
Modulation by Ozone of Glucocorticoid-Regulating Factors in the Lungs in Relation to Stress Axis Reactivity

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Experimental Model
2.2. Biological Samples
2.3. Lavage Corticosteroid-Binding Globulin and Soluble Intracellular Adhesion Molecule-1 (ICAM-1)
2.4. Total Elastase Activity
2.5. RNA Extraction
2.6. Gene Expression Analyses
2.7. Statistical Analyses
3. Results
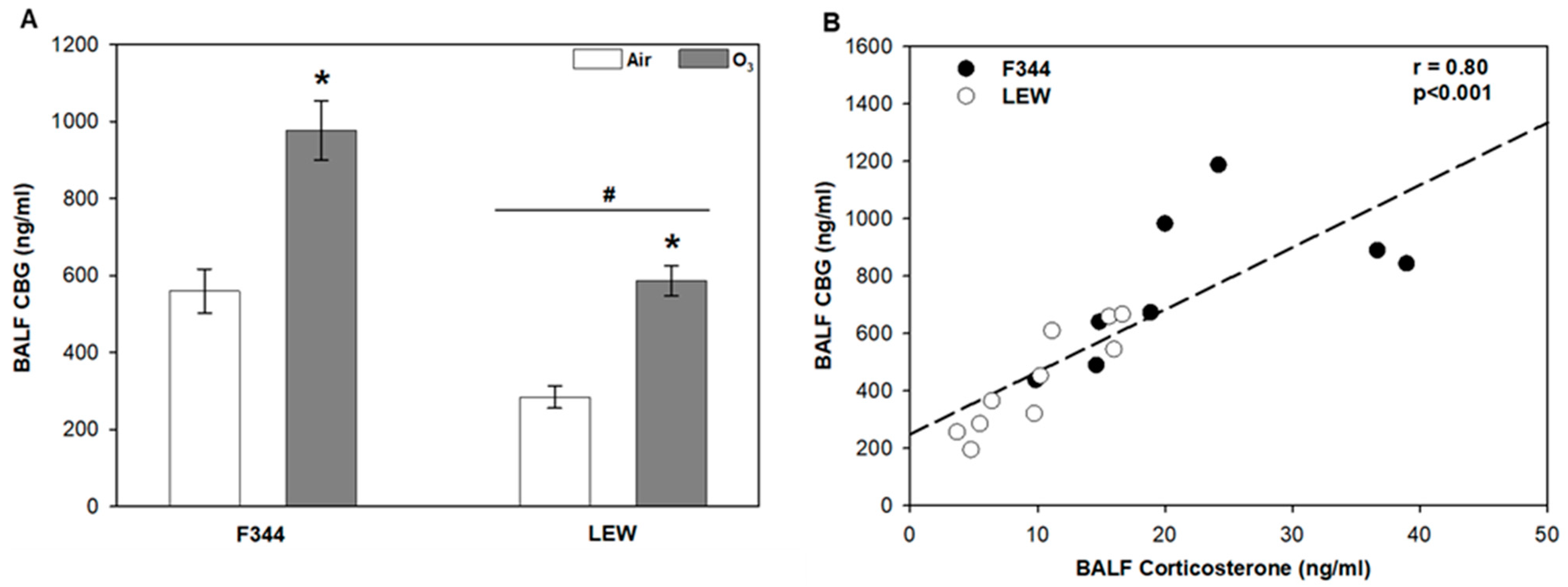
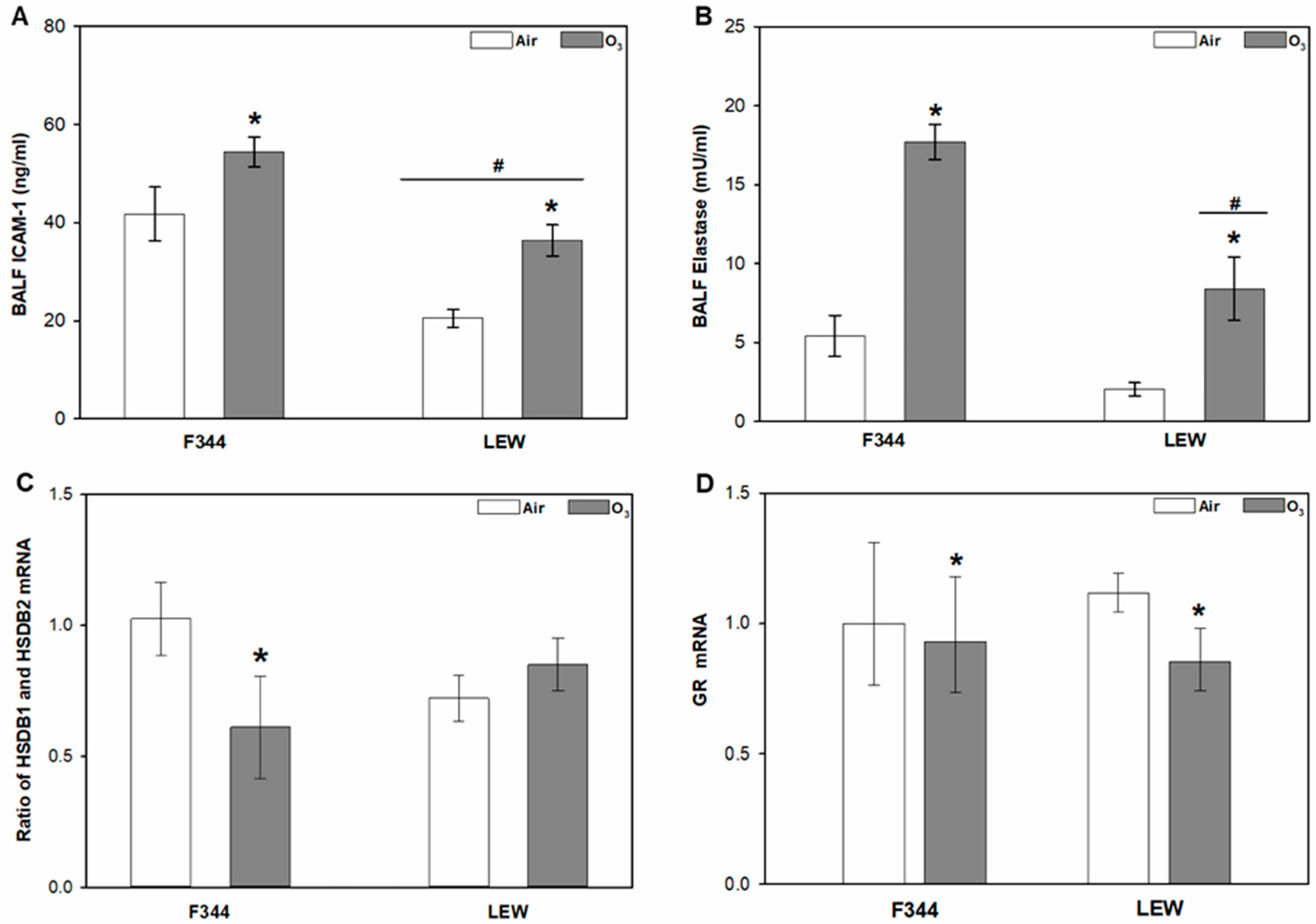
3.1. Effects of O3 on Glucocorticoid-Regulating Factors in the Lungs
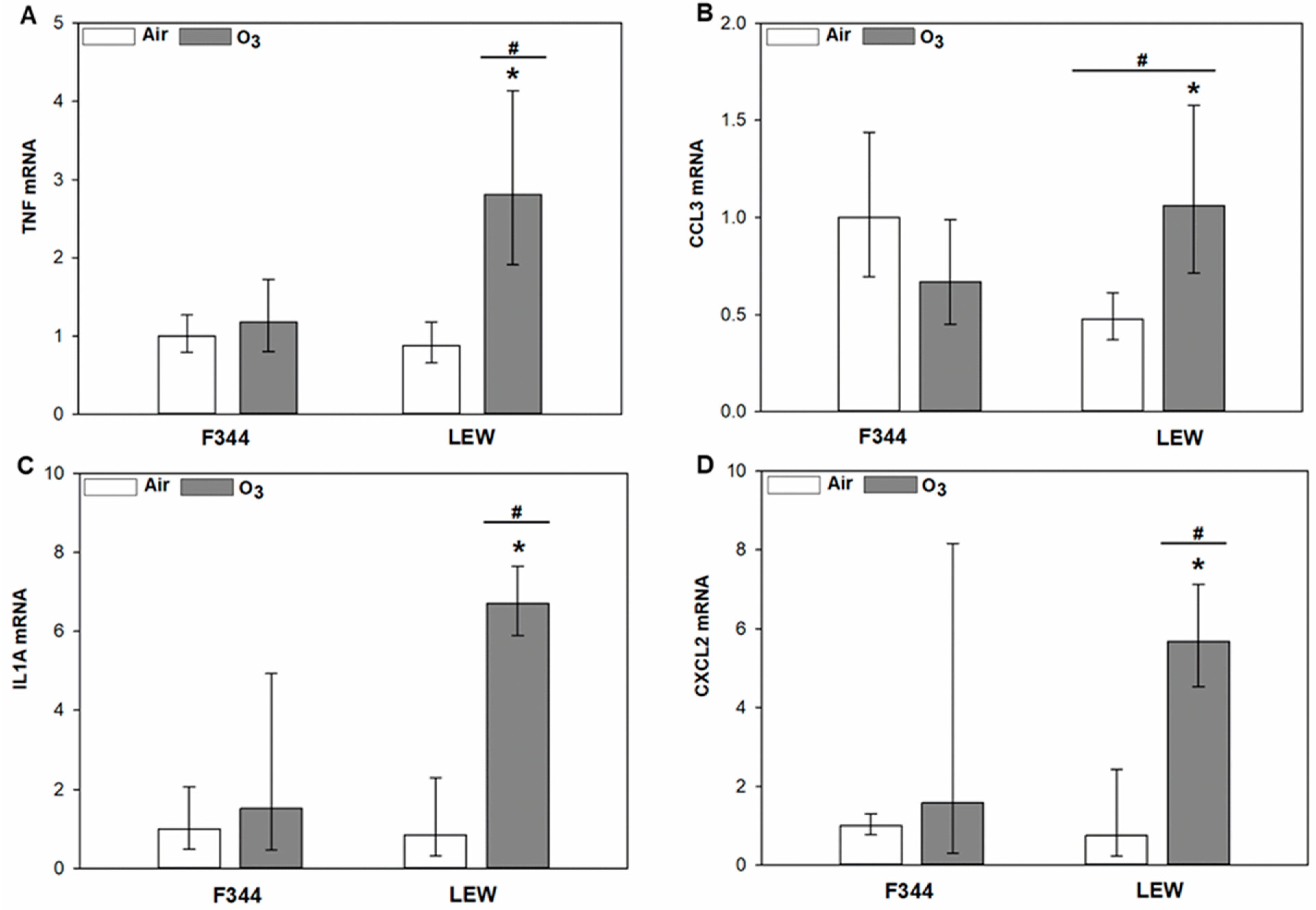
3.2. O3-Dependent Inflammatory Signaling in the Lungs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Katsu, Y.; Iguchi, T. Subchapter 95A—Corticosterone. In Handbook of Hormones; Takei, Y., Ando, H., Tsutsui, K., Eds.; Academic Press: San Diego, CA, USA, 2016. [Google Scholar] [CrossRef]
- Uchoa, E.T.; Aguilera, G.; Herman, J.P.; Fiedler, J.L.; Deak, T.; Cordeiro de Sousa, M.B. Novel Aspects of Hypothalamic-Pituitary-Adrenal Axis Regulation and Glucocorticoid Actions. J. Neuroendocrinol. 2014, 26, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Chrousos, G.P. The Hypothalamic–Pituitary–Adrenal Axis and Immune-Mediated Inflammation. N. Engl. J. Med. 1995, 332, 1351–1363. [Google Scholar] [CrossRef] [PubMed]
- Silverman, M.N.; Sternberg, E.M. Glucocorticoid Regulation of Inflammation and Its Functional Correlates: From HPA Axis to Glucocorticoid Receptor Dysfunction. Ann. N. Y. Acad. Sci. 2012, 1261, 55–63. [Google Scholar] [CrossRef]
- Li, H.; Cai, J.; Chen, R.; Zhao, Z.; Ying, Z.; Wang, L.; Chen, J.; Hao, K.; Kinney, P.L.; Chen, H.; et al. Particulate Matter Exposure and Stress Hormone Levels: A Randomized, Double-Blind, Crossover Trial of Air Purification. Circulation 2017, 136, 618–627. [Google Scholar] [CrossRef]
- Miller, D.B.; Ghio, A.J.; Karoly, E.D.; Bell, L.N.; Snow, S.J.; Madden, M.C.; Soukup, J.; Cascio, W.E.; Gilmour, M.I.; Kodavanti, U.P. Ozone Exposure Increases Circulating Stress Hormones and Lipid Metabolites in Humans. Am. J. Respir. Crit. Care Med. 2016, 193, 1382–1391. [Google Scholar] [CrossRef] [PubMed]
- Thomson, E.M.; Filiatreault, A.; Williams, A.; Rider, C.F.; Carlsten, C. Exposure to Diesel Exhaust and Plasma Cortisol Response: A Randomized Double-Blind Crossover Study. Environ. Health Perspect. 2021, 129, 037701. [Google Scholar] [CrossRef] [PubMed]
- Thomson, E.M.; Vladisavljevic, D.; Mohottalage, S.; Kumarathasan, P.; Vincent, R. Mapping Acute Systemic Effects of Inhaled Particulate Matter and Ozone: Multiorgan Gene Expression and Glucocorticoid Activity. Toxicol. Sci. 2013, 135, 169–181. [Google Scholar] [CrossRef]
- Thomson, E.M.; Pal, S.; Guénette, J.; Wade, M.G.; Atlas, E.; Holloway, A.C.; Williams, A.; Vincent, R. Ozone Inhalation Provokes Glucocorticoid-Dependent and -Independent Effects on Inflammatory and Metabolic Pathways. Toxicol. Sci. 2016, 152, 17–28. [Google Scholar] [CrossRef]
- Thomas, J.; Guénette, J.; Thomson, E.M. Stress Axis Variability Is Associated with Differential Ozone-Induced Lung Inflammatory Signaling and Injury Biomarker Response. Environ. Res. 2018, 167, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Kadmiel, M.; Cidlowski, J.A. Glucocorticoid Receptor Signaling in Health and Disease. Trends Pharmacol. Sci. 2013, 34, 518–530. [Google Scholar] [CrossRef] [PubMed]
- Moisan, M.-P.; Castanon, N. Emerging Role of Corticosteroid-Binding Globulin in Glucocorticoid-Driven Metabolic Disorders. Front. Endocrinol. 2016, 7, 160. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pemberton, P.A.; Stein, P.E.; Pepys, M.B.; Potter, J.M.; Carrell, R.W. Hormone Binding Globulins Undergo Serpin Conformational Change in Inflammation. Nature 1988, 336, 257–258. [Google Scholar] [CrossRef] [PubMed]
- Chapman, K.; Holmes, M.; Seckl, J. 11β-Hydroxysteroid Dehydrogenases: Intracellular Gate-Keepers of Tissue Glucocorticoid Action. Physiol. Rev. 2013, 93, 1139–1206. [Google Scholar] [CrossRef] [PubMed]
- Watterberg, K.L.; Scott, S.M.; Backstrom, C.; Gifford, K.L.; Cook, K.L. Links between Early Adrenal Function and Respiratory Outcome in Preterm Infants: Airway Inflammation and Patent Ductus Arteriosus. Pediatrics 2000, 105, 320–324. [Google Scholar] [CrossRef]
- Zhang, S.; Shen, Z.; Hu, G.; Liu, R.; Zhang, X. Effects of Endogenous Glucocorticoids on Allergic Inflammation and TH1/TH2 Balance in Airway Allergic Disease. Ann. Allergy Asthma Immunol. 2009, 103, 525–534. [Google Scholar] [CrossRef]
- Cohen, H.; Zohar, J.; Gidron, Y.; Matar, M.A.; Belkind, D.; Loewenthal, U.; Kozlovsky, N.; Kaplan, Z. Blunted HPA Axis Response to Stress Influences Susceptibility to Posttraumatic Stress Response in Rats. Biol. Psychiatry 2006, 59, 1208–1218. [Google Scholar] [CrossRef] [PubMed]
- Dhabhar, F.S.; McEwen, B.S.; Spencer, R.L. Stress Response, Adrenal Steroid Receptor Levels and Corticosteroid-Binding Globulin Levels - a Comparison between Sprague-Dawley, Fischer 344 and Lewis Rats. Brain Res. 1993, 616, 89–98. [Google Scholar] [CrossRef]
- Karalis, K.; Crofford, L.; Wilder, R.L.; Chrousos, G.P. Glucocorticoid and/or Glucocorticoid Antagonist Effects in Inflammatory Disease-Susceptible Lewis Rats and Inflammatory Disease-Resistant Fischer Rats. Endocrinology 1995, 136, 3107–3112. [Google Scholar] [CrossRef] [PubMed]
- Sternberg, E.M.; Hill, J.M.; Chrousos, G.P.; Kamilaris, T.; Listwak, S.J.; Gold, P.W.; Wilder, R.L. Inflammatory Mediator-Induced Hypothalamic-Pituitary-Adrenal Axis Activation Is Defective in Streptococcal Cell Wall Arthritis-Susceptible Lewis Rats. Proc. Natl. Acad. Sci. USA 1989, 86, 2374–2378. [Google Scholar] [CrossRef] [PubMed]
- Charmandari, E.; Tsigos, C.; Chrousos, G. Endocrinology of the Stress Response. Annu. Rev. Physiol. 2005, 67, 259–284. [Google Scholar] [CrossRef]
- Ebner, K.; Singewald, N. Individual Differences in Stress Susceptibility and Stress Inhibitory Mechanisms. Curr. Opin. Behav. Sci. 2017, 14, 54–64. [Google Scholar] [CrossRef]
- Elenkov, I.J.; Kvetnansky, R.; Hashiramoto, A.; Bakalov, V.K.; Link, A.A.; Zachman, K.; Crane, M.; Jezova, D.; Rovensky, J.; Dimitrov, M.A.; et al. Low- versus High-Baseline Epinephrine Output Shapes Opposite Innate Cytokine Profiles: Presence of Lewis- and Fischer-like Neurohormonal Immune Phenotypes in Humans? J. Immunol. 2008, 181, 1737–1745. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative Pcr and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Thomas, J.; Stalker, A.; Breznan, D.; Thomson, E.M. Ozone-Dependent Increases in Lung Glucocorticoids and Macrophage Response: Effect Modification by Innate Stress Axis Function. Environ. Toxicol. Pharmacol. 2021, 86, 103662. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.W.; Walker, B.R.; Shaw, R.J.; Honour, J.W.; Jessop, D.S.; Lightman, S.L.; Zumla, A.; Rook, G.a.W. Increased Cortisol: Cortisone Ratio in Acute Pulmonary Tuberculosis. Am. J. Respir. Crit. Care. Med. 2000, 162, 1641–1647. [Google Scholar] [CrossRef]
- Corvol, H.; Nathan, N.; Charlier, C.; Chadelat, K.; Le Rouzic, P.; Tabary, O.; Fauroux, B.; Henrion-Caude, A.; Feingold, J.; Boelle, P.-Y.; et al. Glucocorticoid Receptor Gene Polymorphisms Associated with Progression of Lung Disease in Young Patients with Cystic Fibrosis. Respir. Res. 2007, 8, 88. [Google Scholar] [CrossRef]
- Hammond, G.L. Plasma Steroid-Binding Proteins: Primary Gatekeepers of Steroid Hormone Action. J. Endocrinol. 2016, 230, R13–R25. [Google Scholar] [CrossRef] [PubMed]
- Aman, J.; van der Heijden, M.; van Lingen, A.; Girbes, A.R.J.; van Nieuw Amerongen, G.P.; van Hinsbergh, V.W.M.; Groeneveld, A.B.J. Plasma Protein Levels Are Markers of Pulmonary Vascular Permeability and Degree of Lung Injury in Critically Ill Patients with or at Risk for Acute Lung Injury/Acute Respiratory Distress Syndrome. Crit. Care Med. 2011, 39, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Gulfo, J.; Ledda, A.; Gea-Sorlí, S.; Bonjoch, L.; Closa, D.; Grasa, M.; Esteve, M. New Roles for Corticosteroid Binding Globulin and Opposite Expression Profiles in Lung and Liver. PLoS ONE 2016, 11, e0146497. [Google Scholar] [CrossRef]
- Spadaro, S.; Fogagnolo, A.; Campo, G.; Zucchetti, O.; Verri, M.; Ottaviani, I.; Tunstall, T.; Grasso, S.; Scaramuzzo, V.; Murgolo, F.; et al. Markers of Endothelial and Epithelial Pulmonary Injury in Mechanically Ventilated COVID-19 ICU Patients. Crit. Care 2021, 25, 74. [Google Scholar] [CrossRef]
- Nénan, S.; Boichot, E.; Lagente, V.; Bertrand, C.P. Macrophage Elastase (MMP-12): A pro-Inflammatory Mediator? Mem. Inst. Oswaldo Cruz 2005, 100, 167–172. [Google Scholar] [CrossRef]
- Elkington, P.T.G.; Friedland, J.S. Matrix Metalloproteinases in Destructive Pulmonary Pathology. Thorax 2006, 61, 259–266. [Google Scholar] [CrossRef]
- Soldan, M.; Nagel, G.; Losekam, M.; Ernst, M.; Maser, E. Interindividual Variability in the Expression and NNK Carbonyl Reductase Activity of 11β-Hydroxysteroid Dehydrogenase 1 in Human Lung. Cancer Lett. 1999, 145, 49–56. [Google Scholar] [CrossRef]
- Rider, C.F.; Carlsten, C. Air Pollution and Resistance to Inhaled Glucocorticoids: Evidence, Mechanisms and Gaps to Fill. Pharmacol. Ther. 2019, 194, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.-Q.; Xie, W.-Y.; Tang, Y.-J.; Zhang, J.; Liu, J. Genetic Variation in the Glucocorticoid Pathway Involved in Interindividual Differences in the Glucocorticoid Treatment. Pharmacogenomics 2017, 18, 293–316. [Google Scholar] [CrossRef]
- Bekhbat, M.; Rowson, S.A.; Neigh, G.N. Checks and Balances: The Glucocorticoid Receptor and NFκB in Good Times and Bad. Front. Neuroendocrinol. 2017, 46, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Cain, D.W.; Cidlowski, J.A. Immune Regulation by Glucocorticoids. Nat. Rev. Immunol. 2017, 17, 233–247. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, A.E.; Chapman, K.E. The Anti-Inflammatory and Immunosuppressive Effects of Glucocorticoids, Recent Developments and Mechanistic Insights. Mol. Cell. Endocrinol. 2011, 335, 2–13. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomas, J.; Thomson, E.M. Modulation by Ozone of Glucocorticoid-Regulating Factors in the Lungs in Relation to Stress Axis Reactivity. Toxics 2021, 9, 290. https://doi.org/10.3390/toxics9110290
Thomas J, Thomson EM. Modulation by Ozone of Glucocorticoid-Regulating Factors in the Lungs in Relation to Stress Axis Reactivity. Toxics. 2021; 9(11):290. https://doi.org/10.3390/toxics9110290
Chicago/Turabian StyleThomas, Jith, and Errol M. Thomson. 2021. "Modulation by Ozone of Glucocorticoid-Regulating Factors in the Lungs in Relation to Stress Axis Reactivity" Toxics 9, no. 11: 290. https://doi.org/10.3390/toxics9110290
APA StyleThomas, J., & Thomson, E. M. (2021). Modulation by Ozone of Glucocorticoid-Regulating Factors in the Lungs in Relation to Stress Axis Reactivity. Toxics, 9(11), 290. https://doi.org/10.3390/toxics9110290