
Eupatilin Inhibits Reactive Oxygen Species Generation via Akt/NF-κB/MAPK Signaling Pathways in Particulate Matter-Exposed Human Bronchial Epithelial Cells

 and
and 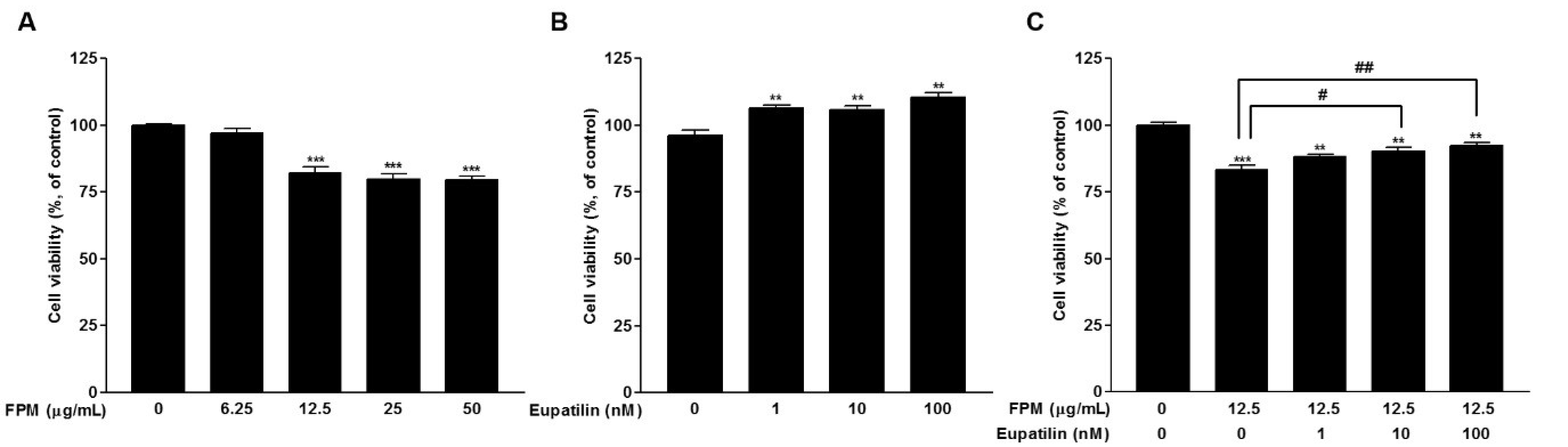{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Cultures
2.3. Cell Viability Assay
2.4. Quantification of Intracellular ROS
2.5. Western Blotting
2.6. Statistical Analyses
3. Result
3.1. Effects of FPM and Eupatilin on the Viability of BEAS-2B Cells
3.2. Opposite Effects of FPM and Eupatilin on ROS Generation in BEAS-2B Cells
3.3. Eupatilin Reduces FPM-Induced ROS Production in BEAS-2B Cells
3.4. FPM Induces Phosphorylation of Akt, p65, and p38 in BEAS-2B Cells
3.5. Eupatilin Reduces Phosphorylation of Akt, p65, and p38 in FPM-Exposed BEAS-2B Cells
3.6. Eupatilin as an Inhibitor of the Akt/NF-κB/MAPK Signaling Pathways in FPM-Exposed BEAS-2B Cells
3.7. Inhibitory Effect of Eupatilin, MK-2206, and BAY11-7082 on ROS Products in BEAS-2B cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Mannucci, P.M.; Harari, S.; Martinelli, I.; Franchini, M. Effects on health of air pollution: A narrative review. Intern. Emerg. Med. 2015, 10, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Yang, C.; Li, J.; Chen, R.; Chen, B.; Gu, D.; Kan, H. Association between long-term exposure to outdoor air pollution and mortality in China: A cohort study. J. Hazard. Mater. 2011, 186, 1594–1600. [Google Scholar] [CrossRef]
- Dockery, D.W.; Pope, C.A.; Xu, X.; Spengler, J.D.; Ware, J.H.; Fay, M.E.; Ferris, B.G.; Speizer, F.E. An Association between Air Pollution and Mortality in Six U.S. Cities. N. Engl. J. Med. 1993, 329, 1753–1759. [Google Scholar] [CrossRef] [PubMed]
- Devries, R.; Kriebel, D.; Sama, S. Outdoor Air Pollution and COPD-Related Emergency Department Visits, Hospital Admissions, and Mortality: A Meta-Analysis. COPD J. Chronic Obstr. Pulm. Dis. 2017, 14, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, H.; Park, Y.H.; Lee, J.T. Assessment of temporal variation for the risk of particulate matters on asthma hospi-talization. Environ. Res. 2017, 156, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Bals, R. Epithelial antimicrobial peptides in host defense against infection. Respir. Res. 2000, 1, 5–50. [Google Scholar] [CrossRef]
- Hiemstra, P. Epithelial antimicrobial peptides and proteins: Their role in host defence and inflammation. Paediatr. Respir. Rev. 2001, 2, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Ru, Q.; Xiong, Q.; Chen, L.; Tian, X.; Yue, K.; Ma, B.; Liu, L.; Wu, R.; Xu, C.; Pi, M.; et al. Lipopolysaccharide accelerates fine particulate matter-induced cell apoptosis in human lung bronchial epithelial cells. Int. J. Occup. Med. Environ. Health 2017, 31, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Li, X.; Nan, A.; Zhang, N.; Chen, L.; Zhou, H.; Zhang, H.; Qiu, M.; Zhu, J.; Ling, Y.; et al. Circular RNA 406961 in-teracts with ILF2 to regulate PM(2.5)-induced inflammatory responses in human bronchial epithelial cells via activation of STAT3/JNK pathways. Environ. Int. 2020, 141, 105755. [Google Scholar] [CrossRef]
- Pfeffer, P.E.; Lu, H.; Mann, E.H.; Chen, Y.-H.; Ho, T.-R.; Cousins, D.J.; Corrigan, C.; Kelly, F.J.; Mudway, I.S.; Hawrylowicz, C.M. Effects of vitamin D on inflammatory and oxidative stress responses of human bronchial epithelial cells exposed to particulate matter. PLoS ONE 2018, 13, e0200040. [Google Scholar] [CrossRef]
- Sotty, J.; Kluza, J.; De Sousa, C.; Tardivel, M.; Anthérieu, S.; Alleman, L.-Y.; Canivet, L.; Perdrix, E.; Loyens, A.; Marchetti, P.; et al. Mitochondrial alterations triggered by repeated exposure to fine (PM2.5-0.18) and quasi-ultrafine (PM0.18) fractions of ambient particulate matter. Environ. Int. 2020, 142, 105830. [Google Scholar] [CrossRef] [PubMed]
- Risom, L.; Møller, P.; Loft, S. Oxidative stress-induced DNA damage by particulate air pollution. Mutat. Res. Mol. Mech. Mutagen. 2005, 592, 119–137. [Google Scholar] [CrossRef]
- Kim, J.-S.; Choi, H.; Oh, J.-M.; Kim, Y.-H.; Kim, S.W.; Kim, S.W.; Kim, B.G.; Cho, J.H.; Lee, J.; Lee, D.C. Effect of fluticasone propionate on human nasal fibroblasts exposed to urban particulate matter. Auris Nasus Larynx 2020, 47, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, K.; Do, D.C.; Zhang, Y.; Hu, X.; Chen, J.; Gao, P. Environmental Exposures and Asthma Development: Autophagy, Mitophagy, and Cellular Senescence. Front. Immunol. 2019, 10, 2787. [Google Scholar] [CrossRef]
- Yu, K.; Li, X.-M.; Xu, X.-L.; Zhang, R.-Y.; Cong, H.-L. Eupatilin protects against tumor necrosis factor-α-mediated inflammation inhuman umbilical vein endothelial cells. Int. J. Clin. Exp. Med. 2015, 8, 22191–22197. [Google Scholar] [PubMed]
- Lee, S.; Lee, M.; Kim, S.-H. Eupatilin inhibits H2O2-induced apoptotic cell death through inhibition of mitogen-activated protein kinases and nuclear factor-kappaB. Food Chem. Toxicol. 2008, 46, 2865–2870. [Google Scholar] [CrossRef]
- Liu, H.; Hao, J.; Wu, C.; Liu, G.; Wang, X.; Yu, J.; Liu, Y.; Zhao, H. Eupatilin Alleviates Lipopolysaccharide-Induced Acute Lung Injury by Inhibiting Inflammation and Oxidative Stress. Med. Sci. Monit. 2019, 25, 8289–8296. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.-J.; Oh, H.-M.; Na, B.-R.; Ramesh, T.P.; Lee, H.-J.; Choi, C.-S.; Choi, S.-C.; Oh, T.-Y.; Chae, J.-R.; Kim, S.-W.; et al. Eupatilin Protects Gastric Epithelial Cells from Oxidative Damage and Down-Regulates Genes Responsible for the Cellular Oxidative Stress. Pharm. Res. 2008, 25, 1355–1364. [Google Scholar] [CrossRef]
- Dominici, F.; Peng, R.D.; Bell, M.L.; Pham, L.; McDermott, A.; Zeger, S.L.; Samet, J.M. Fine Particulate Air Pollution and Hospital Admission for Cardiovascular and Respiratory Diseases. JAMA 2006, 295, 1127–1134. [Google Scholar] [CrossRef]
- Zeng, Q.; Wu, Z.; Jiang, G.; Wu, X.; Li, P.; Ni, Y.; Xiong, X.; Wang, X.; Li, G.; Parasat, X. The association between ambient inhalable particulate matter and the disease burden of respiratory disease: An ecological study based on ten-year time series data in Tianjin, China. Environ. Res. 2017, 157, 71–77. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Peel, J.L.; Hannigan, M.P.; Dutton, S.J.; Sheppard, L.; Clark, M.L.; Vedal, S. The Temporal Lag Structure of Short-term Associations of Fine Particulate Matter Chemical Constituents and Cardiovascular and Respiratory Hospitalizations. Environ. Health Perspect. 2012, 120, 1094–1099. [Google Scholar] [CrossRef] [PubMed]
- Hoek, G.; Krishnan, R.M.; Beelen, R.; Peters, A.; Ostro, B.; Brunekreef, B.; Kaufman, J.D. Long-term air pollution exposure and cardio- respiratory mortality: A review. Environ. Health 2013, 12, 43. [Google Scholar] [CrossRef]
- Long, F.; Jiang, H.; Yi, H.; Su, L.; Sun, J. Particulate matter 2.5 induced bronchial epithelial cell injury via activation of 5′-adenosine monophosphate-activated protein kinase-mediated autophagy. J. Cell Biochem. 2019, 120, 3294–3305. [Google Scholar] [CrossRef]
- Ma, Q.-Y.; Huang, D.-Y.; Zhang, H.-J.; Wang, S.; Chen, X.-F. Exposure to particulate matter 2.5 (PM2.5) induced macrophage-dependent inflammation, characterized by increased Th1/Th17 cytokine secretion and cytotoxicity. Int. Immunopharmacol. 2017, 50, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Abbas, I.; Badran, G.; Verdin, A.; LeDoux, F.; Roumie, M.; Guidice, J.-M.L.; Courcot, D.; Garçon, G. In vitro evaluation of organic extractable matter from ambient PM2.5 using human bronchial epithelial BEAS-2B cells: Cytotoxicity, oxidative stress, pro-inflammatory response, genotoxicity, and cell cycle deregulation. Environ. Res. 2019, 171, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gonzalez, F.J.; Chandel, N.S.; Jain, M.; Budinger, G.S. Reactive oxygen species as signaling molecules in the development of lung fibrosis. Transl. Res. 2017, 190, 61–68. [Google Scholar] [CrossRef]
- Boukhenouna, S.; Wilson, M.A.; Bahmed, K.; Kosmider, B. Reactive Oxygen Species in Chronic Obstructive Pulmonary Disease. Oxidative Med. Cell. Longev. 2018, 2018, 1–9. [Google Scholar] [CrossRef]
- Kim, J.-S.; Oh, J.-M.; Choi, H.; Kim, S.W.; Kim, S.W.; Kim, B.G.; Cho, J.H.; Lee, J.; Lee, D.C. Activation of the Nrf2/HO-1 pathway by curcumin inhibits oxidative stress in human nasal fibroblasts exposed to urban particulate matter. BMC Complement. Med. Ther. 2020, 20, 101–108. [Google Scholar] [CrossRef]
- Du, L.; Chen, J.; Xing, Y.-Q. Eupatilin prevents H2O2-induced oxidative stress and apoptosis in human retinal pigment epithelial cells. Biomed. Pharmacother. 2017, 85, 136–140. [Google Scholar] [CrossRef]
- Watterson, T.L.; Hamilton, B.; Martin, R.S.; Coulombe, R.A. Urban particulate matter activates Akt in human lung cells. Arch. Toxicol. 2011, 86, 121–135. [Google Scholar] [CrossRef]
- Lee, D.C.; Choi, H.; Oh, J.-M.; Lee, J.; Lee, J.; Lee, H.Y.; Kang, J.Y. Urban particulate matter regulates tight junction proteins by inducing oxidative stress via the Akt signal pathway in human nasal epithelial cells. Toxicol. Lett. 2020, 333, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.W.; Lee, T.L.; Chen, Y.C.; Liang, C.J.; Wang, S.H.; Lue, J.H.; Tsai, J.S.; Lee, S.W.; Chen, S.H.; Yang, Y.F.; et al. PM(2.5)-induced oxidative stress increases intercellular adhesion molecule-1 expression in lung epithelial cells through the IL-6/AKT/STAT3/NF-κB-dependent pathway. Part. Fibre Toxicol. 2018, 15, 4. [Google Scholar] [CrossRef]
- Madrid, L.V.; Mayo, M.W.; Reuther, J.Y.; Baldwin, A.S., Jr. Akt stimulates the transactivation potential of the RelA/p65 Subunit of NF-kappa B through utilization of the Ikappa B kinase and activation of the mitogen-activated protein kinase p38. J. Biol. Chem. 2001, 276, 18934–18940. [Google Scholar] [CrossRef] [PubMed]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [PubMed]
- Perkins, N.D. Integrating cell-signalling pathways with NF-kappaB and IKK function. Nat. Rev. Mol. Cell Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef]
- Song, L.; Li, D.; Li, X.; Ma, L.; Bai, X.; Wen, Z.; Zhang, X.; Chen, D.; Peng, L. Exposure to PM2.5 induces aberrant activation of NF-κB in human airway epithelial cells by downregulating miR-331 expression. Environ. Toxicol. Pharmacol. 2017, 50, 192–199. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, W.J.; Xiong, W.; Lu, W.H.; Zheng, H.Y.; Zhou, X.; Yuan, J. PM(2.5) stimulated the release of cytokines from BEAS-2B cells through activation of IKK/NF-κB pathway. Hum. Exp. Toxicol. 2019, 38, 311–320. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; Avruch, J. Mammalian MAPK Signal Transduction Pathways Activated by Stress and Inflammation: A 10-Year Update. Physiol. Rev. 2012, 92, 689–737. [Google Scholar] [CrossRef] [PubMed]
- Ichijo, H.; Nishida, E.; Irie, K.; ten Dijke, P.; Saitoh, M.; Moriguchi, T.; Takagi, M.; Matsumoto, K.; Miyazono, K.; Gotoh, Y. Induction of apoptosis by ASK1, a mammalian MAPKKK that activates SAPK/JNK and p38 signaling pathways. Science 1997, 275, 90–94. [Google Scholar] [CrossRef]
- Lee, J.; Rhee, M.H.; Kim, E.; Cho, J.Y. BAY 11-7082 Is a Broad-Spectrum Inhibitor with Anti-Inflammatory Activity against Multiple Targets. Mediat. Inflamm. 2012, 2012, 1–11. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.C.; Oh, J.-M.; Choi, H.; Kim, S.W.; Kim, S.W.; Kim, B.G.; Cho, J.H.; Lee, J.; Kim, J.-S. Eupatilin Inhibits Reactive Oxygen Species Generation via Akt/NF-κB/MAPK Signaling Pathways in Particulate Matter-Exposed Human Bronchial Epithelial Cells. Toxics 2021, 9, 38. https://doi.org/10.3390/toxics9020038
Lee DC, Oh J-M, Choi H, Kim SW, Kim SW, Kim BG, Cho JH, Lee J, Kim J-S. Eupatilin Inhibits Reactive Oxygen Species Generation via Akt/NF-κB/MAPK Signaling Pathways in Particulate Matter-Exposed Human Bronchial Epithelial Cells. Toxics. 2021; 9(2):38. https://doi.org/10.3390/toxics9020038
Chicago/Turabian StyleLee, Dong Chang, Jeong-Min Oh, Hyunsu Choi, Sung Won Kim, Soo Whan Kim, Byung Guk Kim, Jin Hee Cho, Joohyung Lee, and Ji-Sun Kim. 2021. "Eupatilin Inhibits Reactive Oxygen Species Generation via Akt/NF-κB/MAPK Signaling Pathways in Particulate Matter-Exposed Human Bronchial Epithelial Cells" Toxics 9, no. 2: 38. https://doi.org/10.3390/toxics9020038
APA StyleLee, D. C., Oh, J.-M., Choi, H., Kim, S. W., Kim, S. W., Kim, B. G., Cho, J. H., Lee, J., & Kim, J.-S. (2021). Eupatilin Inhibits Reactive Oxygen Species Generation via Akt/NF-κB/MAPK Signaling Pathways in Particulate Matter-Exposed Human Bronchial Epithelial Cells. Toxics, 9(2), 38. https://doi.org/10.3390/toxics9020038