
Review of the Developments of Bacterial Medium-Chain-Length Polyhydroxyalkanoates (mcl-PHAs)





Abstract
:1. Introduction
2. Mcl-PHA production
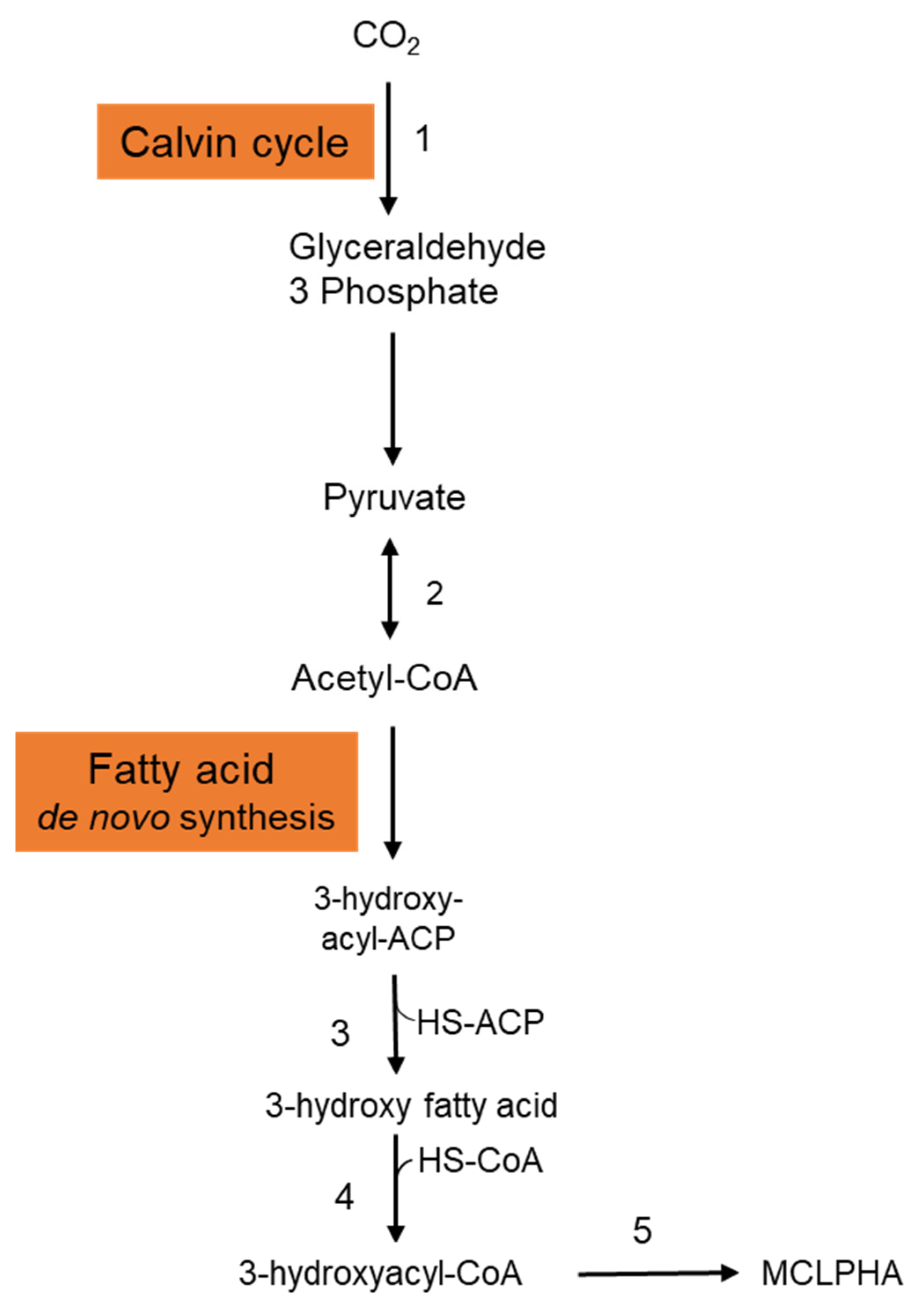
2.1. Inorganic Carbon Sources
2.2. Organic Carbon Sources
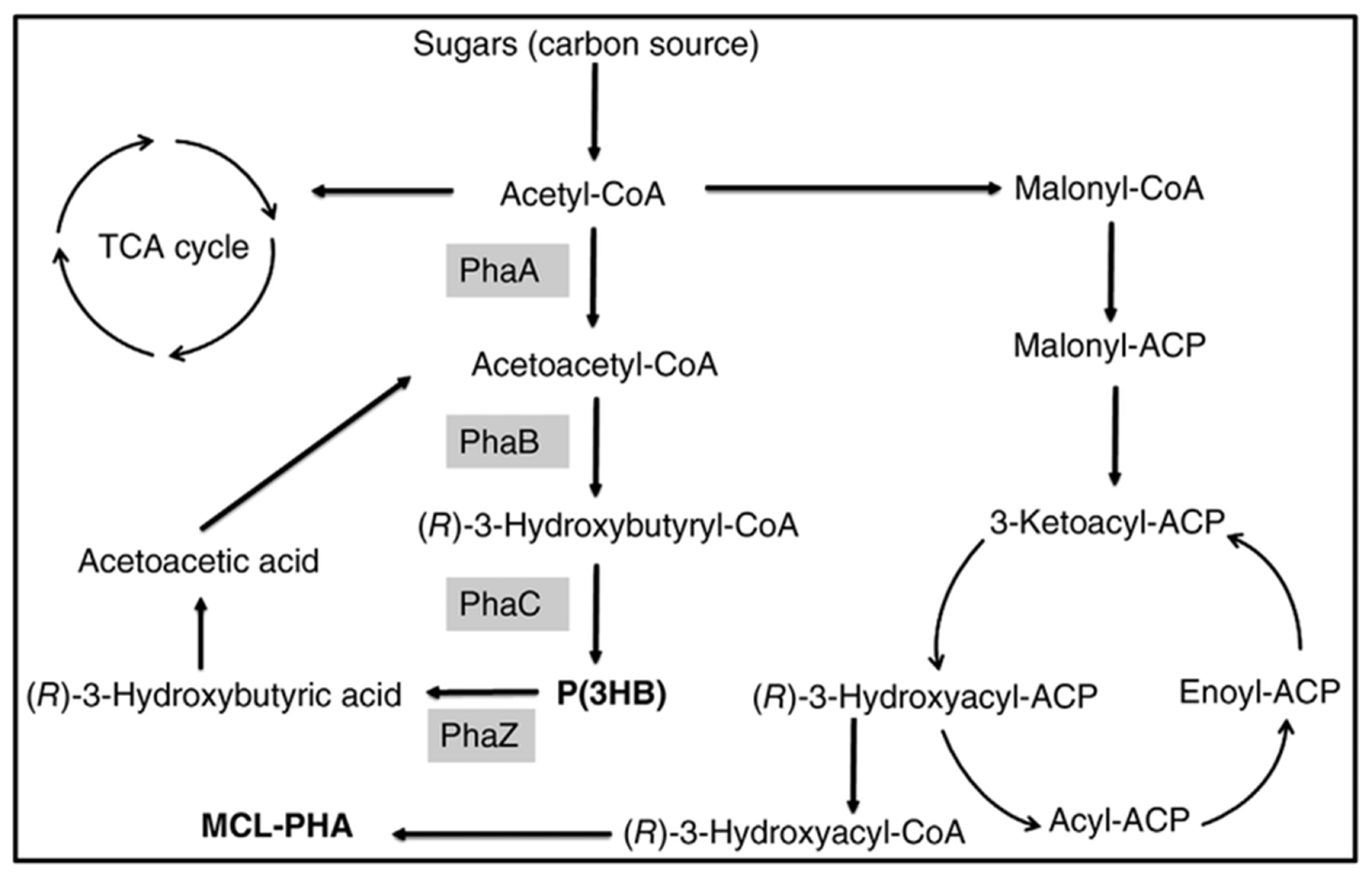
2.2.1. Fatty Acids
2.2.2. Carbohydrates
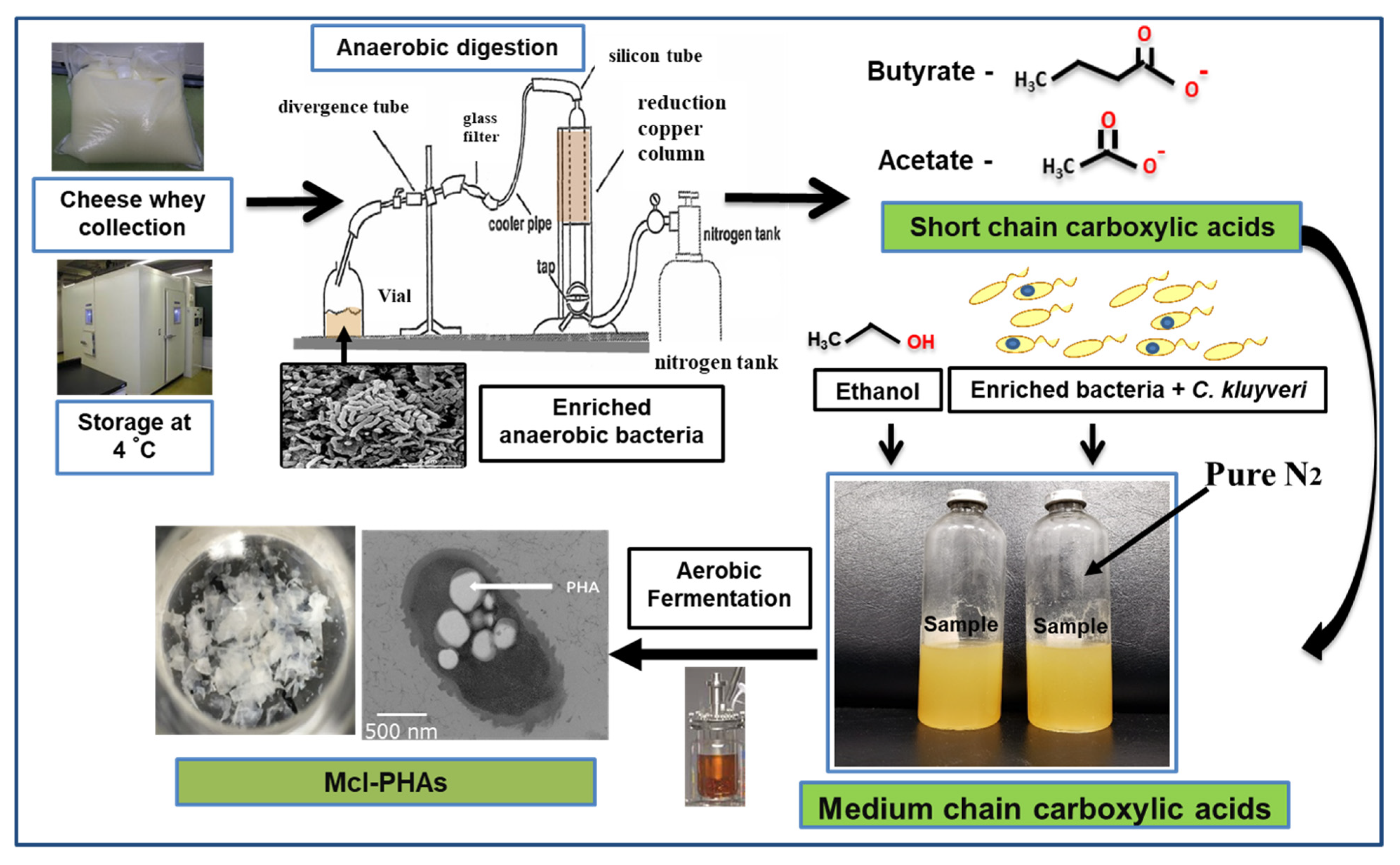
2.2.3. Organic Residues/Wastes and Others
2.2.4. Vegetable Oils
3. Mcl-PHA Production at Various Modes of Operations
3.1. Batch Mode
3.2. Continuous Mode
3.3. Fed-Batch Mode
4. Industries Producing PHAs
4.1. Europe
4.2. Asia
4.3. North and South America
5. News on PHAs
6. Extraction of PHAs
6.1. Solvent Extraction
6.2. Ultrasound-Assisted and Aqueous Two-Phase Extraction
6.3. Chemical and Enzymatic Digestion Method
7. Properties of PHAs
7.1. Melting Point, Crystallization, Gas Barrier, and Solubility
7.2. Mechanical Properties of PHAs
8. Applications of PHAs
9. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lemoigne, M. Dehydration and polymerization product of β-oxybutyric acid. Bull. Soc. Chim. Biol. 1926, 8, 770–782. [Google Scholar]
- Muhammadi, S.; Muhammad, A.; Shafqat, H. Bacterial polyhydroxyalkanoates-eco-friendly next generation plastic: Production, biocompatibility, biodegradation, physical properties and applications. Green Chem. Lett. Rev. 2015, 8, 56–77. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Chang, H.N. Production of poly (hydroxy alkanoic acid). Adv. Biochem. Eng. Biotechnol. 1995, 52, 27–58. [Google Scholar]
- Choonut, A.; Prasertsan, P.; Klomklao, S.; Sangkharak, K. A Novel Green Process for Synthesis of 3-Hydroxyalkanoate Methyl Ester Using Lipase and Novel mcl-co-lcl PHA as Catalyst and Substrate. J. Polym. Environ. 2021, 30, 1423–1434. [Google Scholar] [CrossRef]
- Rakkan, T.; Chana, N.; Chirapongsatonkul, N.; Sangkharak, K. Screening and identification of newly isolated basic red 9-degrading bacteria from textile wastewater and their ability to produce medium-co-long-chain-length polyhydroxyalkanoates. J. Polym. Environ. 2022, 30, 415–423. [Google Scholar] [CrossRef]
- Muhr, A.; Rechberger, E.M.; Salerno, A.; Reiterer, A.; Schiller, M.; Kwicien, M.; Adamus, G.; Kowalczuk, M.; Strohmeier, K.; Schober, S.; et al. Biodegradable latexes from animal-derived waste: Biosynthesis and characterization of mcl-PHA accumulated by P. citronellolis. React. Funct. Polym. 2013, 73, 1391–1398. [Google Scholar] [CrossRef]
- Nitschke, M.; Costa, S.G.V.A.O.; Contiero, J. Rhamnolipids and PHAs: Recent reports on Pseudomonas-derived molecules of increasing industrial interest. Process Biochem. 2011, 46, 621–630. [Google Scholar] [CrossRef]
- Chen, G.-Q.; Jiang, X.-R. Engineering bacteria for enhanced polyhydroxyalkanoates (PHA) biosynthesis. Synth. Syst. Biotechnol. 2017, 2, 192–197. [Google Scholar] [CrossRef]
- Kourmentza, C.; Plácido, J.; Venetsaneas, N.; Burniol-Figols, A.; Varrone, C.; Gavala, H.N.; Reis, M.A.M. Recent advances and challenges towards sustainable polyhydroxyalkanoate (PHA) production. Bioengineering 2017, 4, 55. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Wong, H.H.; Choi, J.; Lee, S.H.; Lee, S.C.; Han, C.S. Production of Medium-Chain-Length Polyhydroxyalkanoates by High-Cell-Density Cultivation of Pseudomonas putida Under Phosphorus Limitation. Biotechnol. Bioeng. 2000, 68, 466–470. [Google Scholar] [CrossRef]
- Riesenberg, D.; Guthke, R. High-cell-density cultivation of microorganisms. Appl. Microbiol. Biotechnol. 1999, 51, 422–430. [Google Scholar] [CrossRef]
- Cavalheiro, J.M.B.T.; Raposo, S.R.; Almeida, M.C.M.D.; Sevrin, M.T.C.C.; Grandfils, C.; Fonseca, M.M.R. Effect of cultivation parameters on the production of poly (3-hydroxybutyrate-co-4-hydroxybutyrate) and poly (3-hydroxybutyrate-4-hydroxybutyrate-3-hydroxyvalerate) by Cupriavidus necator using waste glycerol. Bioresour. Technol. 2012, 111, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.; Duane, G.; Kenny, S.T.; Cerrone, F.; Guzik, M.W.; Babu, R.P.; Casey, E.; O’Connor, K.E. High cell density cultivation of Pseudomonas putida KT2440 using glucose without the need for oxygen enriched air supply. Biotechnol. Bioeng. 2015, 112, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Ryu, H.W.; Hahn, S.K.; Chang, Y.K.; Chang, H.N. Production of poly(3-hydroxybutyrate) by high cell density fed-batch culture of Alcaligenes eutrophus with phosphate limitation. Biotechnol. Bioeng. 1997, 55, 25–32. [Google Scholar] [CrossRef]
- Chen, G.-O.; Zhang, J. Microbial polyhydroxyalkanoates as medical implant biomaterials. Artif. Cells Nanomed. Biotechnol. 2018, 46, 1–18. [Google Scholar] [CrossRef]
- Williams, S.F.; Rizk, S.; Martin, D.P. Poly-4-hydroxybutyrate (P4HB): A new generation of resorbable medical devices for tissue repair and regeneration. Biomed. Eng. 2013, 58, 439–452. [Google Scholar] [CrossRef]
- Skibiński, S.; Cichoń, E.; Haraźna, K.; Marcello, E.; Roy, I.; Witko, M.; Ślósarczyk, A.; Czechowska, J.; Guzik, M.; Zima, A. Functionalized tricalcium phosphate and poly (3-hydroxyoctanoate) derived composite scaffolds as platforms for the controlled release of diclofenac. Ceram. Int. 2021, 47, 3876–3883. [Google Scholar] [CrossRef]
- Cichoń, E.; Haraźna, K.; Skibiński, S.; Witko, T.; Zima, A.; Ślósarczyk, A.; Zimowska, M.; Witko, M.; Leszczyński, B.; Wróbel, A.; et al. Novel bioresorbable tricalcium phosphate/polyhydroxyoctanoate (TCP/PHO) composites as scaffolds for bone tissue engineering applications. J. Mech. Behav. Biomed. Mater. 2019, 98, 235–245. [Google Scholar] [CrossRef]
- Zhang, J.; Cao, Q.; Li, S.; Lu, X.; Zhao, Y.; Guan, J.-S.; Chen, J.-C.; Wu, Q.; Chen, G.-Q. 3-Hydroxybutyrate methyl ester as a potential drug against Alzheimer’s disease via mitochondria protection mechanism. Biomaterials 2013, 34, 7552–7562. [Google Scholar] [CrossRef]
- Xiao, X.-Q.; Zhao, Y.; Chen, G.-Q. The effect of 3-hydroxybutyrate and its derivatives on the growth of glial cells. Biomaterials 2007, 28, 3608–3616. [Google Scholar] [CrossRef]
- Heinrich, D.; Raberg, M.; Fricke, P.; Kenny, S.T.; Gamez, L.M.; Babu, R.P.; O’Connor, K.; Steinbüchel, A. Synthesis gas (syngas)-derived medium-chain-length polyhydroxyalkanoate synthesis in engineered Rhodospirillum Rubrum. Appl. Environ. Microbiol. 2016, 82, 6132–6140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaeger, K.E.; Steinbüchel, A.; Jendrossek, D. Substrate specificities of bacterial polyhydroxyalkanoate depolymerases and lipases: Bacterial lipases hydrolyze poly(ω-hydroxyalkanoates). Appl. Environ. Microbiol. 1995, 61, 3113–3118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volova, T.G.; Kiselev, E.G.; Shishatskaya, E.I.; Zhila, N.O.; Boyandin, A.N.; Syrvacheva, D.A.; Vinogradova, O.N.; Kalacheva, G.S.; Vasiliev, A.D.; Peterson, I.V. Cell growth and accumulation of polyhydroxyalkanoates from CO2 and H2 of a hydrogen-oxidizing bacterium, Cupriavidus eutrophus B-10646. Bioresour. Technol. 2013, 146, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Volova, T.G.; Shishatskaya, E.I.; Zhila, N.O.; Shishatskii, O.N.; Kiselev, Y.G.; Mironov, P.V.; Vasiliev, A.D.; Peterson, I.V.; Sinskey, A.J. Fundamental basis of production and application of biodegradable polyhydroxyalkanoates. J. Sib. Fed. Univ. Biol. 2012, 3, 280–299. [Google Scholar]
- Tanaka, K.; Yoshida, K.; Orita, I.; Fukui, T. Biosynthesis of Poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) from CO2 by a recombinant Cupriavidusnecator. Bioengineering 2021, 8, 179. [Google Scholar] [CrossRef]
- Löwe, H.; Hobmeier, K.; Moos, M.; Kremling, A.; Pflüger-Grau, K. Photoautotrophic production of polyhydroxyalkanoates in a synthetic mixed culture of Synechococcus elongatus cscB and Pseudomonas putida cscAB. Biotechnol. Biofuels 2017, 10, 1–14. [Google Scholar] [CrossRef]
- Riedel, S.L.; Bader, J.; Brigham, C.J.; Budde, C.F.; Yusof, Z.A.; Rha, C.; Sinskey, A.J. Production of poly (3-hydroxybutyrate- co-3-hydroxyhexanoate) by Ralstonia eutropha in high cell density palm oil fermentations. Biotechnol. Bioeng. 2012, 109, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, C.; Kenny, S.T.; Babu, P.R.; Walsh, M.; Narancic, T.; O’Connor, K.E. High cell density conversion of hydrolyzed waste cooking oil fatty acids into medium chain length polyhydroxyalkanoate using Pseudomonas putida KT2440. Catalysts 2019, 9, 468. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.J.; Sun, Z.; Ramsay, J.A.; Ramsay, B.A. Fed-batch production of MCL-PHA with elevated 3-hydroxynonanoate content. AMB Express 2013, 50, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Blunt, W.; Dartiailh, C.; Sparling, R.; Gapes, D.J.; Levin, D.B.; Cicek, N. Development of high cell density cultivation strategies for improved medium chain length polyhydroxyalkanoate productivity using Pseudomonas putida LS46. Bioengineering 2019, 6, 89. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Ramsay, J.A.; Guay, M.; Ramsay, B.A. Enhanced yield of medium-chain-length polyhydroxyalkanoates from nonanoic acid by co-feeding glucose in carbon-limited, fed-batch culture. J. Biotechnol. 2009, 143, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Diniz, S.C.; Taciro, M.K.; Gomez, J.G.C.; Pradella, J.G. High-cell-density cultivation of Pseudomonas putida IPT 046 and medium-chain-length polyhydroxyalkanoate production from sugarcane carbohydrates. Appl. Biochem. Biotech. 2004, 119, 1–69. [Google Scholar] [CrossRef]
- Kellerhals, M.B.; Hazenberg, W.; Witholt, B. High cell density fermentations of Pseudomonas oleovorans for the production of mcl-PHAs in two-liquid phase media. Enz. Microbiol. Technol. 1999, 24, 111–116. [Google Scholar] [CrossRef]
- Kellerhals, M.B.; Kessler, B.; Witholt, B.; Tchouboukov, A.; Brand, H. Renewable long-chain fatty acids for production of biodegradable medium-chain-length polyhydroxyalkanoates (mcl-PHAs) at laboratory and pilot plant scales. Macromolecules 2000, 33, 4690–4698. [Google Scholar] [CrossRef]
- Preusting, H.; Hazenberg, W.; Witholt, B. Continuous production of poly(3-hydroxyalkanoates) by Pseudomonas oleovorans in a high-ceil-density, two-liquid-phase chemostat. Enzym. Microb. Technol. 1993, 15, 311–316. [Google Scholar] [CrossRef]
- Preusting, H.; Houten, R.; Hoefs, A.; Langenberghe, E.K.; Favre-Bulle, O.; Witholt, B. High cell density cultivation of Pseudomonas oleovorans: Growth and production of poly (3-hydroxyalkanoates) in two-liquid phase batch and fed-batch systems. Biotechnol. Bioeng. 1993, 41, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Riedel, S.L.; Jahns, S.; Koenig, S.; Bock, M.C.; Brigham, C.J.; Bader, J.; Stahl, U. Polyhydroxyalkanoates production with Ralstonia eutropha from low quality waste animal fats. J. Biotechnol. 2015, 214, 119–127. [Google Scholar] [CrossRef]
- Sato, S.; Maruyama, H.; Fujiki, T.; Matsumoto, K. Regulation of 3-hydroxyhexanoate composition in PHBH synthesized by recombinant Cupriavidus necator H16 from plant oil by using butyrate as a co-substrate. J. Biosci. Bioeng. 2015, 120, 246–251. [Google Scholar] [CrossRef]
- Cai, L.; Yuan, M.Q.; Liu, F.; Jian, J.; Chen, G.Q. Enhanced production of medium-chain-length polyhydroxyalkanoates (PHA) by PHA depolymerase knockout mutant of Pseudomonas putida KT2442. Bioresour. Technol. 2009, 100, 2265–2270. [Google Scholar] [CrossRef]
- LeMeur, S.; Zinn, M.; Egli, T.; Thöny-Meyer, L.; Ren, Q. Production of medium-chain-length polyhydroxyalkanoates by sequential feeding of xylose and octanoic acid in engineered Pseudomonas putida KT2440. BMC Biotechnol. 2012, 12, 53. [Google Scholar]
- Kahar, P.; Tsuge, T.; Taguchi, K.; Doi, Y. High yield production of polyhydroxyalkanoates from soybean oil by Ralstonia eutropha and its recombinant strain. Polym. Degrad. Stab. 2004, 83, 79–86. [Google Scholar] [CrossRef]
- Jing, H.; Yuan-Zheng, Q.; Dai-Cheng, L.; Guo-Qiang, C. Engineered Aeromonas hydrophila for enhanced production of poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) with alterable monomers composition. FEMS Microbiol. Lett. 2004, 239, 195–201. [Google Scholar]
- Ouyang, S.; Han, J.; Qiu, Y.; Qin, L.; Chen, S.; Wu, Q.; Leski, M.L.; Chen, G. Poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) production in recombinant Aeromonas hydrophila 4ak4 harboring phba, phbb and vgb genes. Macromol. Symp. 2005, 224, 21–34. [Google Scholar] [CrossRef]
- Budde, C.F.; Riedel, S.L.; Willis, L.B.; Rha, C.; Sinskey, A.J. Production of poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) from plant oil by engineered Ralstonia eutropha strains. Appl. Environ. Microbiol. 2011, 77, 2847–2854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thinagaran, L.; Sudesh, K. Evaluation of sludge palm oil as feedstock and development to efficient method for its utilization to produce polyhydroxyalkanoate. Waste Biomass Valoriz. 2017, 10, 709–720. [Google Scholar] [CrossRef]
- Tufail, S.; Munir, S.; Jamil, N. Variation analysis of bacterial polyhydroxyalkanoates production using saturated and unsaturated hydrocarbons. Braz. J. Microbiol. 2017, 48, 629–636. [Google Scholar] [CrossRef]
- Qiu, Y.Z.; Han, J.; Guo, J.J.; Chen, G.Q. Production of poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) from gluconate and glucose by recombinant Aeromonas hydrophila and Pseudomonas putida. Biotechnol. Lett. 2005, 27, 1381–1386. [Google Scholar] [CrossRef]
- Poblete-Castro, I.; Rodriguez, A.L.; Lam, C.M.; Kessler, W. Improved production of medium-chain-length polyhydroxyalkanoates in glucose-based fed-batch cultivations of metabolically engineered Pseudomonas putida strains. J. Microbiol. Biotechnol. 2014, 24, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; He, F.; Liu, X.; Shi, J.; Liang, J.; Wang, S.; Yang, C.; Liu, R. Metabolic engineering of Pseudomonas mendocina NK-01 for enhanced production of medium-chain-length polyhydroxyalkanoates with enriched content of the dominant monomer. Int. J. Biol. Macromol. 2020, 154, 1596–1605. [Google Scholar] [CrossRef]
- Liu, C.; Qi, L.; Yang, S.; He, Y.; Jia, X. Increased sedimentation of a Pseudomonas–Saccharomyces microbial consortium producing medium chain length polyhydroxyalkanoates. Chin. J. Chem. Eng. 2019, 27, 1659–1665. [Google Scholar] [CrossRef]
- Oliveira, G.H.D.; Zaiat, M.; Rodrigues, J.A.D.; Ramsay, J.A.; Ramsay, B.A. Towards the production of mcl-pha with enriched dominant monomer content: Process development for the sugarcane biorefinery context. J. Polym. Environ. 2020, 28, 844–853. [Google Scholar] [CrossRef]
- Insomphun, C.; Mifune, J.; Orita, I.; Numata, K.; Nakamura, S.; Fukui, T. Modification of β-oxidation pathway in Ralstonia eutropha for production of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from soybean oil. J. Biosci. Bioeng. 2014, 117, 184–190. [Google Scholar] [CrossRef]
- Serafim, L.S.; Lemos, P.C.; Albuquerque, M.G.; Reis, M.A. Strategies for PHA production by mixed cultures and renewable waste materials. Appl. Microbiol. Biotechnol. 2008, 81, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.; Kataria, R.; Cerrone, F.; Woods, T.; Kenny, S.; O’Donovan, A.; Guzik, M.; Shaikh, H.; Duane, G.; Gupta, V.K.; et al. Conversion of grass biomass into fermentable sugars and its utilization for medium chain length polyhydroxyalkanoate (mcl-PHA) production by Pseudomonas strains. Bioresour. Technol. 2013, 150, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, M.K.; Kumar, V.; Yadav, V.; Sarsaiya, S.; Awasthi, S.K.; Sindhu, R.; Zhang, Z. Current state of the art biotechnological strategies for conversion of watermelon wastes residues to biopolymers production: A review. Chemosphere 2022, 290, 133310. [Google Scholar] [CrossRef]
- Muhr, A.; Rechberger, E.M.; Salerno, A.; Reiterer, A.; Malli, K.; Strohmeier, K.; Koller, M. Novel description of mcl-PHA biosynthesis by Pseudomonas chlororaphis from animal-derived waste. J. Biotechnol. 2013, 165, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Andler, R.; Valdés, C.; Urtuvia, V.; Andreeßen, C.; Díaz-Barrera, A. Fruit residues as a sustainable feedstock for the production of bacterial polyhydroxyalkanoates. J. Clean. Prod. 2021, 307, 127236. [Google Scholar] [CrossRef]
- Pernicova, I.; Enev, V.; Marova, I.; Obruca, S. Interconnection of waste chicken feather biodegradation and keratinase and mcl-PHA production employing Pseudomonas putida KT2440. Appl. Food Biotechnol. 2019, 6, 83–90. [Google Scholar]
- Liu, H.; Kumar, V.; Jia, L.; Sarsaiya, S.; Kumar, D.; Juneja, A.; Awasthi, M.K. Biopolymer poly-hydroxyalkanoates (PHA) production from apple industrial waste residues: A review. Chemosphere 2021, 284, 131427. [Google Scholar] [CrossRef]
- Blanco, F.G.; Hernández, N.; Rivero-Buceta, V.; Maestro, B.; Sanz, J.M.; Mato, A.; Prieto, M.A. From residues to added-value bacterial biopolymers as nanomaterials for biomedical applications. Nanomaterials 2021, 11, 1492. [Google Scholar] [CrossRef]
- Yadav, B.; Pandey, A.; Kumar, L.R.; Tyagi, R.D. Bioconversion of waste (water)/residues to bioplastics-A circular bioeconomy approach. Bioresour. Technol. 2020, 298, 122584. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Venkateswar Reddy, M.; Imura, K.; Onodera, R.; Kamada, N.; Sano, Y. Two-Stage polyhydroxyalkanoates (PHA) production from cheese whey using Acetobacter pasteurianus C1 and Bacillus sp. CYR1. Bioengineering 2021, 8, 157. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Venkateswar Reddy, M.; Choi, D.B. Cometabolic degradation of toxic trichloroethene or cis-1,2-dichloroethene with phenol and production of poly-β-hydroxybutyrate (PHB). Green Chem. 2021, 23, 2729–2737. [Google Scholar] [CrossRef]
- Reddy, M.V.; Watanabe, A.; Onodera, R.; Mawatari, Y.; Tsukiori, Y.; Watanabe, A.; Kudou, M.; Chang, Y.C. Polyhydroxyalkanoates (PHA) production using single or mixture of fatty acids with Bacillus sp. CYR1: Identification of PHA synthesis genes. Bioresour. Technol. Rep. 2020, 11, 100483–100491. [Google Scholar] [CrossRef]
- Reddy, M.V.; Mawatari, S.; Onodera, R.; Nakamura, Y.; Yajima, Y.; Chang, Y.C. Bacterial conversion of waste into polyhydroxybutyrate (PHB): A new approach of bio-circular economy for treating waste and energy generation. Bioresour. Technol. Rep. 2019, 7, 100246–100254. [Google Scholar] [CrossRef]
- Reddy, M.V.; Mawatari, Y.; Onodera, R.; Nakamura, Y.; Yajima, Y.; Chang, Y.C. Polyhydroxyalkanoates (PHA) production from synthetic waste using Pseudomonas pseudoflava: PHA synthase enzyme activity analysis from P. pseudoflava and P. palleronii. Bioresour. Technol. 2017, 234, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Reddy, M.V.; Mawatari, Y.; Yajima, Y.; Satoh, K.; Venkata Mohan, S.; Chang, Y.C. Production of poly-3-hydroxybutyrate (P3HB) and poly-3-(hydroxybutyrate-co-hydroxyvalerate) P(3HB-co-3HV) from synthetic wastewater using Hydrogenophaga palleronii. Bioresour. Technol. 2016, 215, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Amulya, K.; Venkateswar Reddy, M.; Rohit, M.V.; Mohan, S.V. Wastewater as renewable feedstock for polyhydroxyalkanoates (PHA) production: Understanding the role of reactor microenvironment and system pH. J. Clean. Prod. 2016, 112, 4618–4627. [Google Scholar] [CrossRef]
- Amulya, K.; Venkateswar Reddy, M.; Mohan, S.V. Acidogenic spent wash valorization through polyhydroxyalkanoate (PHA) synthesis coupled with fermentative biohydrogen production. Bioresour. Technol. 2014, 158, 336–342. [Google Scholar] [CrossRef]
- Reddy, M.V.; Mohan, S.V. Influence of aerobic and anoxic microenvironments on polyhydroxyalkanoates (PHA) production from food waste and acidogenic effluents using aerobic consortia. Bioresour. Technol. 2012, 103, 313–321. [Google Scholar] [CrossRef]
- Srikanth, S.; Venkateswar Reddy, M.; Mohan, S.V. Microaerophilic microenvironment at biocathode enhances electrogenesis with simultaneous synthesis of polyhydroxyalkanoates (PHA) in bioelectrochemical system (BES). Bioresour. Technol. 2012, 125, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.V.; Venkateswar Reddy, M.; Subhash, G.V.; Sarma, P.N. Fermentative effluents from hydrogen producing bioreactor as substrate for poly (β-OH) butyrate production with simultaneous treatment: An integrated approach. Bioresour. Technol. 2010, 101, 9382–9386. [Google Scholar] [CrossRef] [PubMed]
- Appaiah, P.; Sunil, L.; Prasanth Kumar, P.K.; Gopala Krishna, A.G. Composition of Coconut Testa, Coconut Kernel and its Oil. J. Am. Oil Chem. Soc. 2014, 91, 917–924. [Google Scholar] [CrossRef]
- Basnett, P.; Marcello, E.; Lukasiewicz, B.; Panchal, B.; Nigmatullin, B.; Knowles, J.C.; Roy, I. Biosynthesis and characterization of a novel, biocompatible medium chain length polyhydroxyalkanoate by Pseudomonas mendocina CH50 using coconut oil as the carbon source. J. Mater. Sci. Mater. Med. 2018, 29, 1–11. [Google Scholar] [CrossRef]
- Valappil, S.P.; Misra, S.K.; Boccaccini, A.R.; Roy, I. Biomedical applications of polyhydroxyalkanoates (PHAs), an overview of animal testing and in vivo responses. Expert Rev. Med. Devices 2006, 3, 853–868. [Google Scholar] [CrossRef]
- Możejko, J.; Ciesielski, S. Saponified waste palm oil as an attractive renewable resource for mcl-polyhydroxyalkanoate synthesis. J. Biosci. Bioeng. 2013, 116, 485–492. [Google Scholar] [CrossRef]
- Mozejko, J.; Wilke, A.; Przybylek, G.; Ciesielski, S. Mcl-PHAs produced by Pseudomonas sp. Gl01 using fed-batch cultivation with waste rapeseed oil as carbon source. J. Microbiol. Biotechnol. 2012, 22, 371–377. [Google Scholar] [CrossRef] [Green Version]
- Song, J.H.; Jeon, C.O.; Choi, M.H.; Yoon, S.C.; Park, W.J. Polyhydroxyalkanoate (PHA) production using waste vegetable oil by Pseudomonas sp. strain DR2. J. Microbiol. Biotechnol. 2008, 18, 1408–1415. [Google Scholar]
- Reddy, M.V.; Yajima, Y.; Mawatari, Y.; Hoshino, T.; Chang, Y.C. Degradation and conversion of toxic compounds into useful bioplastics by Cupriavidus sp. CY-1: Relative expression of PhaC gene under phenol and nitrogen stress. Green Chem. 2015, 17, 4560–4569. [Google Scholar] [CrossRef] [Green Version]
- Mohan, S.V.; Venkateswar Reddy, M. Optimization of critical factors to enhance polyhydroxyalkanoates (PHA) synthesis by mixed culture using Taguchi design of experimental methodology. Bioresour. Technol. 2013, 128, 409–416. [Google Scholar] [CrossRef]
- Reddy, M.V.; Mohan, S.V. Effect of substrate load and nutrients concentration on the Polyhydroxyalkanoates (PHA) production using mixed consortia through wastewater treatment. Bioresour. Technol. 2012, 114, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Rai, R.; Keshavarz, T.; Roether, J.A.; Boccaccini, A.R.; Roy, L. Medium chain length polyhydroxyalkanoates, promising new biomedical materials for the future. Mater. Sci. Eng. R Rep. 2011, 72, 29–47. [Google Scholar] [CrossRef]
- Jung, K.; Hazenberg, W.; Prieto, M.; Witholt, B. Two-stage continuous process development for the production of medium-chain-length poly(3-hydroxyalkanoates). Biotech. Bioeng. 2001, 71, 19–24. [Google Scholar] [CrossRef]
- Lee, M.Y.; Park, W.H.; Lenz, R.W. Hydrophilic bacterial polyesters modified with pendant hydroxyl groups. Polymer 2000, 41, 1703–1709. [Google Scholar] [CrossRef]
- Gao, J.; Ramsay, J.A.; Ramsay, B.A. Fed-batch production of poly-3-hydroxydecanoate from decanoic acid. J. Biotechnol. 2016, 218, 102–107. [Google Scholar] [CrossRef]
- Cerrone, F.; Duane, G.; Casey, E.; Davis, R.; Belton, I.; Kenny, S.T.; Guzik, M.W.; Woods, T.; Babu, R.P.; O’Connor, K. Fed-batch strategies using butyrate for high cell density cultivation of Pseudomonas putida and its use as a biocatalyst. Appl. Microbiol. Biotechnol. 2014, 98, 9217–9228. [Google Scholar] [CrossRef]
- Sun, Z.; Ramsay, J.A.; Guay, M.; Ramsay, B.A. Carbon-limited fed-batch production of medium-chain-length polyhydroxyalkanoates from nonanoic acid by Pseudomonas putida KT2440. Appl. Microbiol. Biotechnol. 2007, 74, 69–77. [Google Scholar] [CrossRef]
- Kim, B.S. Production of medium chain length polyhydroxyalkanoates by fed-batch culture of Pseudomonas oleovorans. Biotechnol. Lett. 2002, 24, 125–130. [Google Scholar] [CrossRef]
- Kim, G.J.; Lee, I.Y.; Yoon, S.C.; Shin, Y.C.; Park, Y.H. Enhanced yield and a high production of medium-chain-length poly(3-hydroxyalkanoates) in a two-step fed-batch cultivation of Pseudomonas putida by combined use of glucose and octanoate. Enzym. Microb. Technol. 1997, 20, 500–505. [Google Scholar] [CrossRef]
- Kaur, G.; Roy, I. Strategies for large-scale production of polyhydroxyalkanoates. Chem. Biochem. Eng. Q. 2015, 29, 157–172. [Google Scholar] [CrossRef]
- Zinn, M.; Witholt, B.; Egli, T. Occurrence, synthesis and medical application of bacterial polyhydroxyalkanoate. Adv. Drug. Del. Rev. 2001, 53, 5–21. [Google Scholar] [CrossRef]
- Egli, T. On multiple-nutrient-limited growth of microorganisms, with special reference to dual limitation by carbon and nitrogen substrates. Antonie Van Leeuwenhoek 1991, 60, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Huijberts, G.N.M.; Eggink, G. Production of poly(3-hydroxyalkanoates) by Pseudomonas putida KT2442 in continuous cultures. Appl. Microbiol. Biotechnol. 1996, 46, 233–239. [Google Scholar] [CrossRef]
- McNeil, B.; Harvey, L.M. Fermentation, a Practical Approach; IRL: Tokyo, Japan, 1990. [Google Scholar]
- Andin, N.; Longieras, A.; Veronese, T.; Marcato, F.; Molina-Jouve, C.; Uribelarrea, J.-L. Improving carbon and energy distribution by coupling growth and medium chain length polyhydroxyalkanoate production from fatty acids by Pseudomonas putida KT2440. Biotechnol. Bioprocess Eng. 2017, 22, 308–318. [Google Scholar] [CrossRef]
- Bio-On Declares Bankruptcy. Available online: http://www.plasticsnews.com/news/bio-declares-bankruptcy (accessed on 3 January 2020).
- A Bio-on e a Hera Criam a Lux-on, o Novo Desafio Para Produzir Bioplástico a Partir de CO2. Available online: http://www.bio-on.it/project.php?lin=portoghese (accessed on 10 December 2018).
- Upcycling Waste to Natural Biopolymers. Available online: http://www.paquesbiomaterials.nl (accessed on 10 December 2018).
- Bioextrax develops world-leading bio-based technologies – accelerating the transition to a sustainable global economy. Available online: http://bioextrax.com (accessed on 25 March 2022).
- Guzik, M.W. Polyhydroxyalkanoates, bacterially synthesized polymers, as a source of chemical compounds for the synthesis of advanced materials and bioactive molecules. Appl. Microbiol. Biotechnol. 2021, 105, 7555–7566. [Google Scholar] [CrossRef]
- Titan An Biopolymers/Enmat. Available online: http://www.tianan-enmat.com/ (accessed on 25 March 2022).
- Mitsubishi Gas Chemical (MGC). Available online: https://www.mgc.co.jp/ (accessed on 25 March 2022).
- Completion of Kaneka Biodegradable Polymer PHBH™ Plant with Annual Production of 5,000 Tons. Available online: http://www.kaneka.co.jp/en/service/news/nr20191219/ (accessed on 19 December 2019).
- CJ Acquires Intellectual Property and Lab Equipment from US Venture Firm. Available online: http://www.ajudaily.com/view/20160823101739592 (accessed on 23 August 2016).
- Polymeron. Available online: http://www.polymerofcanada.com/versamerphas.html (accessed on 25 March 2022).
- Noda, I.; Lindsey, S.B.; Caraway, D. NodaxTM class PHA copolymers: Their properties and applications. In Plastics from Bactria: Natural Functions and Applications; Chen, G.-Q., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 237–255. [Google Scholar]
- Biocycle. Available online: http://ww.biocycle.com.br (accessed on 25 March 2022).
- Yield10 BIOSCIENCE. Available online: http://www.yield10bio.com (accessed on 25 March 2022).
- Tepha Medical Devices. Tepha Medical Devices. Available online: http://www.tepha.com/technology/polymer-processing-material-attributes (accessed on 25 March 2022).
- Jennifer, B.; Emily, G. Plastic from thin air. Popul.Sci. 2014, 285, 24. [Google Scholar]
- Can Plastic Be Made Environmentally Friendly? Available online: http://www.scientificamerican.com/article/can-plastic-be-made-environmentally-friendly (accessed on 29 January 2014).
- Polyhydroxyalkanoate (PHA) Market by Type (Short Chain Length, Medium Chain Length), Production Method (Sugar Fermentation, Vegetable Oil Fermentation, Methane Fermentation), Application, and Region - Global Forecast to 2025. Available online: http://www.marketsandmarkets.com/Market-Reports/pha-market-395.html (accessed on 1 February 2021).
- MARS. Available online: http://www.mars.com/sustainability-plan/healthy-planet/sustainable-packaging (accessed on 25 March 2022).
- After 149 Years, Colgate’s Toothpaste Tubes are Finally Recyclable. Available online: http://www.fastcompany.com/90713412/after-149-years-colgates-toothpaste-tubes-are-finally-recyclable (accessed on 19 January 2022).
- Kosseva, M.R.; Rusbandi, E. Trends in the biomanufacturer of polyhydroxyalkanoates with focus on downstream processing. Int. J. Biol. Macromol. 2018, 107, 762–778. [Google Scholar] [CrossRef]
- Bresan, S.; Sznajder, A.; Hauf, W.; Forchhammer, K.; Pfeiffer, D.; Jendrossek, D. Polyhydroxyalkanoate (PHA) granules have no phospholipids. Sci. Rep. 2016, 6, 26612. [Google Scholar] [CrossRef] [Green Version]
- Jacquel, N.; Lo, C.W.; Wei, Y.H.; Wu, H.S.; Wang, S.S. Isolation and purification of bacterial poly (3-hydroxyalkanoates). Biochem. Eng. J. 2008, 39, 15–27. [Google Scholar] [CrossRef]
- Koller, M.; Niebelschütz, H.; Braunegg, G. Strategies for recovery and purification of poly[(R)-3-hydroxyalkanoates] (PHA) bio polyesters from surrounding biomass. Eng. Life Sci. 2013, 13, 549–562. [Google Scholar] [CrossRef]
- Furrer, P.; Panke, S.; Zinn, M. Efficient recovery of low endotoxin medium-chain length poly([R]-3-hydroxyalkanoate) from bacterial biomass. J. Microbiol. Methods 2007, 69, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Chemat, F.; Vian, M.A.; Cravotto, G. Green extraction of natural products: Concept and principles. Int. J. Mol. Sci. 2012, 13, 8615–8627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yabueng, N.; Napathorn, S.C. Toward non-toxic and simple recovery process of poly (3-hydroxybutyrate) using the green solvent 1,3-dioxolane. Process Biochem. 2018, 69, 197–207. [Google Scholar] [CrossRef]
- Ishak, K.A.; Annuar, M.S.M.; Heidelberg, T.; Gumel, A.M. Ultrasound-assisted rapid extraction of bacterial intra cellular medium- chain-length poly(3-hydroxyalkanoates) (mcl-phas) in medium mixture of solvent/marginal non-solvent. Arab. J. Sci. Eng. 2016, 41, 33–44. [Google Scholar] [CrossRef]
- Noda, I. Process for Recovering Polyhydroxyalkanotes Using Air Classification. U.S. Patent US5849854A, 15 December 1998. [Google Scholar]
- Vilkhu, K.; Mawson, R.; Simons, L.; Bates, D. Applications and opportunities for ultrasound assisted extraction in the food industry-A review. Innov. Food Sci. Emerg. Technol. 2008, 9, 161–169. [Google Scholar] [CrossRef]
- Leong, Y.K.; Koroh, F.E.; Show, P.L.; Lan, J.C.W.; Loh, H.S. Optimization of extractive bioconversion for green polymer via aqueous two-phase system. Chem. Eng. Trans. 2015, 45, 1495–1500. [Google Scholar]
- Leong, Y.K.; Lan, J.C.W.; Loh, H.S.; Ling, T.C.; Ooi, C.W.; Show, P.L. Cloud-point extraction of green-polymers from Cupriavidus necator lysate using thermo separating-based aqueous two-phase extraction. J. Biosci. Bioeng. 2016, 123, 3270–3375. [Google Scholar]
- Leong, Y.K.; Show, P.L.; Lan, J.C.-W.; Loh, H.-S.; Yap, Y.-J.; Ling, T.C. Extraction and purification of polyhydroxyalkanoates (PHAs): Application of thermo separating aqueous two-phase extraction. J. Polym. Res. 2017, 24, 158. [Google Scholar] [CrossRef]
- Leong, K.; Show, P.L.; Lan, J.C.W.; Krishna Moorthy, R.; Chu, D.T.; Nagarajan, D.; Yen, H.W.; Chang, J.S. Application of thermo-separating aqueous two-phase system in extractive bioconversion of polyhydroxyalkanoates by Cupriavidus necator H16. Bioresour. Technol. 2019, 287, 121474. [Google Scholar] [CrossRef]
- López-Abelairas, M.; García-Torreiro, M.; Lú-Chau, T.; Lema, J.M.; Steinbüchel, A. Comparison of several methods for the separation of poly(3-hydroxybutyrate) from Cupriavidus necator H16 cultures. Biochem. Eng. J. 2015, 93, 250–259. [Google Scholar] [CrossRef]
- Gumel, A.M.; Annuar, M.S.M.; Chisti, Y. Recent advances in the production, recovery, and applications of polyhydroxyalkanoates. J. Polym. Environ. 2013, 21, 580–605. [Google Scholar] [CrossRef]
- Dong, Z.; Sun, X.; Zhaolin, D.; Xuenan, S.U.N. A new method of recovering polyhydroxyalkanoate from Azotobacter chroococcum. Chin. Sci. Bull. 2000, 45, 252–256. [Google Scholar] [CrossRef]
- Yasotha, K.; Aroua, M.K.; Ramachandran, K.B.; Tan, I.K.P. Recovery of medium-chain-length polyhydroxyalkanoates (PHAs) through enzymatic digestion treatments and ultrafiltration. Biochem. Eng. J. 2006, 30, 260–268. [Google Scholar] [CrossRef]
- Kachrimanidou, V.; Kopsahelis, N.; Vlysidis, A.; Papanikolaou, S.; Kookos, I.K.; Martínez, B.M.; Rondan, M.C.E.; Kautinas, A.A. Downstream separation of poly(hydroxyalkanoates) using crude enzyme consortia produced via solid state fermentation integrated in a biorefinery concept. Food Bioprod. Process 2016, 100, 323–334. [Google Scholar] [CrossRef]
- Israni, N.; Thapa, S.; Shivakumar, S. Biolytic extraction of poly (3-hydroxybutyrate) from Bacillus megaterium Ti3 using the lytic enzyme of Streptomyces albus Tia1. J. Genet. Eng. Biotechnol. 2018, 16, 265–271. [Google Scholar] [CrossRef]
- Panaitescu, D.M.; Lupescu, I.; Frone, A.N.; Chiulan, I.; Nicolae, C.A.; Tofan, V.; Stefaniu, A.; Somoghi, R.; Trusca, R. Medium chain-length polyhydroxyalkanoate copolymer modified by bacterial cellulose for medical devices. Biomacromolecules 2017, 18, 3222–3232. [Google Scholar] [CrossRef]
- Abe, H.; Ishii, N.; Sato, S.; Tsuge, T. Thermal properties and crystallization behaviors of medium-chain-length poly (3-hydroxyalkanoate) s. Polymer 2012, 53, 3026–3034. [Google Scholar] [CrossRef]
- Chen, G.Q. A microbial polyhydroxyalkanoates (PHA) based bio and material industry. Chem. Soc. Rev. 2009, 38, 2434–2446. [Google Scholar] [CrossRef]
- Gopi, S.; Kontopoulou, M.; Ramsay, B.A.; Ramsay, J.A. Manipulating the structure of medium-chain-length polyhydroxyalkanoate (MCL-PHA) to enhance thermal properties and crystallization kinetics. Int. J. Biol. Macromol. 2018, 119, 1248–1255. [Google Scholar] [CrossRef]
- Nerkar, M.; Ramsay, J.A.; Ramsay, B.A.; Kontopoulou, M. Melt compounded blends of short and medium chain-length poly-3-hydroxyalkanoates. J. Polym. Environ. 2014, 22, 236–243. [Google Scholar] [CrossRef]
- Xiang, H.; Chen, W.; Chen, Z.; Sun, B.; Zhu, M. Significant accelerated crystallization of long chain branched poly (3-hydroxybutyrate-co-3-hydroxyvalerate) with high nucleation temperature under fast cooling rate. Compos. Sci. Technol. 2017, 142, 207–213. [Google Scholar] [CrossRef]
- Noda, I.; Schechtman, L.A. Solvent extraction of polyhydroxyalkanoates from biomass. U.S. Patent 5,942,597, 24 August 1999. [Google Scholar]
- Sobieski, B.J.; Gong, L.; Aubuchon, S.R.; Noda, I.; Chase, D.B.; Rabolt, J.F. Thermally reversible physical gels of poly[(R)-3-hydroxybutyrate-co-(R)-3-hydroxyhexanoate]: Part 1 gelation in dimethylformamide. J. Polym. 2017, 131, 217–223. [Google Scholar] [CrossRef]
- Noda, I.; Green, P.R.; Satkowski, M.M.; Schechtman, L.A. Preparation and properties of a novel class of polyhydroxyalkanoate copolymers. Biomacromolecules 2005, 6, 580–586. [Google Scholar] [CrossRef]
- Tanadchangsaeng, N.; Tsuge, T.; Abe, H. Comonomer compositional distribution, physical properties, and enzymatic degradability of bacterial poly (3-hydroxybutyrate-co-3-hydroxy-4-methylvalerate) polyesters. Biomacromolecules 2010, 11, 1615–1622. [Google Scholar] [CrossRef] [PubMed]
- Kolahchi, A.R.; Kontopoulou, M. Chain extended poly (3-hydroxybutyrate) with improved rheological properties and thermal stability, through reactive modification in the melt state. Polym. Degrad. Stab. 2015, 121, 222–229. [Google Scholar] [CrossRef] [Green Version]
- Muthuraj, R.; Misra, M.; Mohanty, A.K. Biodegradable compatibilized polymer blends for packaging applications: A literature review. J. Appl. Polym. Sci. 2018, 135, 45726. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Misra, M.; Mohanty, A.K. Challenges and new opportunities on barrier performance of biodegradable polymers for sustainable packaging. Prog. Polym. Sci. 2021, 117, 101395. [Google Scholar] [CrossRef]
- Xu, P.; Yang, W.; Niu, D.; Yu, M.; Du, M.; Dong, W.; Chen, M.; Lemstra, P.J.; Ma, P. Multifunctional and robust polyhydroxyalkanoate nanocomposites with superior gas barrier, heat resistant and inherent antibacterial performances. Chem. Eng. J. 2020, 382, 122864. [Google Scholar] [CrossRef]
- Choi, S.Y.; Cho, I.J.; Lee, Y.; Kim, Y.; Kim, K.; Lee, S.Y. Microbial polyhydroxyalkanoates and nonnatural polyesters. Adv. Mater. 2020, 32, 1907138. [Google Scholar] [CrossRef]
- Bugnicourt, E.; Cinelli, P.; Lazzeri, A.; Alvarez, V.A. Polyhydroxyalkanoate (PHA): Review of synthesis, characteristics, processing and potential applications in packaging. Express Polym. Lett. 2014, 8, 791–808. [Google Scholar] [CrossRef] [Green Version]
- Kusaka, S.; Abe, H.; Lee, S.; Doi, Y. Molecular mass of poly [(R)-3-hydroxybutyric acid] produced in a recombinant Escherichia coli. Appl. Microbiol. Biotechnol. 1997, 47, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Lee, S.Y. High level production of supra molecular weight poly (3-hydroxybutyrate) by metabolically engineered Escherichia coli. Biotechnol. Bioprocess Eng. 2004, 9, 196–200. [Google Scholar] [CrossRef]
- Doi, Y.; Kitamura, S.; Abe, H. Microbial synthesis and characterization of poly (3-hydroxybutyrate-co-3-hydroxyhexanoate). Macromolecules 1995, 28, 4822–4828. [Google Scholar] [CrossRef]
- Acuña, J.M.; Rohde, M.; Saldias, C.; Castro, I.P. Fed-Batch mcl-Polyhydroxyalkanoates production in Pseudomonas putida KT2440 and phaZ mutant on biodiesel-derived crude glycerol. Front. Bioeng. Biotechnol. 2021, 9, 1–10. [Google Scholar]
- Dartiailh, C.; Blunt, W.; Sharma, P.K.; Liu, S.; Cicek, N.; Levin, D.B. The thermal and mechanical properties of medium chain-length polyhydroxyalkanoates produced by Pseudomonas putida LS46 on various substrates. Front. Bioeng. Biotechnol. 2021, 8, 1–9. [Google Scholar] [CrossRef]
- Liu, W.; Chen, G.Q. Production and characterization of mediumchain-length polyhydroxyalkanoate with high 3-hydroxytetradecanoate monomer content by fadB and fadA knockout mutant of Pseudomonas putida KT2442. Appl. Microbiol. Biotechnol. 2007, 76, 1153–1159. [Google Scholar] [CrossRef]
- Solaiman, D.K.Y.; Ashby, R.D.; Foglia, T.A. Physiological characterization and genetic engineering of Pseudomonas corrugata for medium-chain-length polyhydroxyalkanoates synthesis from triacylglycerols. Curr. Microbiol. 2002, 44, 189–195. [Google Scholar] [CrossRef]
- Ashby, R.D.; Foglia, T.A. Poly(hydroxyalkanoate) biosynthesis from triglyceride substrates. Appl. Microbiol. Biotechnol. 1998, 49, 431–437. [Google Scholar] [CrossRef]
- Hazer, D.B.; Hazer, B.; Kaymaz, F. Synthesis of microbial elastomers based on soybean oily acids. Biocompatibility studies. Biomed. Mater. 2009, 4, 1–9. [Google Scholar] [CrossRef]
- Kalia, V.C.; Patel, S.K.S.; Shanmugam, R.; Lee, J.K. Polyhydroxyalkanoates: Trends and advances toward biotechnological applications. Bioresour. Technol. 2021, 326, 124737. [Google Scholar] [CrossRef]
- Dill, S.; Demicheva, M.; Fleschutz, B.; Weinlein, R.; Demicheva, M.; Fleschutz, B.; Weinlein, R. Influence of polyhydroxybutyrate content on the crystallization behavior of polyamide 6-polyhydroxy-butyrate blends. Macromol. Symp. 2019, 384, 1800170. [Google Scholar] [CrossRef]
- Wang, S.; Chen, W.; Xiang, H.; Yang, J.; Zhou, Z.; Zhu, M. Modification and potential application of short-chain-length polyhydroxyalkanoate (SCL-PHA). Polymers 2016, 8, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastogi, V.; Samyn, P. Bio-based coatings for paper applications. Coatings 2015, 5, 887–930. [Google Scholar] [CrossRef] [Green Version]
- Xiang, H.; Chen, Z.; Zheng, N.; Zhang, X.; Zhu, L.; Zhou, Z.; Zhu, M. Melt-spun microbial poly (3-hydroxybutyrate-co-3-hydroxyvalerate) fibers with enhanced toughness: Synergistic effect of heterogeneous nucleation, long-chain branching and drawing process. Int. J. Biol. Macromol. 2019, 122, 1136–1143. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.; Poursorkhabi, V.; Mohanty, A.K.; Misra, M. Analysis of porous electrospun fibers from poly (l-lactic acid)/Poly (3-hydroxybutyrate-co-3-hydroxyvalerate) blends. ACS Sustain. Chem. Eng. 2014, 2, 1976–1982. [Google Scholar] [CrossRef]
- Ivanov, V.; Stabnikov, V.; Ahmed, Z.; Dobrenko, S.; Saliuk, A. Production, and applications of crude polyhydroxyalkanoate-containing bioplastic from the organic fraction of municipal solid waste. Int. J. Environ. Sci. Technol. 2015, 12, 725–738. [Google Scholar] [CrossRef] [Green Version]
- Ang, S.L.; Sivashankari, R.; Shaharuddin, B.; Chuah, J.-A.; Tsuge, T.; Abe, H.; Sudesh, K. Potential applications of polyhydroxyalkanoates as a biomaterial for the aging population. Polym. Degrad. Stab. 2020, 181, 109371. [Google Scholar] [CrossRef]
- Tepha Medical Devices Overview. Available online: http://www.tepha.com/technology/overview/ (accessed on 25 March 2022).
- Development of Biodegradable Water Bottle. Available online: http://www.foodpackagingforum.org/news/development-of-biodegradable-water-bottle (accessed on 23 January 2019).
- Wang, Y.; Yin, J.; Chen, G.-Q. Polyhydroxyalkanoates, challenges and opportunities. Curr. Opin. Biotechnol. 2014, 30, 59–65. [Google Scholar] [CrossRef]
- Choonut, A.; Prasertsan, P.; Klomklao, S.; Sangkharak, K. An environmentally friendly process for textile wastewater treatment with a medium-chain-length polyhydroxyalkanoate film. J. Polym. Environ. 2021, 29, 3335–3346. [Google Scholar] [CrossRef]
- Tim, D.S.; Stock, U.; Hrkach, J.; Shinoka, T.; Lien, B.; Moses, M.A.; Stamp, A.; Taylor, G.; Moran, A.M.; Landis, W.; et al. Tissue engineering of autologous aorta using a new biodegradable polymer. Ann. Thorac. Surg. 1999, 68, 2298–2305. [Google Scholar]
- Zhang, L.; Zheng, Z.; Xi, J.; Gao, Y.; Ao, Q.; Gong, Y.; Zhao, N.; Zhang, X. Improved mechanical property and biocompatibility of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) for blood vessel tissue engineering by blending with poly(propylene carbonate). Eur. Polym. J. 2007, 43, 2975–2986. [Google Scholar] [CrossRef]
- Sodian, R.; Hoerstrup, S.P.; Sperling, J.S.; Daebritz, S.; Martin, D.P.; Moran, A.M.; Kim, B.S.; Schoen, F.J.; Vacanti, J.P.; Mayer Jr, J.E. Early in vivo experience with tissue-engineered trileaflet heart valves. Circulation 2007, 102, 22–29. [Google Scholar]
- Stock, U.A.; Nagashima, M.; Kahalil, P.N.; Nollert, G.D.; Herden, T.; Sperling, J.S.; Moran, A.M.; Lien, B.; Martin, D.P.; Schoen, F.J.; et al. Tissue-engineered valved conduits in the pulmonary circulation. J. Thorac. Cardiovasc. Surg. 2000, 119, 732–740. [Google Scholar] [CrossRef] [Green Version]
- Sodian, R.; Loebe, M.; Hein, A.; Martin, D.P.; Hoerstrup, S.P.; Potapov, E.V.; Hausmann, H.; Leuth, T.; Hetzer, R. Application of stereolithography for scaffold fabrication for tissue engineered heart valves. ASAIO J. 2002, 48, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Liu, Y.L.; Cui, B.; Qu, X.H.; Chen, G.Q. Study on decellularized porcine aortic valve/poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) hybrid heart valve in sheep model. Artif. Organs 2007, 31, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, X.; Li, G.; Zhao, N.; Zhang, X. Study on chitosan and PHBHHx used as nerve regeneration conduit material. J. Biomed. Eng. 2002, 19, 25–29. [Google Scholar]
- Ying, H.T.; Ishii, D.; Mahara, A.; Murakami, S.; Yamaoka, T.; Sudesh, K.; Samian, R.; Fujita, M.; Maeda, M.; Iwata, T. Scaffolds from electrospun polyhydroxyalkanoate copolymers: Fabrication, characterization, bioabsorption and tissue response. Biomaterials 2008, 29, 1307–1317. [Google Scholar] [CrossRef]
- Wang, Y.W.; Wu, Q.O.; Chen, G.Q. Attachment, proliferation and differentiation of osteoblasts on random biopolyester poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) scaffolds. Biomaterials 2004, 25, 669–675. [Google Scholar] [CrossRef]
- Zhao, K.; Deng, Y.; Chen, J.C.; Chen, G.Q. Polyhydroxyalkanoate (PHA) scaffolds with good mechanical properties and biocompatibility. Biomaterials 2003, 24, 1041–1054. [Google Scholar] [CrossRef]
- Deng, Y.; Zhao, K.; Zhang, X.F.; Hu, P.; Chen, G.Q. Study on the three-dimensional proliferation of rabbit articular cartilage-derived chondrocytes on polyhydroxyalkanoate scaffolds. Biomaterials 2002, 23, 4049–4056. [Google Scholar] [CrossRef]
- Deng, Y.; Lin, X.J.; Zheng, Z.; Deng, J.G.; Chen, J.C.; Ma, H.; Chen, G.Q. Poly(hydroxybutyrate-co-hydroxyhexanoate) promoted production of extracellular matrix of articular cartilage chondrocytes in vitro. Biomaterials 2003, 24, 4273–4281. [Google Scholar] [CrossRef]
- Wang, Z.; Itoh, Y.; Hosaka, Y.; Kobayashi, I.; Nakano, Y.; Maeda, I.; Umeda, F.; Yamakawa, J.; Kawase, M.; Yagi, K. Novel transdermal drug delivery system with polyhydroxyalkanoate and starburst polyamidoamine dendrimer. J. Biosci. Bioeng. 2003, 95, 541–543. [Google Scholar] [CrossRef]
- Vermeer, C.M.; Rossi, E.; Tamis, J.; Jonkers, H.M.; Kleerebezem, R. From waste to self-healing concrete: A proof-of-concept of a new application for polyhydroxyalkanoate. Resour. Conserv. Recycl. 2021, 164, 105206. [Google Scholar] [CrossRef]
- Haraźna, K.; Cichoń, E.; Skibiński, S.; Witko, T.; Solarz, D.; Kwiecień, I.; Guzik, M. Physicochemical and biological characterisation of diclofenac oligomeric poly (3-hydroxyoctanoate) hybrids as β-TCP ceramics modifiers for bone tissue regeneration. Int. J. Mol. Sci. 2020, 21, 9452. [Google Scholar] [CrossRef]
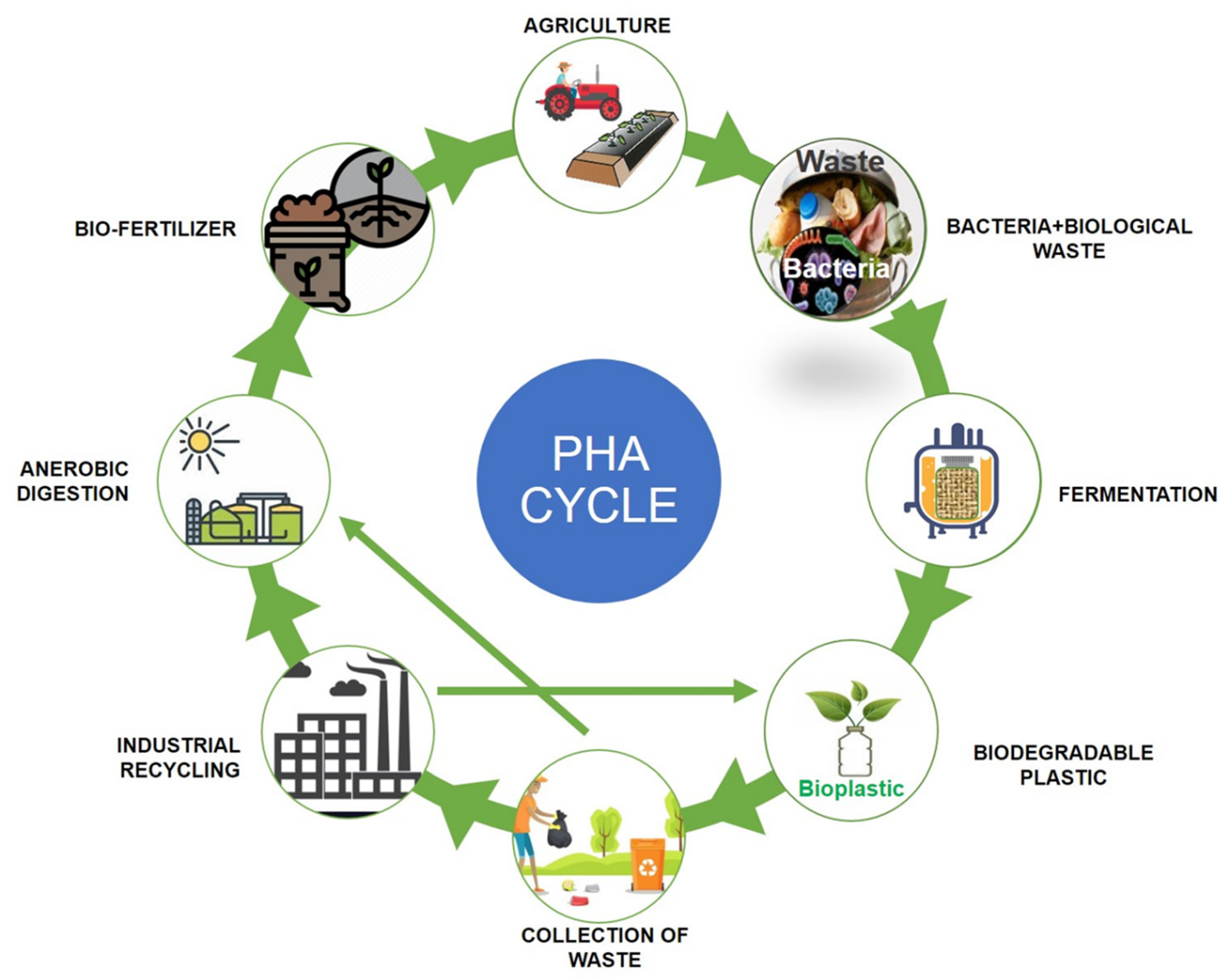
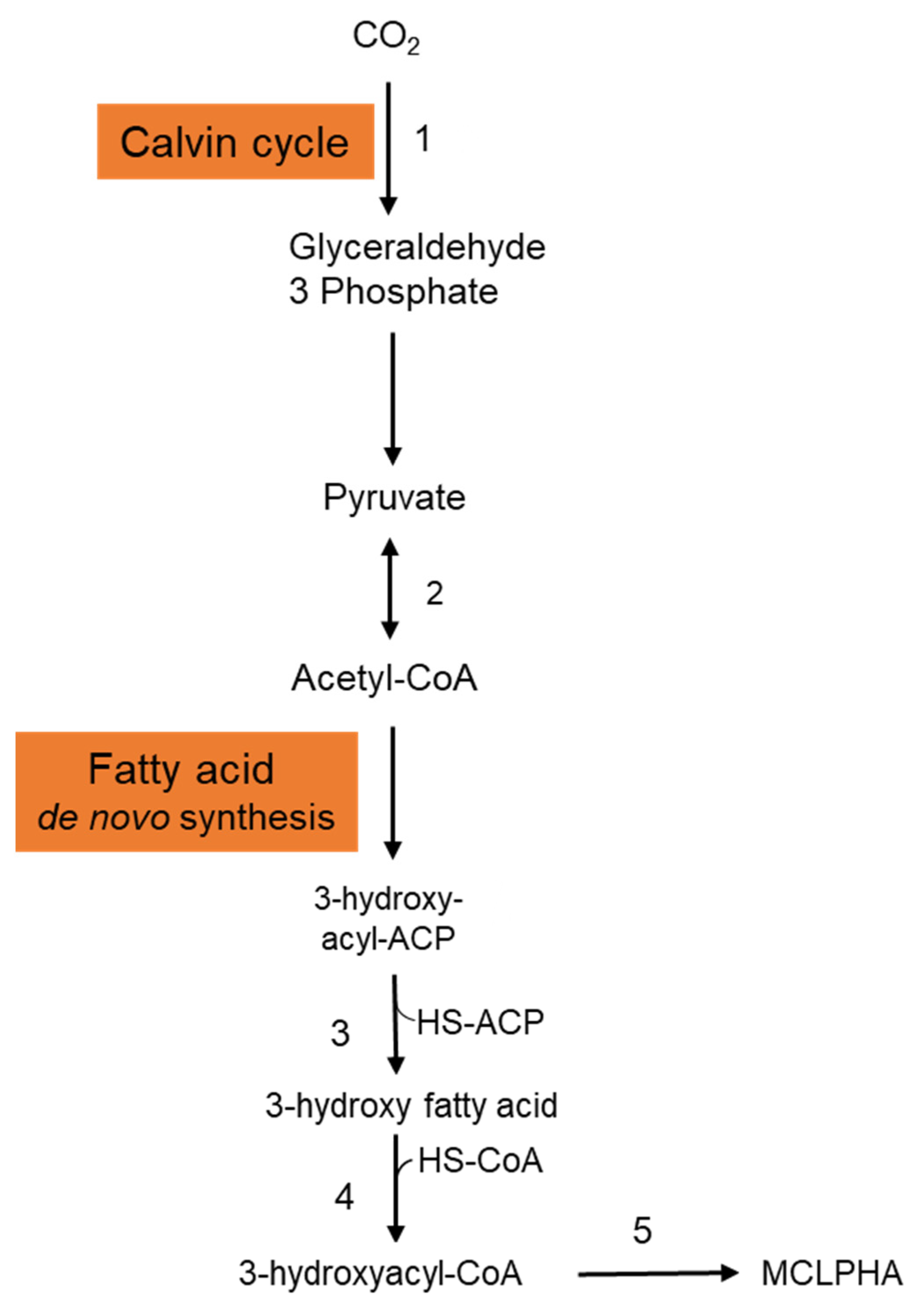
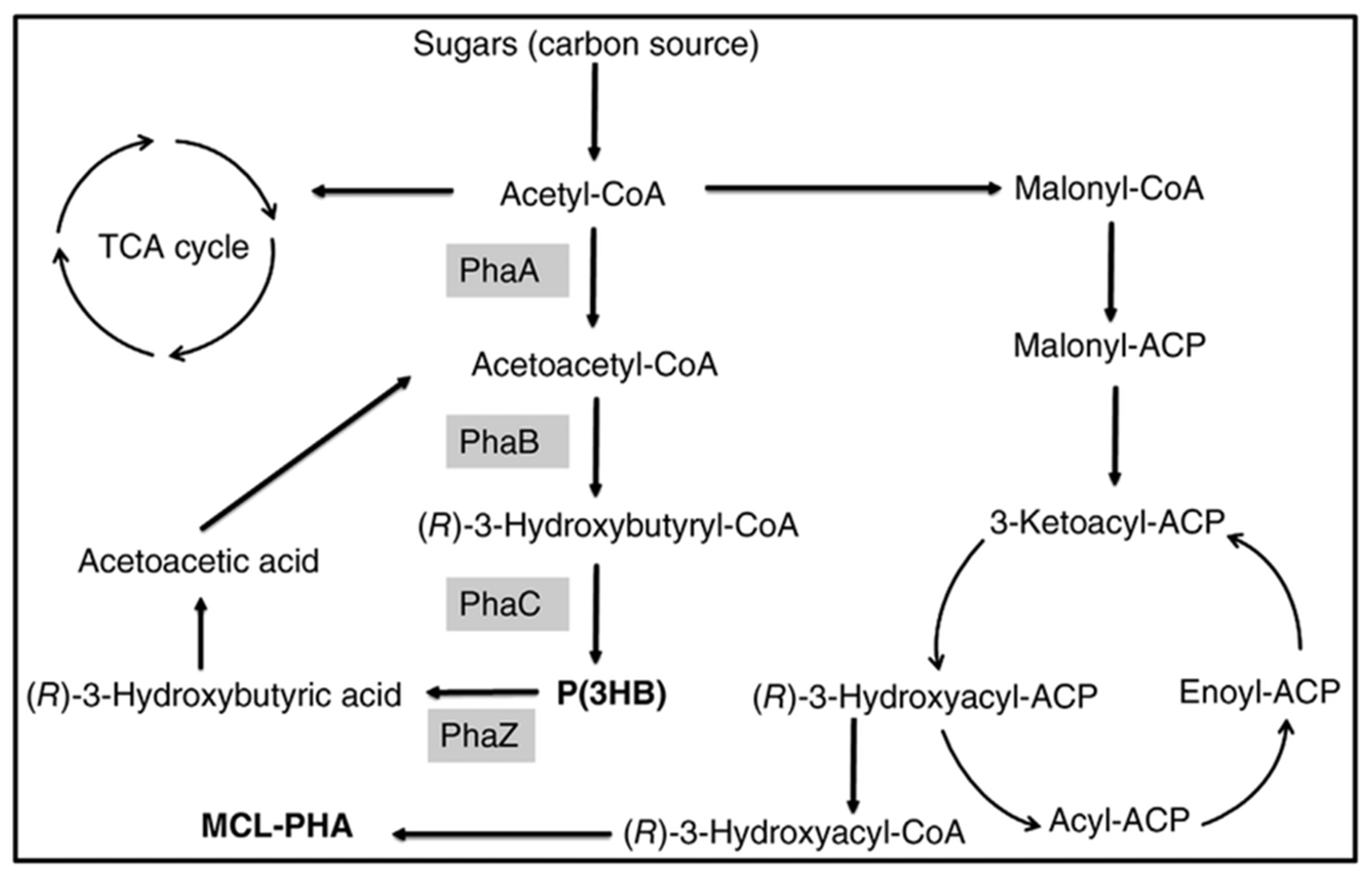


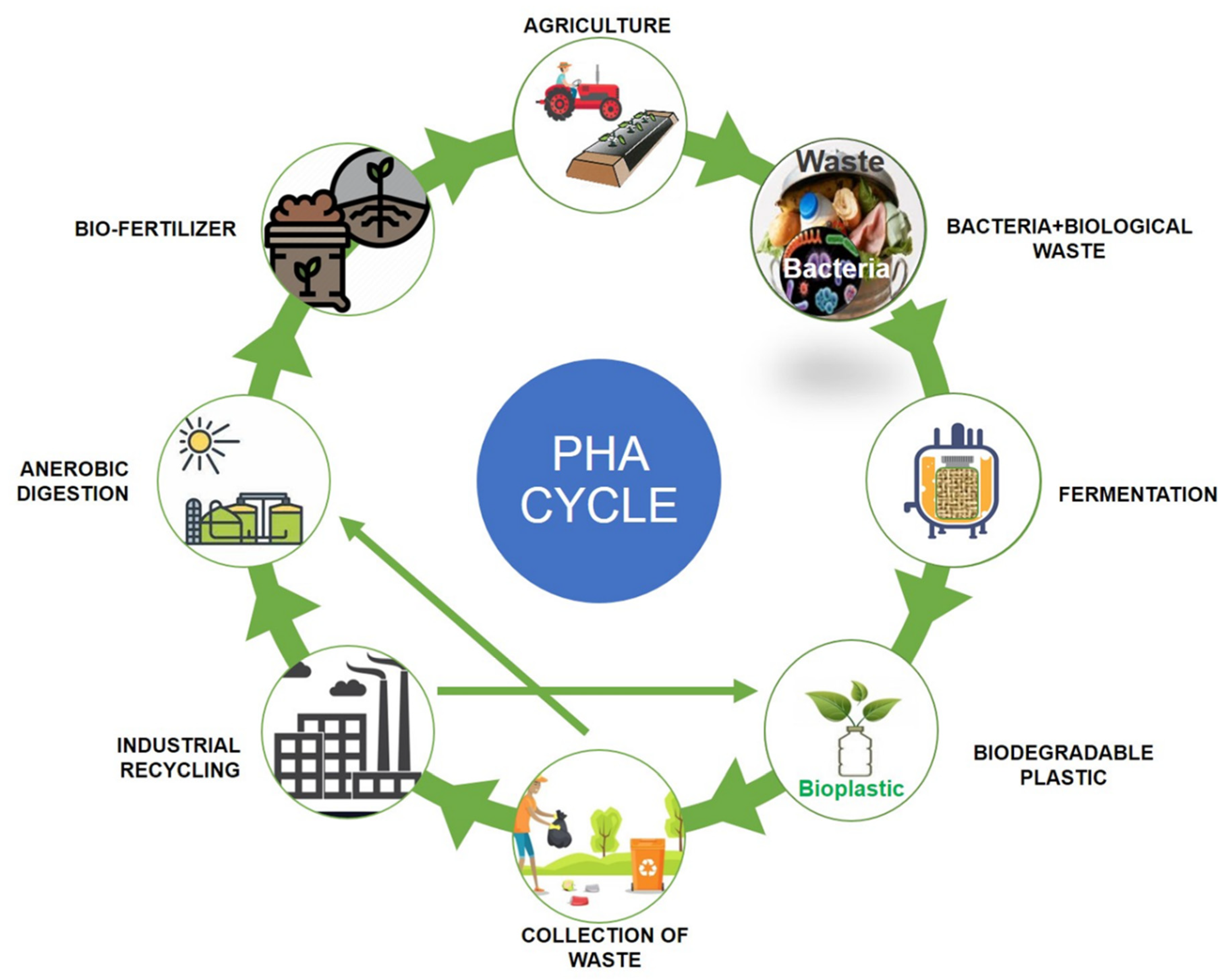{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Carbon Source | DCW (g/L) | PHA Conc. (g/L) | PHA (%) | PHA Composition | Productivity (g/L/h) | Time (h) | References |
|---|---|---|---|---|---|---|---|---|
| P. putida KT2442 | Oleic acid | 141 | 72 | 51 | C6-C8-C10-C12-C14 | 1.91 | 38 | [10] |
| P. putida KT2440 | Glucose + nonanoic acid | 98 | 32 | 33 | C9-C7 | 3.1 | 32 | [13] |
| Recombinant R. eutropha | Palm oil | 139 | 102 | 74 | Mcl-PHA | 1.2 | 96 | [27] |
| P. putida KT2440 | Waste cooking oil | 159 | 58 | 36 | C6-C8-C10-C12-C14 | 1.93 | 30 | [28] |
| P. putida KT2440 | Glucose + nonanoic acid + acrylic acid | 71 | 53 | 75 | C9-C7 | 1.8 | 30 | [29] |
| P. putida LS46 | Octanoic acid | 29 | 17 | 61 | C6-C8-C10-C12 | 0.66 | 27 | [30] |
| P. putida KT2440 | Glucose + nonanoic acid | 71 | 40 | 56 | C5-C7-C9 | 1.44 | 28 | [31] |
| P. putida IPT 046 | Glucose + fructose | 50 | 31 | 63 | Mcl-PHA | 0.8 | 42 | [32] |
| P. oleovorans | n-octane | 112 | 5.6 | 5 | Mcl-PHA | 0.091 | 61 | [33] |
| P. putida KT2442 | Octanoic acid | 51 | 18 | 35 | C8 | 0.41 | 43 | [34] |
| P. putida KT2442 | Oleic acid | 90 | 18 | 20 | C6-C8-C10-C12-C14 | 0.57 | 32 | [34] |
| P. oleovorans | n-octane | 12 | 3.4 | 28 | Mcl-PHA | 0.58 | 120 | [35] |
| P. oleovorans | n-octane | 37 | 12 | 33 | Mcl-PHA | 0.25 | 48 | [36] |
| Bacteria | Substrate | Mode of Operation | PHA Production | Reference |
|---|---|---|---|---|
| P. putida KT2440 | Glucose and nonanoic acid | Fed-batch mode | 32% | [13] |
| P. mendocina | Octanoate | Batch mode | 31% | [82] |
| P. oleovorans | n-octane | Continuous mode | 63% | [83] |
| P. putida | Oleic acid | Fed-batch mode | 51% | [84] |
| P. putida LS46 | Octanoic acid | Fed-batch mode | 61% | [30] |
| P. putida KT2440 | Decanoic and acetic acids | Fed-batch mode | 74% | [85] |
| P. putida KT2440 | Glucose and nonanoic acid | Fed-batch mode | 56% | [31] |
| P. putida CA-3 | Decanoic and butyric acid | Fed-batch mode | 65% | [86] |
| P. putida KT2440 | Nonanoic acid | Fed-batch mode | 75% | [87] |
| P. oleovorans | Octanoic acid | Fed-batch mode | 62% | [88] |
| P. putida BM01 | Glucose and octanoate | Fed-batch mode | 66% | [89] |
| Bacteria | Carbon Source | Mw (kDa) | Mn (kDa) | PDI | References |
|---|---|---|---|---|---|
| P. putida KT2440 | Waste cooking oil | 45 | 22 | 2.04 | [28] |
| P. putida KT2440 | Nonanoic acid + undecanoic acid | 115 | - | 1.8 | [31] |
| P. putida KT2442 | Oleic acid | 135 | 49 | 2.76 | [34] |
| P. putida KT2442 | Octanoic acid | 187 | 78 | 2.4 | [34] |
| P. putida KT2442 | Vegetable-free fatty acids | 168 | 65 | 2.68 | [34] |
| P. putida KT2442 | Animal-free fatty acids | 180 | 71 | 2.53 | [34] |
| P. putida KT2440 | Biodiesel-derived crude glycerol | 462 | 193 | 2.4 | [154] |
| P. putida LS46 | Hexanoic acid | 49 | 22 | 2.3 | [155] |
| P. putida LS46 | Heptanoic acid | 82 | 35 | 2.3 | [155] |
| P. putida LS46 | Octanoic acid | 115 | 54 | 2.2 | [155] |
| P. putida LS46 | Nonanoic acid | 55 | 26 | 2.3 | [155] |
| P. putida LS46 | Decanoic acid | 49 | 21 | 2.4 | [155] |
| P. putida LS46 | Lauric acid | 131 | 63 | 2.1 | [155] |
| P. putida LS46 | Myristic acid | 86 | 44 | 2.0 | [155] |
| P. putida KT2442 mutant | Dodecanoic acid (15%) | 100 | 80 | 1.25 | [156] |
| P. putida KT2442 mutant | Dodecanoic acid (39%) | 157 | 108 | 1.45 | [156] |
| P. putida KT2442 mutant | Tetradecanoic acid (49%) | 95 | 67 | 1.43 | [156] |
| P. corrugat | Coconut oil | 343 | 74 | 4.6 | [157] |
| P. resinovorans | Coconut oil | 165 | 101 | 1.63 | [158] |
| P. resinovorans | Sunflower oil | 112 | 65 | 1.72 | [158] |
| P. resinovorans | Soybean oil | 127 | 70 | 1.81 | [158] |
| P. oleovorans | Octanoic acid | 189 | 51 | 3.69 | [159] |
| P. oleovorans | Soybean oil | 130 | 72 | 1.70 | [159] |
| P. oleovorans | Undecanoic acid | 260 | 135 | 1.92 | [159] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reddy, V.U.N.; Ramanaiah, S.V.; Reddy, M.V.; Chang, Y.-C. Review of the Developments of Bacterial Medium-Chain-Length Polyhydroxyalkanoates (mcl-PHAs). Bioengineering 2022, 9, 225. https://doi.org/10.3390/bioengineering9050225
Reddy VUN, Ramanaiah SV, Reddy MV, Chang Y-C. Review of the Developments of Bacterial Medium-Chain-Length Polyhydroxyalkanoates (mcl-PHAs). Bioengineering. 2022; 9(5):225. https://doi.org/10.3390/bioengineering9050225
Chicago/Turabian StyleReddy, V. Uttej Nandan, S. V. Ramanaiah, M. Venkateswar Reddy, and Young-Cheol Chang. 2022. "Review of the Developments of Bacterial Medium-Chain-Length Polyhydroxyalkanoates (mcl-PHAs)" Bioengineering 9, no. 5: 225. https://doi.org/10.3390/bioengineering9050225
APA StyleReddy, V. U. N., Ramanaiah, S. V., Reddy, M. V., & Chang, Y.-C. (2022). Review of the Developments of Bacterial Medium-Chain-Length Polyhydroxyalkanoates (mcl-PHAs). Bioengineering, 9(5), 225. https://doi.org/10.3390/bioengineering9050225