sample Origin | Fosfomycin Resistant/Total | Date/ Period | Reference |
|---|---|---|---|---|---|---|
| USA | E. coli | NS 1 | Dogs, cats | 3/275 | 2008–2010 | [48] |
| France | MRSA | fosB | Dogs, cats | 19/23 (19 fosB) | 2006–2010 | [49] |
| China | E. coli | fosA3 | Dogs, cats | 33/323 (29 fosA3) | 2006–2010 | [50] |
| China | E. coli | fosA3 | Dog | 1/1 (1 fosA3) | 2008 | [51] |
| China | E. coli | fosA3 | Dogs, cats | 12/766 (11 fosA3) | 2008–2010 | [52] |
| USA, Canada | MRSP 2 | fosB | Dogs | 7/31 (27 fosB) | NR 3 | [53] |
| France | P. aeruginosa | NS | Dogs | 22/46 | 2008–2011 | [54] |
| Germany | Ac. baumannii | NR | Dogs, cats | 25/25 | 2000–2013 | [55] |
| China | Enterobacteriacae (E. coli, Pr. mirabilis, E. fergusonii, C. freundii E. aerogenes, Kl. oxytoca, Kl. pneumoniae | fosA3, fosA | Dogs, cats, pet owners | 19/171 (16 fosA3: 8 E. coli, 4 Pr. mirabilis, 3 E. fergusonii, 1 C. freundii and 3 fosA: 1 E. aerogenes, 1 Kl. oxytoca, 1 Kl. pneumoniae) | 2013 | [56] |
| Japan | P. aeruginosa | NR | Dogs, cats | 71/200 | NR | [57] |
| Japan | Enterobacteriacae (381 E. coli, 50 Kl. pneumoniae, 56 P. mirabilis) | fosA3 | Dog, cat | 3/487 (3 fosA3: 2 E. coli, 1 Kl. pneumoniae) | 2016 | [58] |
| Canada | Enterobacteriacae (Kl. pneumoniae, E. cloacae) | fosA | Dogs | 3/47 (3 fosA: 2 Kl. pneumoniae, 1 E. cloacae) | 2015–2016 | [59] |
| Germany | Staphylococcus cohnii subsp. Cohnii | NR | Dog | 1/1 | 2015–2016 | [60] |
| India | XDR 4 E. coli | NR | Dog | 1/1 | NR | [61] |
| Brazil | P. aeruginosa | fosA | Dog | 1/1 (1 fosA) | 2016 | [62] |
| China | E. coli | NR | Dog | 1/1 | 2013 | [63] |
| Ecuador | E. coli | NR | Dogs | 10/23 | 2017 | [64] |
| Canada | E. coli, St. pseudintermedius | NF 5 (Absence of fosA, fosA3, fosB, fosC2) | Dogs | 7/274, 4/113 | 2013–2016 | [65] |
| Australia | Enterobacteriaceae, P. aeruginosa | fosA | Small Animal Hospital environment | 65/656, 23/59 | 2016–2017 | [66] |
| Brazil | Kl. pneumoniae (susceptible in the AST) | fosA | Dog | 1/1 (1 fosA) | 2018 | [67] |
| Brazil | Kl. pneumoniae (susceptible in the AST) | fosA | Dog | 1/1 (1 fosA) | 2019 | [68] |
| Taiwan | Enterobacter cloacae | fosA3, fosA | Dogs, cats | 8/19 (8 fosA, 3 co-carried fosA3) | 2010–2013 | [69] |
| Ecuador | E. coli | fosA3 | Dog | 1/1 (1 fosA3) | 2016 | [70] |
| USA | Salmonella spp. | fosA7 (fosASH) 6 | Dogs | 2/27 (2 fosA7) | 2013–2014 | [71] |
| China | Kl. pneumoniae | fosA | Dogs, cats | 105/105 (105 fosA) | 2017–2019 | [72] |
| Germany (China) 7 | Salmonella enterica serovar Telelkebir | fosA7 (fosASH) 6 | Dog | 1/1 (1 fosA7) | 2007 | [73] |
| Hungary | Staphylococcus aureus | fosB, murA and glpT modification | Dog and owner | 4/27 (2 fosB, 2 murA and glpT) | NR | [74] |
| USA (China) 7 | Micrococcus luteus | murA | Dog | 1/1 (1 murA) | 2019 | [75] |
| China | E. coli (mcr-1) | fosA3 | Dogs, cats | 7/7 (7 fosA3) | 2021 | [76] |
| Caribbean | Klebsiella pneumoniae | fosA | Dogs, cats | 2/2 (2 fosA) | 2011–2018 | [77] |
| South Africa | E.coli [MIC:(R), DD:(S)] 8 | NR | Dog | 1/1 | NR | [78] |
| USA | Staphylococcus aureus | fosB | Dogs, cats | 42/53 (42 fosB) | 2017–2020 | [79] |
| China | Salmonella enterica serovar Dublin | fosX | Dogs | NR | 2018 | [80] |
| Microorganisms | Number of References | Fosfomycin-Resistant Isolates * | Related Fosfomycin ARGs ** |
|---|---|---|---|
| E. coli | 13 | 86 | fosA3 (58) |
| Kl. pneumoniae | 7 | 113 | fosA (112), fosA3 (1) |
| P. aeruginosa | 4 | 117 | fosA (24) |
| St. aureus | 3 | 65 | fosB (63), murA (2), glpT (2) |
| Salmonella spp. | 3 | 3 | fosASH (3), fosX |
| St. pseudintermedius | 2 | 11 | fosB (7) |
| Enterobacter cloacae | 2 | 9 | fosA (9), fosA3 (3) |
| E. aerogenes | 1 | 1 | fosA (1) |
| Kl. oxytoca | 1 | 1 | fosA (1) |
| E. fergusonii | 1 | 3 | fosA3 (3) |
| Pr. mirabilis | 1 | 4 | fosA3 (4) |
| C. freundii | 1 | 1 | fosA3 (1) |
| Micrococcus luteus | 1 | 1 | murA (1) |
| Ac. baumannii | 1 | 25 | NR |
| Staphylococcus cohnii | 1 | 1 | NR |
| Country /Area | Bacterial Species | Fosfomycin Related ARGs | Other ARGs with ≥50% Prevalence among Fos-Resistant Isolates 1 | Agents with ≥50% Resistance Rates among Fos-Resistant Isolates 1,2 | Reference |
|---|---|---|---|---|---|
| France | S. aureus | fosB | mecA, blaZ, aadD | ENR, ERY, FOX, KAN, LIN, PEN, SPI, TOB | [49] |
| China | E. coli | fosA3 | blaCTX-M, rmtB | AMK, CHL, CTX, GEN, TET | [50] |
| China | E. coli | fosA3 | blaCTX-M, rmtB | --- | [51] |
| China | E. coli | fosA3 | blaCTX-M | CHL, CIP, COT, GEN, NAL, TET | [52] |
| USA, Canada | S. pseudi-ntermedius | fosB | mecA | b-lactams 3 | [53] |
| Germany | A. baumannii | NR | --- | CXM, CFD, PIT, SAM | [55] |
| China | E. coli, Pr. mirabilis, E. fergusonii, C. freundii | fosA3 | blaCTX-M | AMK, AMP, FAZ, CHL, CIP, GEN, FFC, KAN | [56] |
| Japan | E. coli, Kl. pneumoniae | fosA3 | ESBL, pAmpC | --- | [58] |
| Canada | Kl. pneumoniae, E. cloacae | fosA | blaCTX-M-15, aac(3)-IIa, strA, strB, aac(6′)Ib-cr, blaOXA-1, blaSHV-83, blaTEM-1-B, qnrB1, sul2, drfA14, tetA | --- | [59] |
| Germany | Staphylococcus cohnii | NR | mecA | OXA | [60] |
| India | E. coli | NR | blaCTX-M, blaAmpCblaTEM, blaNDM-1, sul1 | AMK, AMC, AZT, CAZ, CFD, CFM, CHL, CIP, COL, CRO, CTX, CTC, CTR, ERT, FEP, FOX, GAT, GEN, IMP, MER, MOX, NOR, NIT, OFL, PMB, SXT, TET, TOB, VAN | [61] |
| Brazil | P. aeruginosa | fosA | blaVIM-2, blaPAO, blaOXA-4, blaOXA-50, aadA2, aac(3)-Id, aph(3)-IIb, catB7, cmlA1, sul1, dfrB5, tetG | AMK, AMC, CAZ, CIP, CHL, CRO, CTX, FEP, FOX, GEN, IMP, MER, NAL, PIT, STX, TET, TIC | [62] |
| China | E. coli | NR | blaNDM-1, drfA17, sul1, aadA5 | CAZ, CTX, CIP, ERT, GEN, IMP, MER, PIP, TET | [63] |
| Ecuador | E. coli | NR | blaCTX-M | AZT, CAZ, CHL, CIP, CTX, DOX, FEP, LEV, NAL, NOR, TET | [64] |
| Canada | S. pseudinter- medius | NF | mecA | PEN, OXA, AMP, CLI | [65] |
| Brazil | Kl. pneumoniae | fosA | blaCTX-M-15, blaSHV, blaOXA-1, aph(3”)-Ib, aph(6)-Id, aac (3)-IIa, tetA, aac(6′)-Ib-cr, qnrB1, oqxA and oqxB, dfrA | AMC, CAZ, CIP, CTX, ENR FEP, FUR, GEN, LEV, NOR, OFL, TET | [67] |
| Brazil | Kl. pneumoniae | fosA | blaCTX-M-15, blaSHV, blaOXA-1, 1, aadA2, aph(3′)-Ia], mphA, catB3, aac(6′)Ib-cr, oqxA, oqxB], sul1, tetA, dfrA12, GyrA, ParC | AMC, AZT, CIP, CRO, ENR, FEP, FUR, LEV, NAL, SXT, TET | [68] |
| Taiwan | Enterobacter cloacae | fosA3, fosA | bla TEM | AMP, SUD | [69] |
| Ecuador | E. coli | fosA3 | mcr-1.1, blaCTX-M-3, blaTEM-206, blaTEM-1B, tetA, GyrA, ParC | CIP, COL, CRO, FEP, SAM | [70] |
| China | Kl. pneumoniae | fosA | blaSHV, oqxA, oqxB, sul | AMC, DOX, FFC, SXT | [72] |
| Germany (China) | Salmonella enterica | fosA7 (fosASH) | aac(6′)-Iaa_1 | --- | [73] |
| Hungary | Staphylococcus aureus | fosB, murA, glpT | blaZ | PEN | [74] |
| China | E. coli | fosA3 | aac(3)-IVa, aph(3′)-IIa, aph(3′)-Ia, and aph(4)-Ia, blaCTX-M-65, blaTEM-1B, floR, drfA14, mcr-1, sul2, qnrS1, mdfA | COL, CTX, FFC | [76] |
| South Africa | E.coli | NR | --- | AMC, AMP, CEP, ENR, FUR, NEO, PEN | [78] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lysitsas, M.; Chatzipanagiotidou, I.; Billinis, C.; Valiakos, G. Fosfomycin Resistance in Bacteria Isolated from Companion Animals (Dogs and Cats). Vet. Sci. 2023, 10, 337. https://doi.org/10.3390/vetsci10050337
Lysitsas M, Chatzipanagiotidou I, Billinis C, Valiakos G. Fosfomycin Resistance in Bacteria Isolated from Companion Animals (Dogs and Cats). Veterinary Sciences. 2023; 10(5):337. https://doi.org/10.3390/vetsci10050337
Chicago/Turabian StyleLysitsas, Marios, Irene Chatzipanagiotidou, Charalambos Billinis, and George Valiakos. 2023. "Fosfomycin Resistance in Bacteria Isolated from Companion Animals (Dogs and Cats)" Veterinary Sciences 10, no. 5: 337. https://doi.org/10.3390/vetsci10050337
APA StyleLysitsas, M., Chatzipanagiotidou, I., Billinis, C., & Valiakos, G. (2023). Fosfomycin Resistance in Bacteria Isolated from Companion Animals (Dogs and Cats). Veterinary Sciences, 10(5), 337. https://doi.org/10.3390/vetsci10050337