1. Introduction
The importance of water bodies such as reservoirs cannot be overstated. Reservoirs play a critical role in maintaining ecological balance, providing water resources for irrigation, supporting biodiversity, and serving as vital habitats for aquatic life [
1,
2]. In the face of climate change and increasing human pressure, these artificial water bodies are instrumental in water management, supplying water for both agricultural and domestic purposes [
3]. The Chongón Reservoir, located in the Parque Lago National Recreation Area in Guayaquil, Ecuador, is an important artificial ecosystem that has become central to the local environment and the livelihoods of surrounding communities.
Artificial reservoirs like Chongón are particularly vulnerable to ecological disturbances due to their limited water flow and human interventions. One of the most pervasive challenges affecting reservoirs globally is the spread of invasive aquatic species, with water hyacinth (WH) (
Eichhornia crassipes) being among the most problematic [
4,
5]. The rapid proliferation of this invasive plant poses significant challenges, including reducing water quality, disrupting native biodiversity, and increasing water loss through evapotranspiration [
6]. In Chongón Reservoir, the unchecked growth of WH has resulted in a notable impact on the ecological integrity of the system, affecting its capacity to provide essential ecosystem services.
Chongón Reservoir is crucial for irrigation, biodiversity, and recreation but faces ecological challenges from invasive water hyacinth proliferation.
The Chongón Reservoir provides essential ecosystem services, such as water storage, habitat support, and recreation, but these services are threatened by the spread of water hyacinth, increasing evapotranspiration and water losses [
7,
8]. Increased water loss poses a direct threat to water availability for local communities and agricultural activities, which rely on the reservoir as a crucial water source. Therefore, there is a pressing need to understand the dynamics of WH coverage and its effects on evapotranspiration to develop informed management strategies [
9,
10].
WH, originally native to the Amazon Basin, is a highly adaptable plant that has spread across numerous regions worldwide, primarily due to human activities [
11]. This plant is known for its rapid growth and ability to form dense mats that can cover entire water surfaces, effectively blocking sunlight and reducing oxygen levels in the water. This process results in the decline of native aquatic vegetation and a decrease in the diversity of aquatic fauna, ultimately threatening the entire ecosystem [
12]. Understanding and mitigating the impact of WH on reservoirs like Chongón is crucial for preserving the ecological health of these systems.
Remote sensing technology has proven to be an effective tool for monitoring changes in aquatic vegetation over time [
13,
14]. By utilizing the Normalized Difference Vegetation Index (NDVI) derived from satellite imagery, researchers can accurately quantify changes in vegetation coverage, including the proliferation of invasive species such as HW [
15,
16]. This study employs remote sensing data combined with meteorological records to model the spatial and temporal dynamics of WH coverage in the Chongón Reservoir over a 20-year period from 2002 to 2022.
The study focuses on examining how the proliferation of WH influences evapotranspiration rates in the Chongón Reservoir and the resulting implications for water availability and ecosystem health. By applying linear regression models, the relationships between WH coverage, evapotranspiration, and climatic variables are explored. Statistical modeling provides deeper insights into the drivers of evapotranspiration and helps identify key factors contributing to water loss in the reservoir [
17].
The objectives of this research are: (a) to model the relationship between WH coverage and evapotranspiration rates in the Chongón Reservoir, (b) determine the spatial and temporal variability of WH coverage using NDVI, and (c) provide recommendations for the management of invasive species to preserve the ecological integrity of the reservoir. By achieving these objectives, this study aims to contribute valuable knowledge for the effective management of aquatic ecosystems and inform policy decisions that support sustainable water resource management.
3. Results
3.1. Water Hyacinth Coverage in Chongón Reservoir
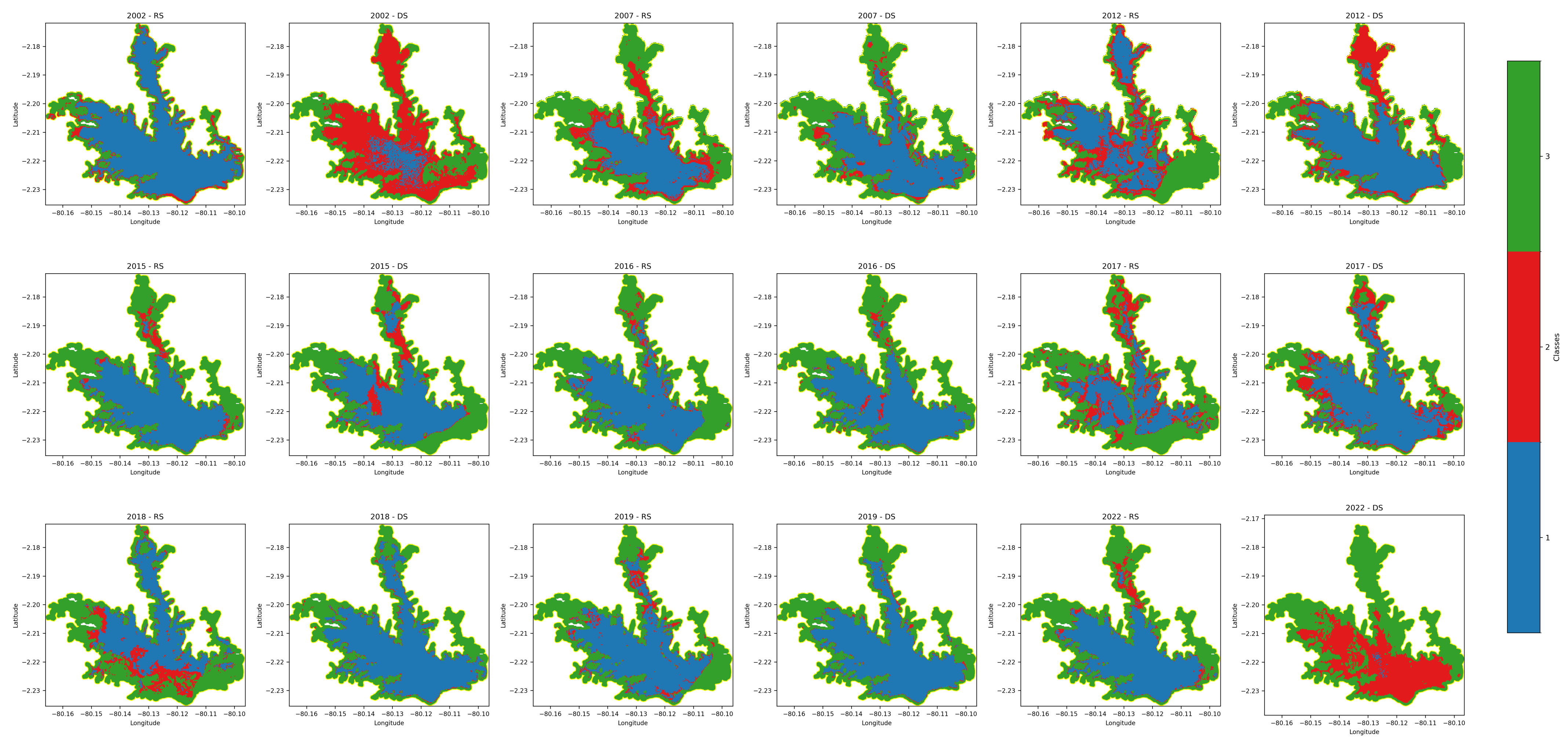
The assessment of water hyacinth coverage (WHC) in the Chongón Reservoir revealed significant spatial and temporal variations over the nine-year study period. Utilizing satellite imagery from Landsat 7, 8, and 9, along with the Normalized Difference Vegetation Index (NDVI), we quantified the extent of WH proliferation during both the rainy season (December to May) and the dry season (June to November).
Our analysis indicated a general trend of increasing WHC over time, with notable fluctuations between seasons. In 2002, during the rainy season, WH covered approximately 1.68 km2, representing 10.42% of the reservoir’s surface area. In contrast, the dry season of the same year showed an increase in coverage to 3.85 km2 (23.87%).
By 2017, the rainy season coverage had expanded significantly to 6.82 km
2, accounting for 42.33% of the reservoir’s area. Similarly, during the dry season of 2019, the coverage reached 4.68 km
2 (29.05%). These findings demonstrate a substantial proliferation over the years.
Figure 3 illustrates the changes in WHC from 2002 to 2022 for both seasons.
Generally, higher coverage was observed during the rainy seasons compared to the dry seasons. The mean coverage during the rainy seasons was 4.99 km2, whereas the dry seasons exhibited a mean coverage of 4.22 km2. However, exceptions were noted in certain years. For instance, in 2002 and 2022, the dry season coverage surpassed that of the rainy season, indicating that factors other than precipitation, such as temperature and water management practices, may influence the proliferation.
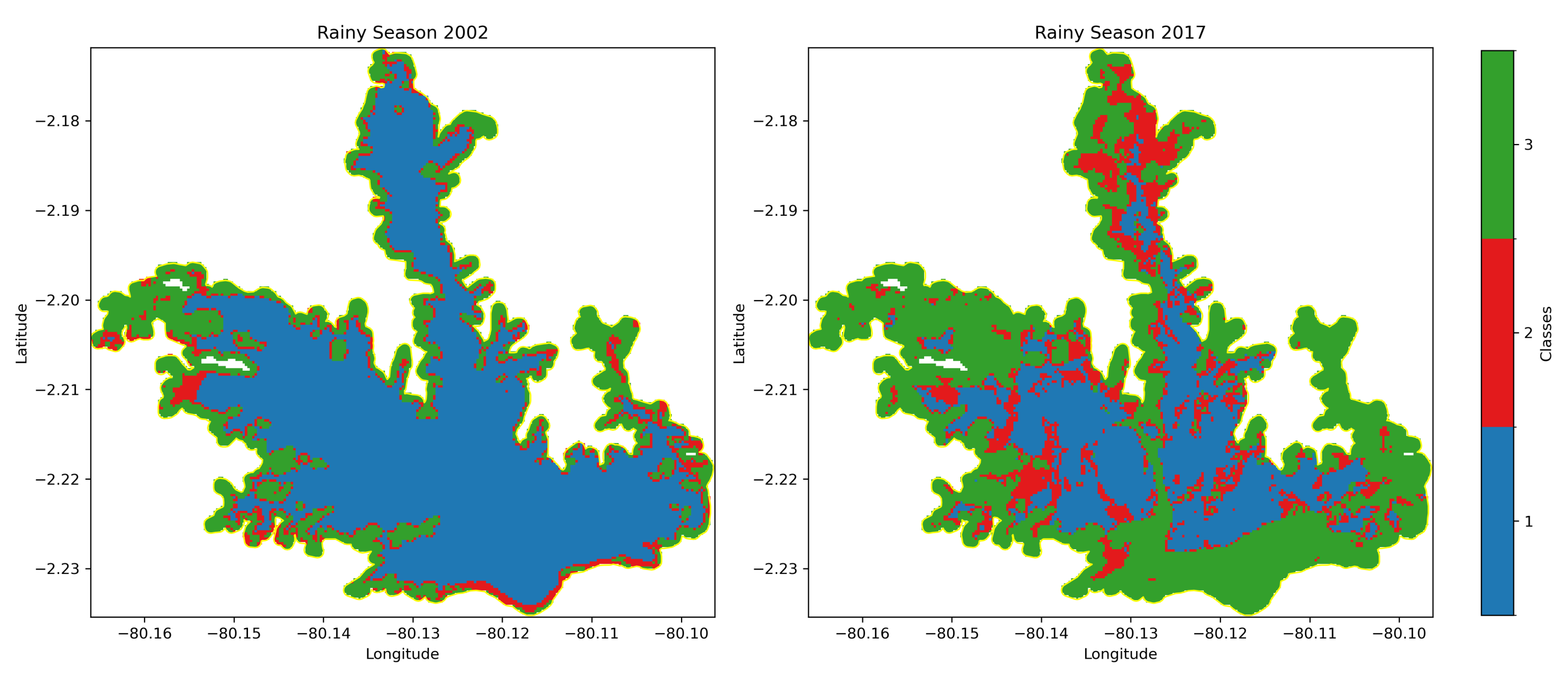
Spatial analysis revealed that WH initially occupied areas along the reservoir’s periphery but gradually extended towards central regions over time. The densest infestations were predominantly located in the northern and northeastern sections of the reservoir.
Figure 4 depict the spatial distribution of WH during the rainy seasons of 2002 and 2017, respectively.
Table 2 summarizes the WHC and corresponding percentages of the reservoir’s surface area for selected years.
The data exhibit variability in WHC between different years and seasons. Notably, the highest coverage during the rainy season was recorded in 2017 (6.82 km2, 42.33%), while the highest dry season coverage occurred in 2015 (5.97 km2, 37.07%). The lowest coverage was observed in 2002 and 2022, indicating possible fluctuations due to environmental factors or management interventions.
3.2. Evapotranspiration Rates Associated with Water Hyacinth
The evapotranspiration rates associated with WH in the Chongón reservoir were calculated to quantify the water loss due to this invasive species during different seasons. This section presents the results of the evapotranspiration analysis, highlighting seasonal, monthly, and yearly variations.
Reference evapotranspiration (
) was calculated using the Penman–Monteith method, based on corrected meteorological data obtained from NASA’s POWER database and validated against observations from the ESPOL-GEA meteorological station. Data correction involved bias adjustment using quantile mapping to ensure accuracy [
35].
Figure 5 presents the Taylor diagrams that illustrate the variation in
on different temporal scales: monthly, seasonal, and yearly. The diagrams help visualize the correlation, standard deviation and centered RMSD between observed and corrected
, providing information on the precision of the bias correction method.
Taylor diagrams provide a visual summary of how well modeled data match observed data by simultaneously illustrating correlation coefficients, standard deviations, and root mean square differences (RMSD). These diagrams allow quick assessment of the accuracy and performance of statistical corrections applied to satellite-derived evapotranspiration data.
Figure 6 shows the box plots of
for both the rainy and dry seasons, before and after bias correction. The results indicate that
is higher during the dry season, reflecting higher temperatures, increased solar radiation, and lower humidity typical of this period. The comparison between the plots before and after correction highlights the impact of the bias adjustment in aligning the model estimates with the observed data.
The specific evapotranspiration was calculated using the crop coefficient method, where:
Here:
is the evapotranspiration of WH (mm/year).
is the reference evapotranspiration (mm/year).
is the crop coefficient for WH.
The values of the crop coefficient were determined from the literature [
36,
37] and adjusted for local conditions:
The calculated evapotranspiration rates showed distinct seasonal variations.
Table 3 summarizes the statistical measures of
for both seasons.
The mean evapotranspiration rate during the rainy season was approximately 2309.90 mm/year, higher than the dry season mean of 1917.87 mm/year. This difference reflects the increased physiological activity of the plants during the rainy season, driven by higher humidity and favorable growth conditions.
The results indicate that the WH evapotranspiration rates are consistently higher during the rainy season across all years studied. The variability is also greater in the rainy season, suggesting that environmental conditions during this period have a significant influence on plant transpiration rates.
The higher evapotranspiration rates during the rainy season can be attributed to:
Increased Biomass: WH tends to grow more rapidly during the rainy season, increasing leaf area and transpiration surface.
Higher Humidity: Elevated humidity levels reduce stomatal closure, allowing for higher transpiration rates.
Optimal Temperatures: Favorable temperatures enhance metabolic processes and water uptake.
In contrast, during the dry season, despite higher , the actual evapotranspiration is lower due to reduced plant growth and possible water stress conditions.
The significant evapotranspiration rates imply substantial water loss from the reservoir, particularly during the rainy season when the plant’s coverage and activity are at their peak. This has implications for water resource management, as the invasive species exacerbates water scarcity issues.
3.3. Net Daily Water Loss Due to Evapotranspiration
The net daily water loss (NDWL) due to evapotranspiration associated with WH was calculated to quantify the additional water loss from the reservoir caused by the presence of WH. The evaporation rate from open water surfaces was calculated using the Penman equation, adjusted for open water conditions. The mean evaporation rate from open water was found to be 1600 mm/year, which translates to approximately 4.38 mm/day.
Table 4 presents the calculated NDWL values for the years studied during both the rainy and dry seasons.
The NDWL values indicate the additional water loss from the reservoir due to the presence of WH, after accounting for the evaporation that would occur from open water surfaces. The highest NDWL was observed during the rainy season of 2017, corresponding to the highest WH coverage recorded.
The analysis shows that NDWL increases with the expansion of WH coverage. The seasonal variation of NDWL reflects both the changes in WH area and the differences in evapotranspiration rates between the rainy and dry seasons. A Pearson correlation analysis between NDWL and WHC confirmed a strong positive correlation (, ), indicating that NDWL is directly related to the extent of WH coverage.
Additionally, the net daily water loss was standardized by water hyacinth coverage (
Table 5), clearly confirming higher standardized evapotranspiration rates during the rainy seasons compared to dry seasons.
3.4. Relationship Between Coverage and Water Loss Due to Evapotranspiration
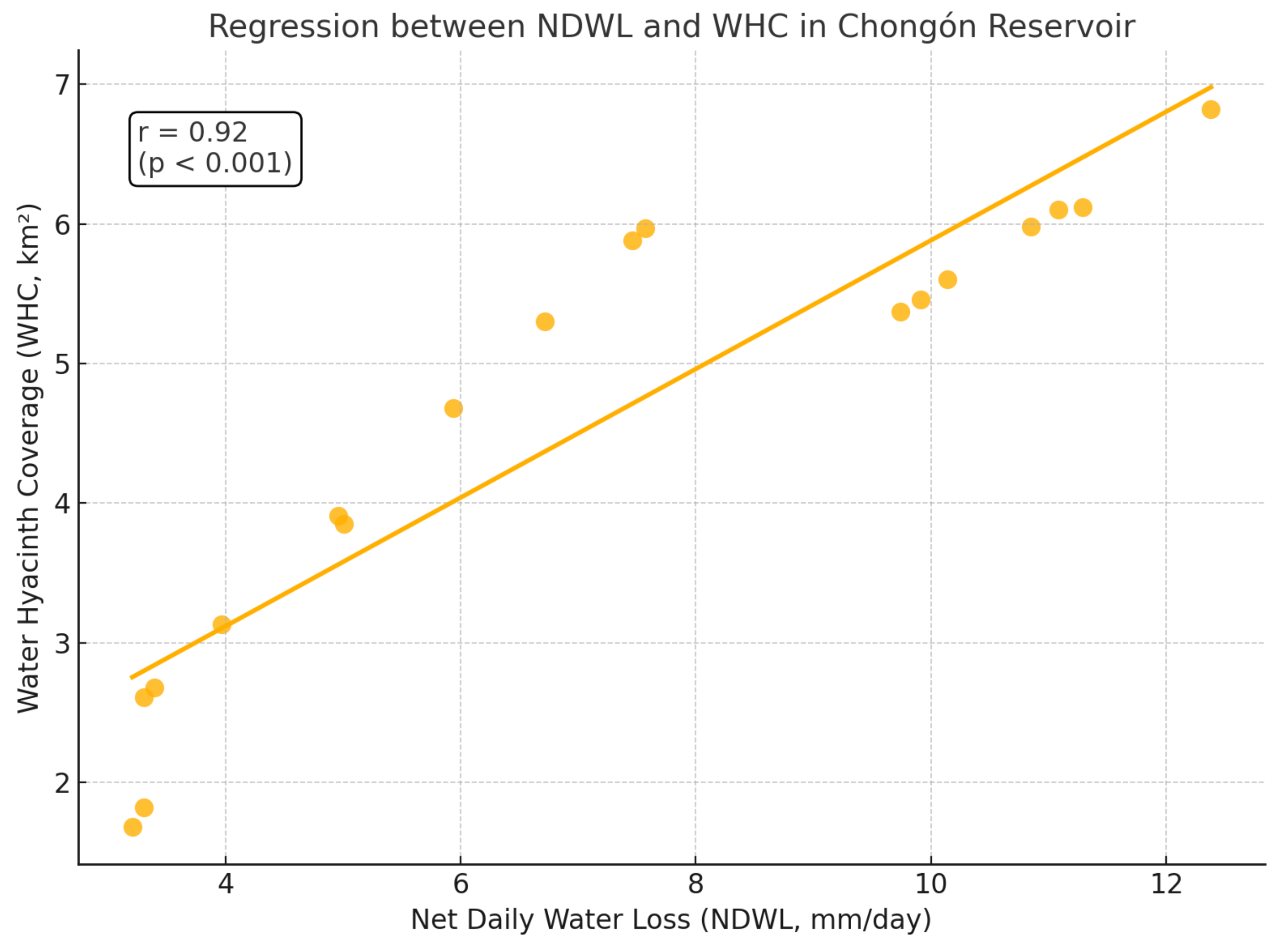
The relationship between the WHC in the Chongón Reservoir and the water loss due to evapotranspiration was analyzed to understand the impact of this invasive species on the reservoir’s water balance. Statistical analyses were performed to assess the correlation between the extent of WHC and the net daily water loss (NDWL) attributed to evapotranspiration.
The correlation between WHC and NDWL was evaluated using Pearson’s correlation coefficient (r). The analysis revealed a strong positive correlation between WHC and NDWL, with a coefficient of . This indicates that as the coverage increases, the water loss due to evapotranspiration also increases significantly.
Figure 7 clearly illustrates this strong linear relationship, visually confirming the direct influence of WH coverage on evapotranspiration-driven water loss.
Table 6 presents the correlation coefficients between WHC and other climatic variables, such as reference evapotranspiration (
), temperature, precipitation, and solar radiation. The strong correlation with NDWL highlights the substantial impact of WHC on water loss from the reservoir.
The negative correlation between WHC and reference evapotranspiration () suggests a weak and negligible relationship, indicating that the WH extent is not directly influenced by the reference evapotranspiration. Similarly, the correlations with temperature and precipitation variables are not statistically significant, implying that other factors may be influencing the plant proliferation.
A linear regression model was developed to quantify the relationship between WHC and NDWL. The model is expressed by the equation:
where:
WHC is the coverage area of WH (km2).
is the intercept of the model.
is the slope coefficient representing the change in coverage per unit change in NDWL.
NDWL is the net daily water loss (mm/year).
The regression analysis yielded the following model parameters:
The model explains approximately 87.59% of the variability in WHC, as indicated by the coefficient of determination (). The slope coefficient is statistically significant (p-value < 0.001), confirming the strong positive relationship between NDWL and WHC.
The analysis indicates that the expansion of WH in the reservoir significantly contributes to increased water loss through evapotranspiration. As the coverage area grows, the cumulative transpiration from the plants leads to higher NDWL values, exacerbating water scarcity issues in the region.
3.5. Model Development
Based on the correlation analysis, NDWL showed a strong positive correlation with WHC (, ). Other climatic variables, such as solar radiation, minimum temperature, and minimum wind speed, were also considered for inclusion in the model due to their potential influence on evapotranspiration processes.
A multiple linear regression analysis was performed using WHC as the dependent variable and NDWL, solar radiation (Rad
s), minimum temperature (Temp
min), and minimum wind speed (WS
min) as independent variables. The regression equation is expressed as:
The estimated coefficients and statistical significance are presented in
Table 7.
The model’s overall fit was evaluated using the coefficient of determination and F-test statistics. The multiple regression model exhibited an value of 0.7085 and an adjusted of 0.6188, indicating that approximately 70.85% of the variability in WHC is explained by the model. The F-statistic was 7.9 with a p-value of 0.001856, suggesting that the model is statistically significant.
Despite the overall significance, individual predictor variables showed varying levels of statistical significance. Solar radiation (Rads) had a p-value of 0.0835, indicating marginal significance, while NDWL, minimum temperature, and minimum wind speed were not statistically significant predictors at the 0.05 level.
Given the strong correlation between WHC and NDWL observed earlier, a simpler linear regression model using only NDWL as the predictor was also developed:
The estimated coefficients for the simple model are provided in
Table 8.
This simple model demonstrated a higher explanatory power, with an of 0.8759 and an adjusted of 0.8681. This indicates that approximately 87.59% of the variability in WHC is explained by NDWL alone. Both the intercept and the NDWL coefficient were statistically significant (p-values < 0.01).
Residual analysis was conducted for both models to assess the adequacy of the fit. The residuals of the simple model were randomly distributed around zero without apparent patterns, suggesting that the assumptions of linear regression were satisfied. In contrast, the residuals of the multiple regression model showed greater dispersion and potential heteroscedasticity, indicating that the model may not fully capture the variability in the data.
Comparing both models, the simple linear regression model with NDWL as the sole predictor was preferred due to its higher predictive power and statistical significance of the coefficients. The inclusion of additional climatic variables did not substantially improve the model’s performance and introduced predictors that were not statistically significant.
Therefore, the final model selected for predicting WHC based on NDWL is:
This model implies that for each additional unit increase in NDWL (mm/day), the WHC increases by approximately km2, holding all else constant.
The development of this model provides a quantitative tool for predicting the impact of WH on evapotranspiration and subsequent water loss in the Chongón Reservoir. By focusing on NDWL, which is directly influenced by the WHC, reservoir management can better anticipate water loss due to evapotranspiration associated with this invasive species.
4. Discussion
The present study aimed to assess the impact of water hyacinth coverage (WHC) on evapotranspiration rates in the Chongón Reservoir over a 20-year period using remote sensing techniques. Our findings revealed significant spatial and temporal variations in WHC, with a strong positive correlation between the extent of coverage and the net daily water loss due to evapotranspiration.
Several environmental conditions significantly influence water hyacinth proliferation in freshwater ecosystems. Factors such as elevated nutrient concentrations, particularly nitrogen and phosphorus, substantially enhance the growth and expansion of water hyacinth [
38,
39]. In the Chongón Reservoir, runoff from adjacent agricultural areas may contribute high nutrient loads, favoring rapid biomass accumulation.
Additionally, seasonal fluctuations in water level and temperature variations directly affect growth dynamics. For instance, elevated water levels during the rainy season typically provide optimal conditions for water hyacinth expansion due to increased habitat availability and reduced physiological stress [
40,
41]. Understanding these contextual environmental factors provides critical insight into the observed temporal variations of water hyacinth coverage, emphasizing the need for integrated nutrient and water-level management strategies in reservoir ecosystems.
Firstly, the WHC in the reservoir showed a substantial increase from 10.42% in 2002 to a peak of 42.33% in 2017 during the rainy seasons. This proliferation corresponds with elevated evapotranspiration rates associated with the plant, which were significantly higher during the rainy season (mean of 2309.90 mm/year) compared to the dry season (mean of 1917.87 mm/year). The strong positive correlation (
,
) between WHC and net daily water loss underscores the substantial impact of this invasive species on the reservoir’s water balance [
42,
43,
44].
It is important to note that the strong correlation observed between evapotranspiration-related variables and water hyacinth coverage might partly result from both datasets originating from satellite-derived sources. While the quantile mapping bias correction approach mitigates some systematic biases, future studies incorporating independent, ground-based evapotranspiration measurements are recommended to validate these results more robustly.
Previous studies have investigated water hyacinth impacts on evapotranspiration in various freshwater ecosystems [
43,
45,
46]. However, the current research distinguishes itself by employing a long-term, high-resolution remote sensing analysis spanning two decades specifically focused on the Chongón Reservoir, an ecologically critical region subject to significant climatic variability due to El Niño events. Additionally, the integration of bias-corrected satellite-derived meteorological data and detailed seasonal comparisons provides a novel and comprehensive perspective on the seasonal dynamics of water hyacinth evapotranspiration impacts. Such an approach allows for improved understanding and predictive capability, enhancing regional water resource management strategies specific to tropical reservoir ecosystems under invasive species pressure.
These results are consistent with previous studies that have documented the aggressive growth of WH and its effects on evapotranspiration and water resources. For instance, Harun et al. [
45] found similar increases in evapotranspiration rates due to WH proliferation in the Nile River, leading to significant water loss [
42,
47]. Additionally, Getahun and Kefale [
48] reported that WH infestation in Lake Tana resulted in increased evapotranspiration, exacerbating water scarcity issues.
The elevated evapotranspiration rates associated with WH can be attributed to the plant’s high transpiration capacity, driven by its large leaf area and rapid growth rate [
47]. During the rainy season, favorable environmental conditions such as higher humidity and optimal temperatures enhance the physiological activity of the plant, leading to increased water uptake and transpiration [
49,
50,
51]. This not only reduces the water volume in the reservoir but also affects the water quality by depleting dissolved oxygen levels, which can harm aquatic life [
52].
The implications of these findings are significant for the management of the Chongón Reservoir. The substantial water loss due to evapotranspiration by WH poses a threat to water availability for municipal and agricultural use, especially during dry periods [
53]. The dense mats of WH also hinder navigation, fishing activities, and can lead to the decline of native biodiversity [
12]. Therefore, controlling the spread of WH is crucial to preserving the reservoir’s ecological integrity and ensuring sustainable water resource management.
Despite the robustness of our findings, this study has certain limitations. The reliance on satellite imagery and remote sensing techniques, while effective for large-scale monitoring, may be subject to errors due to cloud cover, atmospheric interference, and resolution constraints [
54,
55]. Additionally, the crop coefficient values used to estimate evapotranspiration were derived from literature and may not perfectly reflect local conditions. Ground-based measurements of evapotranspiration and WH biomass could improve the accuracy of the estimates.
Future research should focus on integrating more precise field data to validate and refine the remote sensing models [
56]. Investigating the effectiveness of different WH control methods, such as biological control agents or mechanical removal, could provide practical solutions for reservoir management [
57]. Moreover, exploring the potential use of WH biomass for bioenergy production could offer sustainable avenues for mitigating the impact of this invasive species while generating economic benefits [
58].
5. Conclusions
This study assessed the impact of water hyacinth (WH; Eichhornia crassipes) on evapotranspiration rates in the Chongón Reservoir over a 20-year period using remote sensing techniques. The analysis revealed a significant increase in water hyacinth coverage (WHC), from 10.42% in 2002 to a peak of 42.33% in 2017 during the rainy seasons. A strong positive correlation (, ) was found between WHC and net daily water loss due to evapotranspiration, indicating that the proliferation of this invasive species substantially contributes to water loss in the reservoir.
The evapotranspiration rates associated with WH were significantly higher during the rainy season (mean of 2309.90 mm/year) compared to the dry season (mean of 1917.87 mm/year). This elevated evapotranspiration during the rainy season can be attributed to favorable environmental conditions that enhance the physiological activity of the plant, leading to increased water uptake and transpiration. The substantial water loss due to evapotranspiration by WH poses a threat to water availability for municipal and agricultural use, especially during dry periods.
The findings underscore the critical need for effective management strategies to control the spread of WH in the Chongón Reservoir. Implementing control measures such as mechanical removal, biological control agents, or the utilization of WH biomass for bioenergy production could mitigate the impact of this invasive species while providing potential economic benefits. Continuous monitoring using remote sensing techniques is essential for tracking the effectiveness of management interventions and ensuring the sustainability of the reservoir’s water resources.
Practical measures for controlling water hyacinth in Chongón Reservoir could include mechanical removal to immediately reduce coverage, biological control using natural predators (e.g., specialized insects), or integrated management combining both mechanical and biological approaches. Chemical control (herbicides) could also be considered, though its ecological implications must be carefully evaluated. Future management plans should evaluate these options carefully, balancing effectiveness, sustainability, and potential environmental impacts.
In conclusion, the study demonstrates that WH proliferation has a significant impact on evapotranspiration rates and water loss in the Chongón Reservoir. Addressing this issue is crucial for preserving the ecological integrity of the reservoir and securing water resources for the surrounding communities. Future research should focus on refining evapotranspiration estimates through ground-based measurements and exploring innovative control methods to manage WH infestations effectively.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}