
Perfusion of MC3T3E1 Preosteoblast Spheroids within Polysaccharide-Based Hydrogel Scaffolds: An Experimental and Numerical Study at the Bioreactor Scale

 ,
,  ,
,  and
and
Abstract
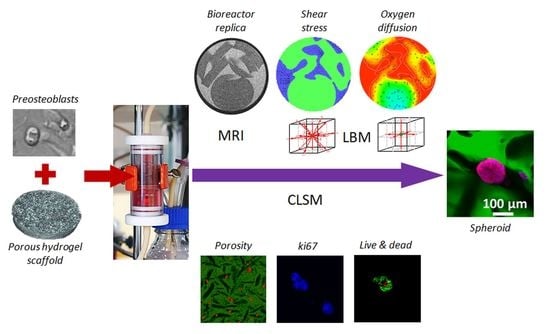
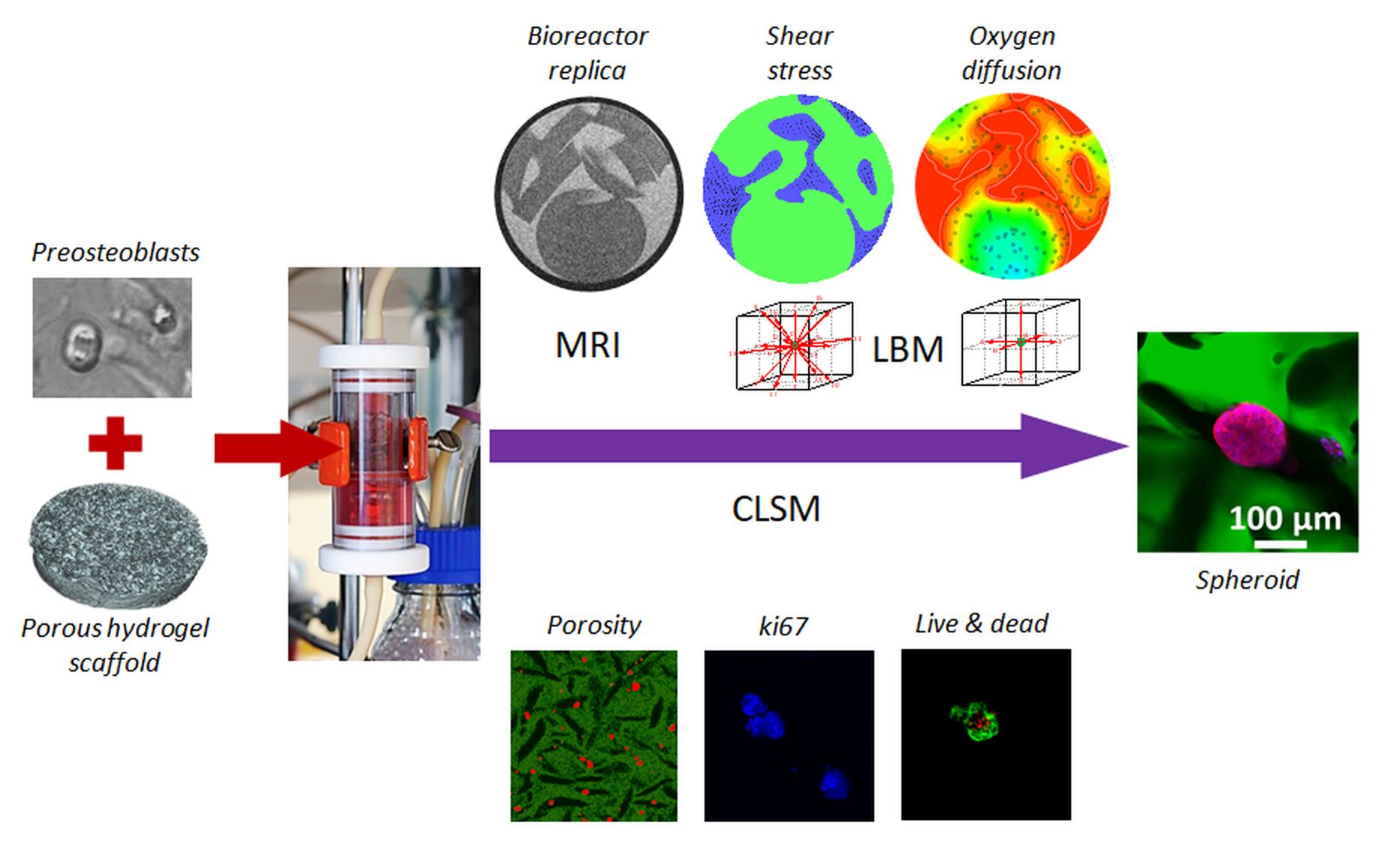{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Porous Polysaccharide-Based Hydrogel Scaffolds Preparation
2.2. MC3T3E1 Cell Seeding in Porous Hydrogel Scaffolds
2.3. 3D Dynamic Culture in a Perfusion Bioreactor
2.4. Evaluation of Cell Number, Division, and Viability by CLSM
2.4.1. Cell Number per Scaffold
2.4.2. Cell Proliferation
2.4.3. Cell Viability Assessment
2.5. MRI Acquisitions
2.5.1. Cell Labelling with Superparamagnetic Iron Oxide Nanoparticles
2.5.2. MRI Sequences
2.6. Measurement of the Oxygen Diffusion Coefficient inside the Hydrogel
2.6.1. Experimental Setup
2.6.2. Measurement Protocol
2.6.3. Identification of the Oxygen Diffusion Coefficient
2.7. Computational Fluid Dynamic (CFD) Simulation of the Perfusion Bioreactor
2.7.1. Modelling and Lattice Boltzmann Method Implementation
2.7.2. Simulation of a Segment of the Perfusion Bioreactor
2.8. Simulation of the Oxygen Transport in the Hydrogel
2.8.1. Modelling and LBM Implementation
2.8.2. Case Studies
- Oxygen transport at the scaffold scale.
- b
- Oxygen transport at the perfusion bioreactor scale.
2.9. Data Analysis
3. Results
3.1. Dynamic vs. Static 3D Cell Culture Conditions
3.1.1. Assessment of Cell Number
3.1.2. Assessment of Cell Proliferation
3.1.3. Assessment of Cell Viability
3.2. Assessment of Cell Number by MRI within the Porous Hydrogel Scaffolds
3.3. Oxygen Diffusion Coefficient within the Hydrogel
3.4. Digitalization of the Bioreactors by MRI Acquisitions
3.5. Numerical Simulations of Hydrodynamics and Oxygen Transport Using LB Methods
3.5.1. Perfusion Flow through the Stack Acquired by MRI
3.5.2. Effect of Spheroid Size and Number on Oxygen Level in the Scaffold
3.5.3. Oxygen Transport at the Perfusion Bioreactor Scale
4. Discussion
4.1. 3D Dynamic Cell Culture Conditions
4.2. Perfusion Bioreactor Modelling Based on Experimental Data
4.2.1. The Issues of Imaging the Hierarchically Porous Bioreactor
4.2.2. Hydrodynamics and Dissolved Oxygen Transport Simulations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fennema, E.; Rivron, N.; Rouwkema, J.; van Blitterswijk, C.; de Boer, J. Spheroid culture as a tool for creating 3D complex tissues. Trends Biotechnol. 2013, 31, 108–115. [Google Scholar] [CrossRef]
- Hofer, M.; Lutolf, M.P. Engineering organoids. Nat. Rev. Mater. 2021, 6, 402–420. [Google Scholar] [CrossRef]
- Griffith, L.G.; Swartz, M.A. Capturing complex 3D tissue physiology in vitro. Nat. Rev. Mol. Cell Biol. 2006, 7, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Mammoto, T.; Ingber, D.E. Mechanical control of tissue and organ development. Dev. Camb. Engl. 2010, 137, 1407–1420. [Google Scholar] [CrossRef] [PubMed]
- Vianello, S.; Lutolf, M.P. Understanding the Mechanobiology of Early Mammalian Development through Bioengineered Models. Dev. Cell 2019, 48, 751–763. [Google Scholar] [CrossRef] [PubMed]
- Chabanon, M.; Duval, H.; Grenier, J.; Beauchesne, C.; Goyeau, B.; David, B. Histological Method to Study the Effect of Shear Stress on Cell Proliferation and Tissue Morphology in a Bioreactor. Tissue Eng. Regen. Med. 2019, 16, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Wittkowske, C.; Reilly, G.C.; Lacroix, D.; Perrault, C.M. In Vitro Bone Cell Models: Impact of Fluid Shear Stress on Bone Formation. Front. Bioeng. Biotechnol. 2016, 4, 87. [Google Scholar] [CrossRef]
- Bae, H.; Ahari, A.F.; Shin, H.; Nichol, J.W.; Hutson, C.B.; Masaeli, M.; Kim, S.-H.; Aubin, H.; Yamanlar, S.; Khademhosseini, A. Cell-laden microengineered pullulan methacrylate hydrogels promote cell proliferation and 3D cluster formation. Soft Matter. 2011, 7, 1903. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, J.; Oliveira, H.; Aid, R.; Bareille, R.; Siadous, R.; Letourneur, D.; Mao, Y.; Kohn, J.; Amédée, J. Influence of the three-dimensional culture of human bone marrow mesenchymal stromal cells within a macroporous polysaccharides scaffold on Pannexin 1 and Pannexin 3. J. Tissue Eng. Regen. Med. 2018, 12, e1936–e1949. [Google Scholar] [CrossRef]
- Pietrzyk-Nivau, A.; Poirault-Chassac, S.; Gandrille, S.; Derkaoui, S.-M.; Kauskot, A.; Letourneur, D.; Visage, C.L.; Baruch, D. Three-Dimensional Environment Sustains Hematopoietic Stem Cell Differentiation into Platelet-Producing Megakaryocytes. PLoS ONE 2015, 10, e0136652. [Google Scholar] [CrossRef]
- Fricain, J.C.; Aid, R.; Lanouar, S.; Maurel, D.B.; Le Nihouannen, D.; Delmond, S.; Letourneur, D.; Vilamitjana, J.A.; Catros, S. In-vitro and in-vivo design and validation of an injectable polysaccharide-hydroxyapatite composite material for sinus floor augmentation. Dent. Mater. 2018, 34, 1024–1035. [Google Scholar] [CrossRef]
- Johnson, C.; Natarajan, V.; Antoniou, C. Verification of energy dissipation rate scalability in pilot and production scale bioreactors using computational fluid dynamics. Biotechnol. Prog. 2014, 30, 760–764. [Google Scholar] [CrossRef] [PubMed]
- Loubière, C.; Delafosse, A.; Guedon, E.; Chevalot, I.; Toye, D.; Olmos, E. Dimensional analysis and CFD simulations of microcarrier ‘just-suspended’ state in mesenchymal stromal cells bioreactors. Chem. Eng. Sci. 2019, 203, 464–474. [Google Scholar] [CrossRef]
- Sarkar, J.; Shekhawat, L.K.; Loomba, V.; Rathore, A.S. CFD of Mixing of Multi-Phase Flow in a Bioreactor Using Population Balance Model. Biotechnol. Prog. 2016, 32, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Kerdouss, F.; Bannari, A.; Proulx, P.; Bannari, R.; Skrga, M.; Labrecque, Y. Two-phase mass transfer coefficient prediction in stirred vessel with a CFD model. Comput. Chem. Eng. 2008, 32, 1943–1955. [Google Scholar] [CrossRef]
- Amer, M.; Feng, Y.; Ramsey, J.D. Using CFD simulations and statistical analysis to correlate oxygen mass transfer coefficient to both geometrical parameters and operating conditions in a stirred-tank bioreactor. Biotechnol. Prog. 2019, 35, e2785. [Google Scholar] [CrossRef]
- Zajec, V.E.; Novak, U.; Kastelic, M.; Japelj, B.; Lah, L.; Pohar, A.; Likozar, B. Dynamic multiscale metabolic network modeling of Chinese hamster ovary cell metabolism integrating N-linked glycosylation in industrial biopharmaceutical manufacturing. Biotechnol. Bioeng. 2021, 118, 397–411. [Google Scholar] [CrossRef]
- György, R.; Kostoglou, M.; Mantalaris, A.; Georgiadis, M.C. Development of a multi-scale model to simulate mesenchymal stem cell osteogenic differentiation within hydrogels in a rotating wall bioreactor. Biochem. Eng. J. 2022, 186, 108566. [Google Scholar] [CrossRef]
- Porter, B.; Zauel, R.; Stockman, H.; Guldberg, R.; Fyhrie, D. 3-D computational modeling of media flow through scaffolds in a perfusion bioreactor. J. Biomech. 2005, 38, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Sayed, A.A.M.; Hussein, M.A.; Becker, T. An innovative lattice Boltzmann model for simulating Michaelis–Menten-based diffusion–advection kinetics and its application within a cartilage cell bioreactor. Biomech. Model. Mechanobiol. 2010, 9, 141–151. [Google Scholar] [CrossRef]
- Sacco, R.; Causin, P.; Zunino, P.; Raimondi, M.T. A multiphysics/multiscale 2D numerical simulation of scaffold-based cartilage regeneration under interstitial perfusion in a bioreactor. Biomech. Model. Mechanobiol. 2011, 10, 577–589. [Google Scholar] [CrossRef]
- Hossain, M.S.; Bergstrom, D.J.; Chen, X.B. Prediction of cell growth rate over scaffold strands inside a perfusion bioreactor. Biomech. Model. Mechanobiol. 2015, 14, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Mehrian, M.; Lambrechts, T.; Papantoniou, I.; Geris, L. Computational Modeling of Human Mesenchymal Stromal Cell Proliferation and Extra-Cellular Matrix Production in 3D Porous Scaffolds in a Perfusion Bioreactor: The Effect of Growth Factors. Front. Bioeng. Biotechnol. 2020, 8, 376. [Google Scholar] [CrossRef] [PubMed]
- Fricain, J.C.; Schlaubitz, S.; Le Visage, C.; Arnault, I.; Derkaoui, S.M.; Siadous, R.; Catros, S.; Lalande, C.; Bareille, R.; Renard, M.; et al. A nano-hydroxyapatite—Pullulan/dextran polysaccharide composite macroporous material for bone tissue engineering. Biomaterials 2013, 34, 2947–2959. [Google Scholar] [CrossRef]
- Czekanska, E.M.; Stoddart, M.J.; Ralphs, J.R.; Richards, R.G.; Hayes, J.S. A phenotypic comparison of osteoblast cell lines versus human primary osteoblasts for biomaterials testing: Phenotypic Comparison of Osteoblast Cell Lines. J. Biomed. Mater. Res. A 2014, 102, 2636–2643. [Google Scholar] [CrossRef] [PubMed]
- Quarles, L.D.; Yohay, D.A.; Lever, L.W.; Caton, R.; Wenstrup, R.J. Distinct proliferative and differentiated stages of murine MC3T3-E1 cells in culture: An in vitro model of osteoblast development. J. Bone Miner. Res. 1992, 7, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Zaloga, J.; Pöttler, M.; Leitinger, G.; Friedrich, R.P.; Almer, G.; Lyer, S.; Baum, E.; Tietze, R.; Heimke-Brinck, R.; Mangge, H.; et al. Pharmaceutical formulation of HSA hybrid coated iron oxide nanoparticles for magnetic drug targeting. Eur. J. Pharm. Biopharm. 2016, 101, 152–162. [Google Scholar] [CrossRef]
- Potier, E.; Ferreira, E.; Andriamanalijaona, R.; Pujol, J.-P.; Oudina, K.; Logeart-Avramoglou, D.; Petite, H. Hypoxia affects mesenchymal stromal cell osteogenic differentiation and angiogenic factor expression. Bone 2007, 40, 1078–1087. [Google Scholar] [CrossRef]
- Muschler, G.F.; Nakamoto, C.; Griffith, L.G. Engineering Principles of Clinical Cell-based Tissue Engineering. J. Bone Joint Surg.-Am. Vol. 2004, 86, 1541–1558. [Google Scholar] [CrossRef]
- Xu, H.; Othman, S.F.; Magin, R.L. Monitoring Tissue Engineering Using Magnetic Resonance Imaging. J. Biosci. Bioeng. 2008, 106, 515–527. [Google Scholar] [CrossRef]
- Grenier, J.; Duval, H.; Barou, F.; Lv, P.; David, B.; Letourneur, D. Mechanisms of pore formation in hydrogel scaffolds textured by freeze-drying. Acta Biomater. 2019, 94, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.K.; Contera, S.A.; Justesen, J.; Duch, M.; Hansen, O.; Chevallier, J.; Foss, M.; Pedersen, F.; Besenbacher, F. Cell volume increase in murine MC3T3-E1 pre-osteoblasts attaching onto biocompatible Tantalum observed by magnetic AC mode Atomic Force Microscopy. Eur. Cell. Mater. 2005, 10, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Milford, D.; Rosbach, N.; Bendszus, M.; Heiland, S. Mono-Exponential Fitting in T2-Relaxometry: Relevance of Offset and First Echo. PLoS ONE 2015, 10, e0145255. [Google Scholar] [CrossRef] [PubMed]
- Hulst, A.C.; Hens, H.J.H.; Buitelaar, R.M.; Tramper, J. Determination of the effective diffusion coefficient of oxygen in gel materials in relation to gel concentration. Biotechnol. Tech. 1989, 3, 199–204. [Google Scholar] [CrossRef]
- Carman, P.C.; Haul, R.A. Measurement of diffusion coefficients. Proc. R. Soc. Lond Ser. Math. Phys. Sci. 1954, 222, 109–118. [Google Scholar] [CrossRef]
- Lakhotia, S.; Papoutsakis, E.T. Agitation induced cell injury in microcarrier cultures. Protective effect of viscosity is agitation intensity dependent: Experiments and modeling. Biotechnol. Bioeng. 1992, 39, 95–107. [Google Scholar] [CrossRef]
- d’Humières, D. Multiple–relaxation–time lattice Boltzmann models in three dimensions. Philos. Trans. R. Soc. Lond Ser. Math. Phys. Eng. Sci. 2002, 360, 437–451. [Google Scholar] [CrossRef]
- Duval, H.; Masson, D.; Guillot, J.-B.; Schmitz, P.; d’Humières, D. Two-dimensional lattice-Boltzmann model of hydrosol depth filtration. AICHE J. 2006, 52, 39–48. [Google Scholar] [CrossRef]
- Thibeaux, R.; Duval, H.; Smaniotto, B.; Vennat, E.; Néron, D.; David, B. Assessment of the interplay between scaffold geometry, induced shear stresses, and cell proliferation within a packed bed perfusion bioreactor. Biotechnol. Prog. 2019, 35, e2880. [Google Scholar] [CrossRef]
- Ginzburg, I.; d’Humières, D. Multireflection boundary conditions for lattice Boltzmann models. Phys. Rev. E 2003, 68, 066614. [Google Scholar] [CrossRef]
- Stahl, B.; Chopard, B.; Latt, J.J. Measurements of wall shear stress with the lattice Boltzmann method and staircase approximation of boundaries. Comput. Fluids 2010, 39, 1625–1633. [Google Scholar] [CrossRef]
- Kunz-Schughart, L.A.; Doetsch, J.; Mueller-Klieser, W.; Groebe, K. Proliferative activity and tumorigenic conversion: Impact on cellular metabolism in 3-D culture. Am. J. Physiol.-Cell Physiol. 2000, 278, C765–C780. [Google Scholar] [CrossRef]
- Cho, C.H.; Park, J.; Nagrath, D.; Tilles, A.W.; Berthiaume, F.; Toner, M.; Yarmush, M.L. Oxygen uptake rates and liver-specific functions of hepatocyte and 3T3 fibroblast co-cultures. Biotechnol. Bioeng. 2007, 97, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Ginzburg, I. Prediction of the moments in advection-diffusion lattice Boltzmann method. I. Truncation dispersion, skewness, and kurtosis. Phys. Rev. E 2017, 95, 013304. [Google Scholar] [CrossRef] [PubMed]
- Pothirajan, P.; Dorcemus, D.; Nukavarapu, S.; Kotecha, M. True MRI assessment of stem cell chondrogenesis in a tissue engineered matrix. In Proceedings of the 2014 36th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Chicago, IL, USA, 26–30 August 2014; pp. 3933–3936. [Google Scholar]
- Hidalgo-Bastida, L.A.; Cartmell, S.H.; Cells, M.S. Osteoblasts and Extracellular Matrix Proteins: Enhancing Cell Adhesion and Differentiation for Bone Tissue Engineering. Tissue Eng. Part B Rev. 2010, 16, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Samavedi, S.; Whittington, A.R.; Goldstein, A.S. Calcium phosphate ceramics in bone tissue engineering: A review of properties and their influence on cell behavior. Acta Biomater. 2013, 9, 8037–8045. [Google Scholar] [CrossRef]
- Napolitano, A.P.; Chai, P.; Dean, D.M.; Morgan, J.R. Dynamics of the Self-Assembly of Complex Cellular Aggregates on Micromolded Nonadhesive Hydrogels. Tissue Eng. 2007, 13, 2087–2094. [Google Scholar] [CrossRef] [PubMed]
- Brassard, J.A.; Lutolf, M.P. Engineering Stem Cell Self-organization to Build Better Organoids. Cell Stem Cell 2019, 24, 860–876. [Google Scholar] [CrossRef] [PubMed]
- Fujimichi, Y.; Otsuka, K.; Tomita, M.; Iwasaki, T. An Efficient Intestinal Organoid System of Direct Sorting to Evaluate Stem Cell Competition in Vitro. Sci. Rep. 2019, 9, 20297. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Renner, M.; Martin, C.-A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral organoids model human brain development and microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef]
- Qian, X.; Nguyen, H.N.; Song, M.M.; Hadiono, C.; Ogden, S.C.; Hammack, C.; Yao, B.; Hamersky, G.R.; Jacob, F.; Zhong, C.; et al. Brain-Region-Specific Organoids Using Mini-bioreactors for Modeling ZIKV Exposure. Cell 2016, 165, 1238–1254. [Google Scholar] [CrossRef]
- Clevers, H. Modeling Development and Disease with Organoids. Cell 2016, 165, 1586–1597. [Google Scholar] [CrossRef]
- Fatehullah, A.; Tan, S.H.; Barker, N. Organoids as an in vitro model of human development and disease. Nat. Cell Biol. 2016, 18, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Grenier, J.; Duval, H.; Lv, P.; Barou, F.; Le Guilcher, C.; Aid, R.; David, B.; Letourneur, D. Interplay between Crosslinking Degree and Ice Nucleation Controls the Porous Structure of Freeze-Dried Hydrogel Scaffolds. Biomater. Adv. 2022, 139, 212973. [Google Scholar] [CrossRef] [PubMed]
- David, B.; Dore, E.; Jaffrin, M.; Legallais, C. Mass transfers in a fluidized bed bioreactor using alginate beads for a future bioartificial liver. Int. J. Artif. Organs 2004, 27, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Rauh, J.; Milan, F.; Günther, K.-P.; Stiehler, M. Bioreactor Systems for Bone Tissue Engineering. Tissue Eng. Part B Rev. 2011, 17, 263–280. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Kassab, G.S. Role of shear stress and stretch in vascular mechanobiology. J. R. Soc. Interface 2011, 8, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Homan, K.A.; Gupta, N.; Kroll, K.T.; Kolesky, D.B.; Skylar-Scott, M.; Miyoshi, T.; Mau, D.; Valerius, M.T.; Ferrante, T.; Bonventre, J.V.; et al. Flow-enhanced vascularization and maturation of kidney organoids in vitro. Nat. Methods 2019, 16, 255–262. [Google Scholar] [CrossRef]
- Kapur, S.; Baylink, D.J.; Lau, K.-H.W. Fluid flow shear stress stimulates human osteoblast proliferation and differentiation through multiple interacting and competing signal transduction pathways. Bone 2003, 32, 241–251. [Google Scholar] [CrossRef]
- Yu, X.; Botchwey, E.A.; Levine, E.M.; Pollack, S.R.; Laurencin, C.T. Bioreactor-based bone tissue engineering: The influence of dynamic flow on osteoblast phenotypic expression and matrix mineralization. Proc. Natl. Acad. Sci. USA 2004, 101, 11203–11208. [Google Scholar] [CrossRef]
- Bancroft, G.N.; Sikavitsas, V.I.; van den Dolder, J.; Sheffield, T.L.; Ambrose, C.G.; Jansen, J.A.; Mikos, A.G. Fluid flow increases mineralized matrix deposition in 3D perfusion culture of marrow stromal osteoblasts in a dose-dependent manner. Proc. Natl. Acad. Sci. USA 2002, 99, 12600–12605. [Google Scholar] [CrossRef] [PubMed]
- Sikavitsas, V.I.; Bancroft, G.N.; Holtorf, H.L.; Jansen, J.A.; Mikos, A.G. Mineralized matrix deposition by marrow stromal osteoblasts in 3D perfusion culture increases with increasing fluid shear forces. Proc. Natl. Acad. Sci. USA 2003, 100, 14683–14688. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.A.; Chen, C.W.; Chen, C.P.; Tseng, C.S. Enhancement of cell growth in tissue-engineering constructs under direct perfusion: Modeling and simulation. Biotechnol. Bioeng. 2007, 97, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, D.P.; Parker, T.L.; Walker, D.A.; Alexander, C.; Ashford, M.B.; Gellert, P.R.; Garnett, M.C.; Volume, M.S. Resazurin and Acid Phosphatase Viability Assays for High-Throughput Screening of Tumour Spheroids and Stem Cell Neurospheres. PLoS ONE 2014, 9, e103817. [Google Scholar] [CrossRef]
- Gombotz, W.R.; Guanghui, W.; Horbett, T.A.; Hoffman, A.S. Protein adsorption to poly(ethylene oxide) surfaces. J. Biomed. Mater. Res. 1991, 25, 1547–1562. [Google Scholar] [CrossRef]
- Rekha, M.R.; Chandra, P.S. Pullulan as a Promising Biomaterial for Biomedical Applications: A Perspective. Trends Biomater. Artif. Organs 2007, 20, 116–121. [Google Scholar]
- Autissier, A.; Letourneur, D.; Le Visage, C. Pullulan-based hydrogel for smooth muscle cell culture. J. Biomed. Mater. Res. A 2007, 82A, 336–342. [Google Scholar] [CrossRef]
- Thébaud, N.-B.; Pierron, D.; Bareille, R.; Le Visage, C.; Letourneur, D.; Bordenave, L. Human endothelial progenitor cell attachment to polysaccharide-based hydrogels: A pre-requisite for vascular tissue engineering. J. Mater. Sci. Mater. Med. 2007, 18, 339–345. [Google Scholar] [CrossRef]
- Frangos, J.A.; McIntire, L.V.; Eskin, S.G. Shear stress induced stimulation of mammalian cell metabolism. Biotechnol. Bioeng. 1988, 32, 1053–1060. [Google Scholar] [CrossRef]
- Liu, B.; Han, S.; Hedrick, B.P.; Modarres-Sadeghi, Y.; Lynch, M.E. Perfusion applied to a 3D model of bone metastasis results in uniformly dispersed mechanical stimuli. Biotechnol. Bioeng. 2018, 115, 1076–1085. [Google Scholar] [CrossRef]
- Carman, P.C. Permeability of saturated sands, soils and clays. J. Agric. Sci. 1939, 29, 262–273. [Google Scholar] [CrossRef]
- David, B.; Bonnefont-Rousselot, D.; Oudina, K.; Degat, M.-C.; Deschepper, M.; Viateau, V.; Bensidhoum, M.; Oddou, C.; Petite, H. A Perfusion Bioreactor for Engineering Bone Constructs: An In Vitro and In Vivo Study. Tissue Eng. Part C Methods 2011, 17, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Bouet, G.; Marchat, D.; Cruel, M.; Malaval, L.; Vico, L. In vitro three-dimensional bone tissue models: From cells to controlled and dynamic environment. Tissue Eng. Part B Rev. 2015, 21, 133–156. [Google Scholar] [CrossRef] [PubMed]
- Voronov, R.; VanGordon, S.; Sikavitsas, V.I.; Papavassiliou, D.V. Computational modeling of flow-induced shear stresses within 3D salt-leached porous scaffolds imaged via micro-CT. J. Biomech. 2010, 43, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Liegibel, U.; Sommer, U.; Bundschuh, B.; Schweizer, B.; Hilscher, U.; Lieder, A.; Nawroth, P.; Kasperk, C. Fluid Shear of Low Magnitude Increases Growth and Expression of TGFβ1 and Adhesion Molecules in Human Bone Cells in Vitro. Exp. Clin. Endocrinol. Diabetes 2004, 112, 356–363. [Google Scholar] [CrossRef]
- Leclerc, E.; David, B.; Griscom, L.; Lepioufle, B.; Fujii, T.; Layrolle, P.; Legallaisa, C. Study of osteoblastic cells in a microfluidic environment. Biomaterials 2006, 27, 586–595. [Google Scholar] [CrossRef]
- Chhabra, M.; Prausnitz, J.M.; Radke, C.J. Polarographic Method for Measuring Oxygen Diffusivity and Solubility in Water-Saturated Polymer Films: Application to Hypertransmissible Soft Contact Lense. Ind. Eng. Chem. Res. 2008, 47, 3540–3550. [Google Scholar] [CrossRef]
- McClelland, R.E.; MacDonald, J.M.; Coger, R.N. Modeling O2 transport within engineered hepatic devices. Biotechnol. Bioeng. 2003, 82, 12–27. [Google Scholar] [CrossRef]
- Glicklis, R.; Merchuk, J.C.; Cohen, S. Modeling mass transfer in hepatocyte spheroids via cell viability, spheroid size, and hepatocellular functions. Biotechnol. Bioeng. 2004, 86, 672–680. [Google Scholar] [CrossRef]
- Bhise, N.S.; Manoharan, V.; Massa, S.; Tamayol, A.; Ghaderi, M.; Miscuglio, M.; Lang, Q.; Zhang, Y.S.; Shin, S.R.; Calzone, G.; et al. A liver-on-a-chip platform with bioprinted hepatic spheroids. Biofabrication 2016, 8, 014101. [Google Scholar] [CrossRef]
- Sharifi, F.; Firoozabadi, B.; Firoozbakhsh, K. Numerical Investigations of Hepatic Spheroids Metabolic Reactions in a Perfusion Bioreactor. Front. Bioeng. Biotechnol. 2019, 7, 221. [Google Scholar] [CrossRef] [PubMed]
- Seguin, D.; Montillet, A.; Brunjail, D.; Comiti, J. Liquid—Solid mass transfer in packed beds of variously shaped particles at low Reynolds numbers: Experiments and model. Chem. Eng. J. Biochem. Eng. J. 1996, 63, 1–9. [Google Scholar] [CrossRef]
- Lee, J.; Cuddihy, M.J.; Cater, G.M.; Kotov, N.A. Engineering liver tissue spheroids with inverted colloidal crystal scaffolds. Biomaterials 2009, 30, 4687–4694. [Google Scholar] [CrossRef] [PubMed]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grenier, J.; David, B.; Journé, C.; Cicha, I.; Letourneur, D.; Duval, H. Perfusion of MC3T3E1 Preosteoblast Spheroids within Polysaccharide-Based Hydrogel Scaffolds: An Experimental and Numerical Study at the Bioreactor Scale. Bioengineering 2023, 10, 849. https://doi.org/10.3390/bioengineering10070849
Grenier J, David B, Journé C, Cicha I, Letourneur D, Duval H. Perfusion of MC3T3E1 Preosteoblast Spheroids within Polysaccharide-Based Hydrogel Scaffolds: An Experimental and Numerical Study at the Bioreactor Scale. Bioengineering. 2023; 10(7):849. https://doi.org/10.3390/bioengineering10070849
Chicago/Turabian StyleGrenier, Jérôme, Bertrand David, Clément Journé, Iwona Cicha, Didier Letourneur, and Hervé Duval. 2023. "Perfusion of MC3T3E1 Preosteoblast Spheroids within Polysaccharide-Based Hydrogel Scaffolds: An Experimental and Numerical Study at the Bioreactor Scale" Bioengineering 10, no. 7: 849. https://doi.org/10.3390/bioengineering10070849
APA StyleGrenier, J., David, B., Journé, C., Cicha, I., Letourneur, D., & Duval, H. (2023). Perfusion of MC3T3E1 Preosteoblast Spheroids within Polysaccharide-Based Hydrogel Scaffolds: An Experimental and Numerical Study at the Bioreactor Scale. Bioengineering, 10(7), 849. https://doi.org/10.3390/bioengineering10070849