
Network Toxicology and Molecular Docking to Elucidate the Mechanisms of Intestinal Toxicity Induced by P-Phenylenediamine Antioxidants and Their Quinone Derivatives


Abstract
1. Introduction
2. Methods
2.1. Target Construction of PPDs and PPDQs
2.2. Target Construction of Intestinal Toxicity
2.3. Acquisition of Intersection Targets
2.4. Constructing of PPI Network
2.5. Constructing of Drug–Target–Pathway Network
2.6. GO and KEGG Pathway Analysis
2.7. Molecular Docking
3. Results
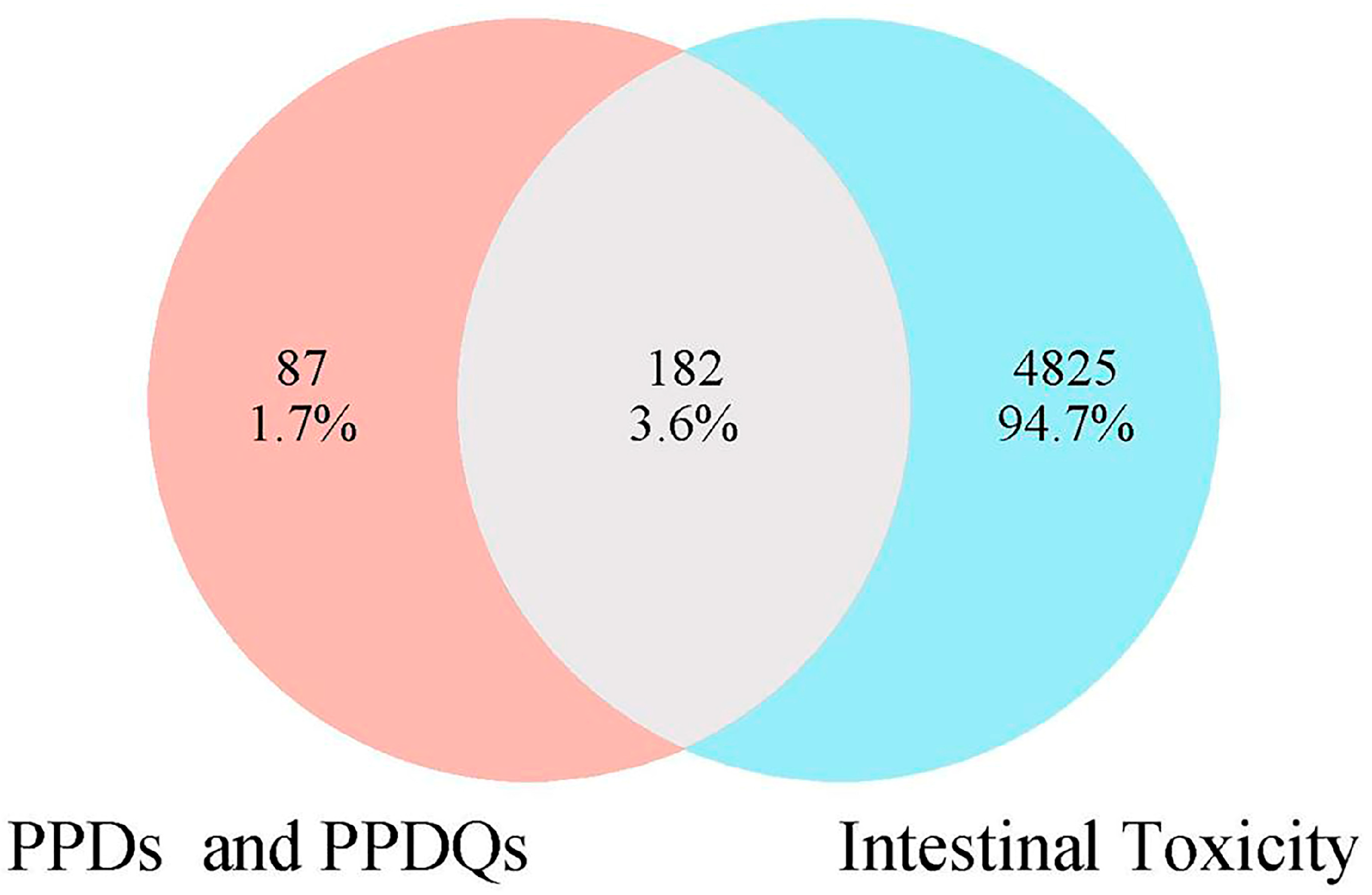
3.1. Potential Targets of PPDs- and PPDQs-Induced Intestinal Toxicity
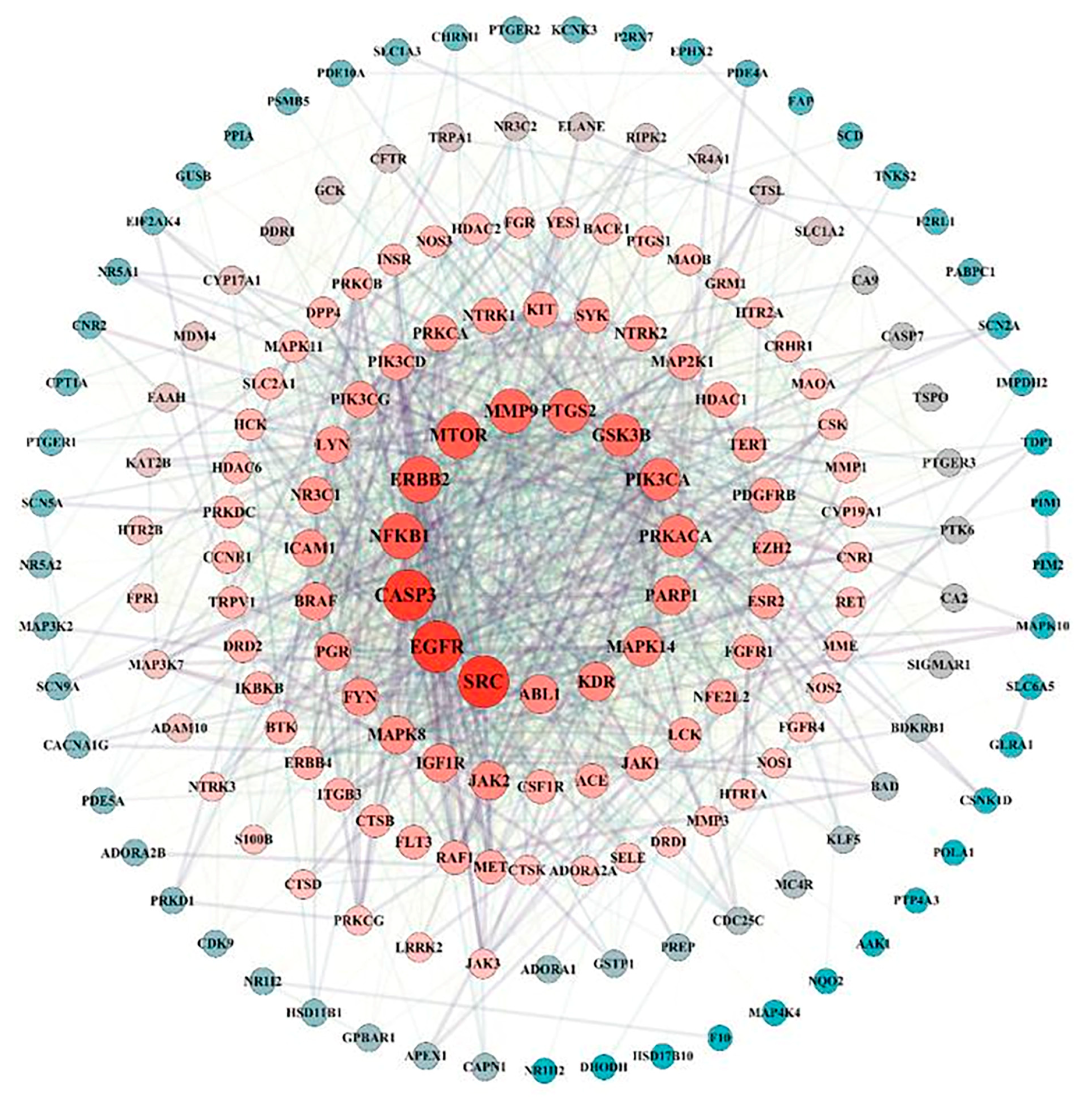
3.2. PPI Network and Key Target Acquisition
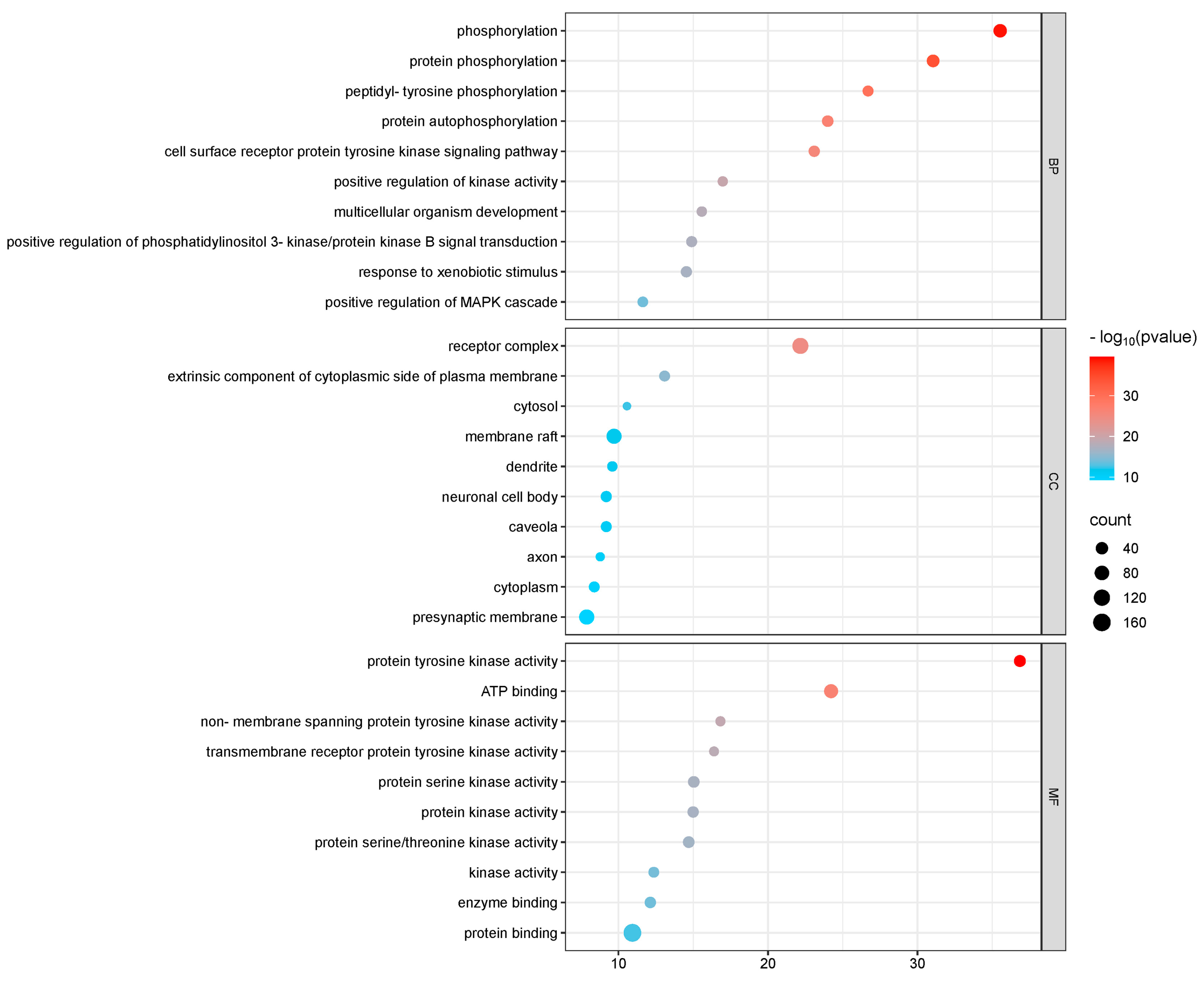
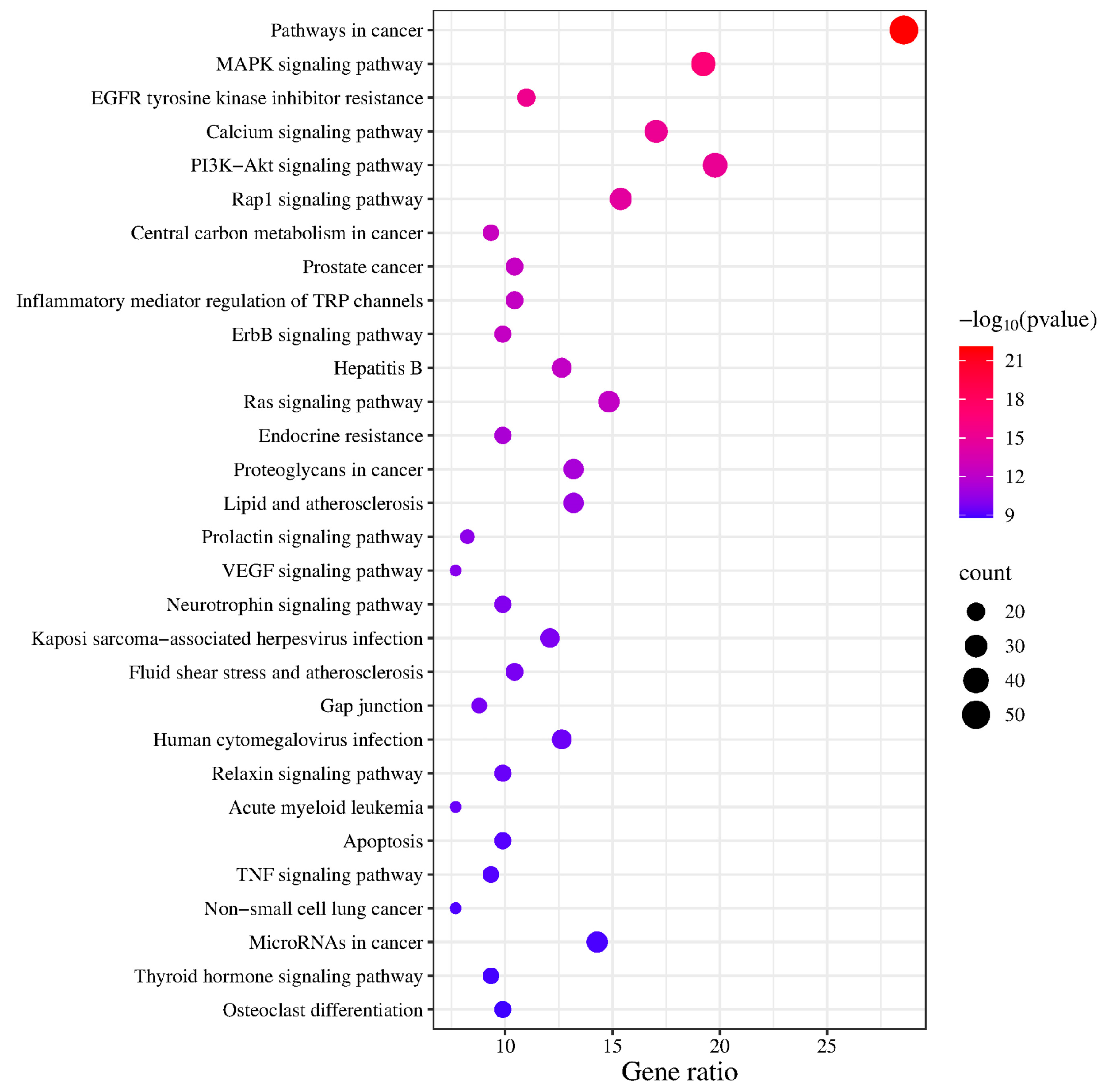
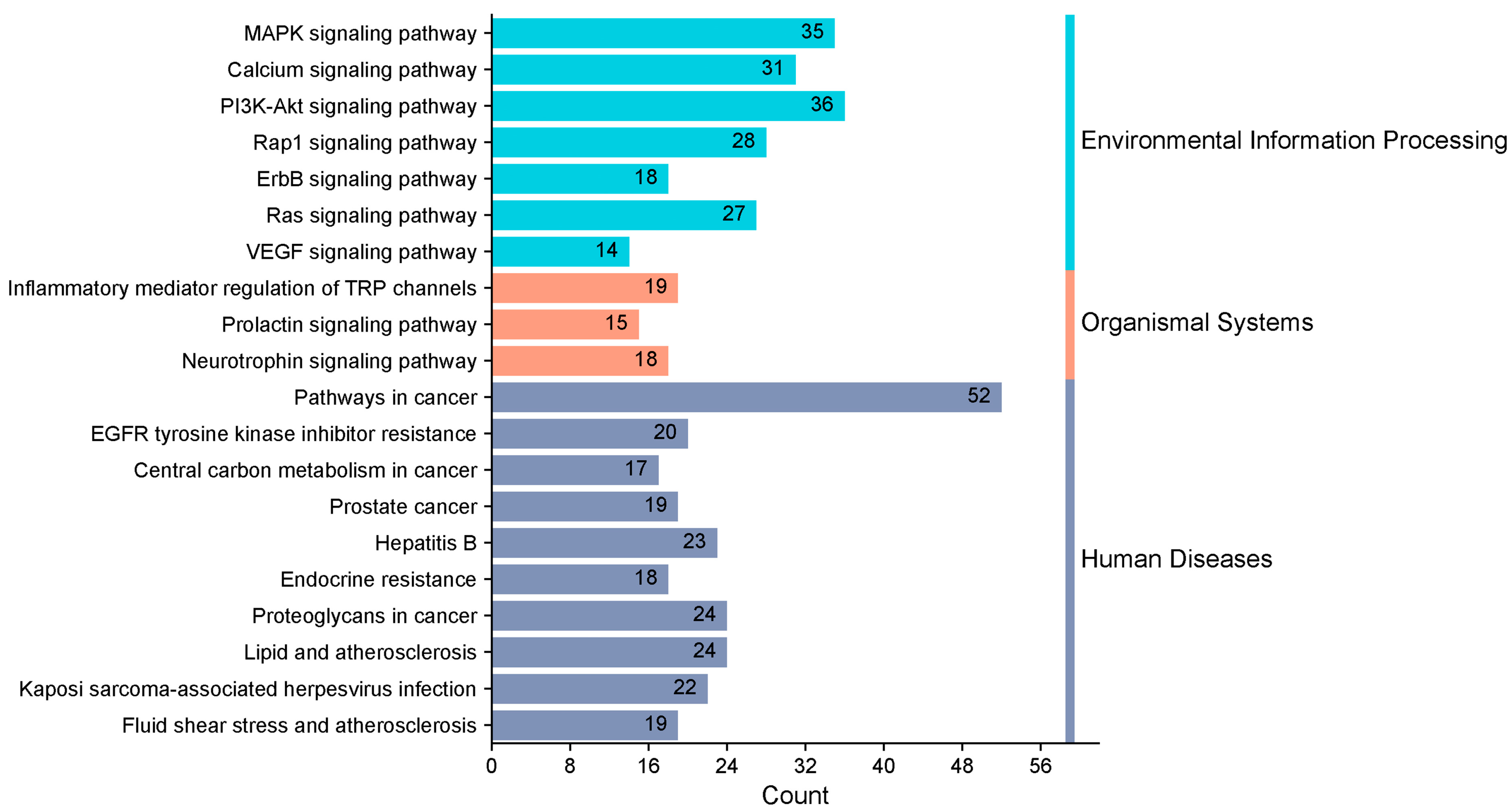
3.3. Target Function Analysis and Pathway Enrichment Analysis
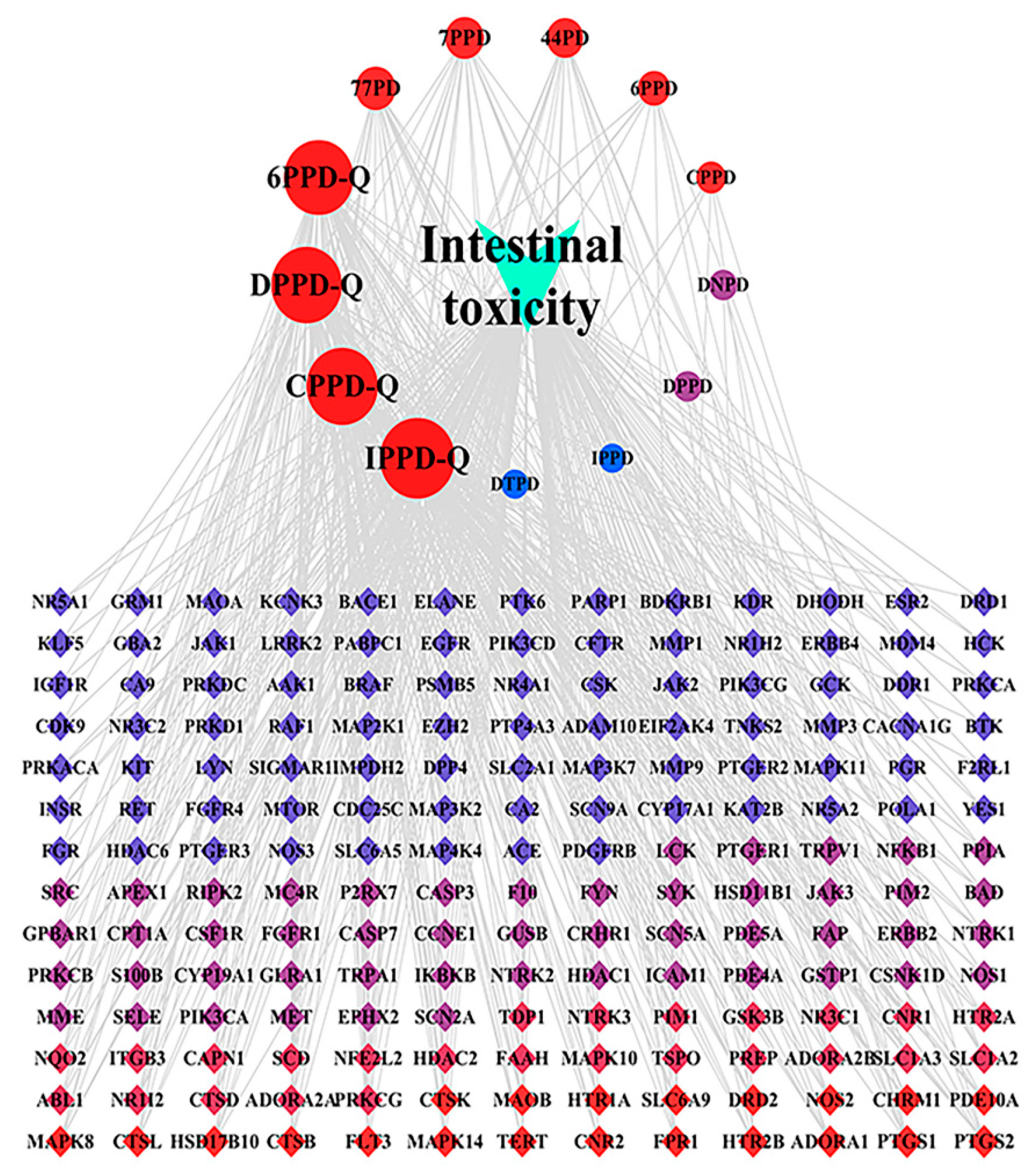
3.4. Analysis of the Intestinal Toxicity Network Induced by PPDs and PPDQs
3.5. Molecular Docking for IPPD-Q and Core Target Proteins of Intestinal Toxicity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, H.; Huang, Z.; Liu, Y.; Hu, L.; He, L.; Liu, Y.; Zhao, J.; Ying, G. Occurrence and risks of 23 tire additives and their transformation products in an urban water system. Environ. Int. 2023, 171, 107715. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Zhang, Z.; Zeb, A.; Fu, X.; Shi, X.; Liu, J.; Wang, J.; Wang, Q.; Chen, C.; Sun, W.; et al. Environmental occurrence, fate, human exposure, and human health risks of p-phenylenediamines and their quinones. Sci. Total. Environ. 2024, 957, 177742. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Li, Y.; Sun, Y.; Liu, L.; Shen, M.; Du, B. Widespread Occurrence and Transport of p-Phenylenediamines and their Quinones in Sediments across Urban Rivers, Estuaries, Coasts, and Deep-Sea Regions. Environ. Sci. Technol. 2023, 57, 2393–2403. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Hu, J.; Yuan, Z.; Wang, S.; Tong, L. P-phenylenediamines (PPDs) and PPD-quinones (PPD-Qs) in human urine and breast milk samples: Urgent need for focus on PPD-Qs and the establishment of health threshold criteria. J. Hazard. Mater. 2024, 480, 136176. [Google Scholar] [CrossRef]
- Varshney, S.; Gora, A.H.; Siriyappagouder, P.; Kiron, V.; Olsvik, P.L.A. Toxicological effects of 6PPD and 6PPD quinone in zebrafish larvae. J. Hazard. Mater. 2022, 424, 127623. [Google Scholar] [CrossRef]
- Peng, W.; Liu, C.; Chen, D.; Duan, X.; Zhong, L. Exposure to N-(1,3-dimethylbutyl)-N′-phenyl-p-phenylenediamine (6PPD) affects the growth and development of zebrafish embryos/larvae. Ecotoxicol. Environ. Saf. 2022, 232, 113221. [Google Scholar] [CrossRef]
- Greer, J.B.; Dalsky, E.M.; Lane, R.F.; Hansen, J.D. Tire-Derived transformation product 6PPD-Quinone induces mortality and transcriptionally disrupts vascular permeability pathways in developing coho salmon. Environ. Sci. Technol. 2023, 57, 10940–10950. [Google Scholar] [CrossRef]
- Ji, J.; Huang, J.; Cao, N.; Hao, X.; Wu, Y.; Ma, Y.; An, D.; Pang, S.; Li, X. Multiview behavior and neurotransmitter analysis of zebrafish dyskinesia induced by 6PPD and its metabolites. Sci. Total Environ. 2022, 838, 156013. [Google Scholar] [CrossRef]
- Hua, X.; Feng, X.; Liang, G.; Chao, J.; Wang, D. Long-term exposure to 6-PPD quinone reduces reproductive capacity by enhancing germline apoptosis associated with activation of both DNA damage and cell corpse engulfment in Caenorhabditis elegans. J. Hazard. Mater. 2023, 454, 131495. [Google Scholar] [CrossRef]
- Fang, L.; Fang, C.; Di, S.; Yu, Y.; Wang, C.; Wang, X.; Jin, Y. Oral exposure to tire rubber-derived contaminant 6PPD and 6PPD-quinone induce hepatotoxicity in mice. Sci. Total Environ. 2023, 869, 161836. [Google Scholar] [CrossRef]
- Sawicki, T.; Ruszkowska, M.; Danielewicz, A.; Wiedzka, E.N.; Ukowicz, T.A.; Owicz, K.E.P. A review of colorectal cancer in terms of epidemiology, risk factors, development, symptoms and diagnosis. Cancers 2021, 13, 2025. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Gan, X.; Shen, B.; Jiang, J.; Shen, H.; Lei, Y.; Liang, Q.; Bai, C.; Huang, C.; Wu, W.; et al. 6PPD and its metabolite 6PPDQ induce different developmental toxicities and phenotypes in embryonic zebrafish. J. Hazard. Mater. 2023, 455, 131601. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liang, G.; Chao, J.; Wang, D. Comparison of intestinal toxicity in enhancing intestinal permeability and in causing ROS production of six PPD quinones in Caenorhabditis elegans. Sci. Total Environ. 2024, 927, 172306. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Qiu, J.; Zhao, Y. Exploring the Potential Toxicological Mechanisms of Vine Tea on the Liver Based on Network Toxicology and Transcriptomics. Front. Pharmacol. 2022, 13, 855926. [Google Scholar] [CrossRef]
- Huang, S. Efficient analysis of toxicity and mechanisms of environmental pollutants with network toxicology and molecular docking strategy: Acetyl tributyl citrate as an example. Sci. Total Environ. 2023, 905, 167904. [Google Scholar] [CrossRef]
- Chen, D.; Hou, X. Aspartame carcinogenic potential revealed through network toxicology and molecular docking insights. Sci. Rep. 2024, 14, 11492. [Google Scholar] [CrossRef]
- Cheng, M.; Li, M.; Zhang, Y.; Gu, X.; Gao, W.; Zhang, S.; Liu, J. Exploring the mechanism of PPCPs on human metabolic diseases based on network toxicology and molecular docking. Environ. Int. 2025, 196, 109324. [Google Scholar] [CrossRef]
- Tao, W.; Xu, X.; Wang, X.; Li, B.; Wang, Y.; Li, Y.; Yang, L. Network pharmacology-based prediction of the active ingredients and potential targets of Chinese herbal Radix Curcumae formula for application to cardiovascular disease. J. Ethnopharmacol. 2013, 145, 1–10. [Google Scholar] [CrossRef]
- He, J.; Zhu, X.; Xu, K.; Li, Y.; Zhou, J. Network toxicological and molecular docking to investigate the mechanisms of toxicity of agricultural chemical Thiabendazole. Chemosphere 2024, 363, 142711. [Google Scholar] [CrossRef]
- Villalobo, A. Ca2+ signaling and src functions in tumor cells. Biomolecules 2023, 13, 1739. [Google Scholar] [CrossRef]
- Samak, G.; Chaudhry, K.K.; Gangwar, R.; Narayanan, D.; Jaggar, J.H.; Rao, R. Calcium/Ask1/MKK7/JNK2/c-Src signalling cascade mediates disruption of intestinal epithelial tight junctions by dextran sulfate sodium. Biochem. J. 2015, 465, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Elfiky, A.; Han, M.; Chen, C.; Saif, M.W. The role of src in colon cancer and its therapeutic implications. Clin. Color. Cancer 2014, 13, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Simatou, A.; Simatos, G.; Goulielmaki, M.; Spandidos, D.; Baliou, S.; Zoumpourlis, V. Historical retrospective of the SRC oncogene and new perspectives (Review). Mol. Clin. Oncol. 2020, 13, 21. [Google Scholar] [CrossRef]
- Jin, W. Regulation of src family kinases during colorectal cancer development and its clinical implications. Cancers 2020, 12, 1339. [Google Scholar] [CrossRef]
- Kang, X.; Li, R.; Li, X.; Xu, X. EGFR mutations and abnormal trafficking in cancers. Mol. Biol. Rep. 2024, 51, 924. [Google Scholar] [CrossRef]
- Zhou, J.; Ji, Q.; Li, Q. Resistance to anti-EGFR therapies in metastatic colorectal cancer: Underlying mechanisms and reversal strategies. J. Exp. Clin. Cancer Res. 2021, 40, 328. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Shin, D.; Kim, J.W.; Keam, B.; Kim, T.M.; Kim, H.J.; Kim, D.-W.; Wu, H.-G.; Paek, S.H.; Kim, Y.W.; et al. Additional prognostic role of EGFR activating mutations in lung adenocarcinoma patients with brain metastasis: Integrating with lung specific GPA score. Lung Cancer 2014, 86, 363–368. [Google Scholar] [CrossRef]
- Napolitano, S.; Martini, G.; Ciardiello, D.; Del Tufo, S.; Martinelli, E.; Troiani, T.; Ciardiello, P.F. Targeting the EGFR signalling pathway in metastatic colorectal cancer. The lancet. Gastroenterol. Hepatol. 2024, 9, 664. [Google Scholar] [CrossRef]
- Xie, Y.; Chen, Y.; Fang, J. Comprehensive review of targeted therapy for colorectal cancer. Signal Transduct. Target. Ther. 2020, 5, 22. [Google Scholar] [CrossRef]
- Martinelli, E.; Ciardiello, D.; Martini, G.; Troiani, T.; Cardone, C.; Vitiello, P.P.; Normanno, N.; Rachiglio, A.M.; Maiello, E.; Latiano, T.; et al. Implementing anti-epidermal growth factor receptor (EGFR) therapy in metastatic colorectal cancer: Challenges and future perspectives. Ann. Oncol. 2020, 31, 30–40. [Google Scholar] [CrossRef]
- Eskandari, E.; Eaves, C.J. Paradoxical roles of caspase-3 in regulating cell survival, proliferation, and tumorigenesis. J. Cell Biol. 2022, 221, e202201159. [Google Scholar] [CrossRef]
- Galluzzi, L.; López-Soto, A.; Kumar, S.; Kroemer, G. Caspases connect Cell-Death signaling to organismal homeostasis. Immunity 2016, 44, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.A.; Thapa, R.; Afzal, O.; Agrawal, N.; Almalki, W.H.; Kazmi, I.; Alzarea, S.I.; Altamimi, A.S.A.; Prasher, P.; Singh, S.K.; et al. The pyroptotic role of Caspase-3/GSDME signalling pathway among various cancer: A Review. Int. J. Biol. Macromol. 2023, 242, 124832. [Google Scholar] [CrossRef]
- Zhou, M.; Liu, X.; Li, Z.; Huang, Q.; Li, F.; Li, C.Y. Caspase-3 regulates the migration, invasion and metastasis of colon cancer cells. Int. J. Cancer 2018, 143, 921–930. [Google Scholar] [CrossRef]
- Flanagan, L.; Meyer, M.; Fay, J.; Curry, S.; Bacon, O.; Duessmann, H.; John, K.; Boland, K.C.; McNamara, D.A.; Kay, E.W.; et al. Low levels of Caspase-3 predict favourable response to 5FU-based chemotherapy in advanced colorectal cancer: Caspase-3 inhibition as a therapeutic approach. Cell Death Dis. 2016, 7, e2087. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Ou, J.; Zhang, C.; Chen, J.; Chen, J.; Li, A.; Huang, B.; Zhao, X. Rapamycin promotes the intestinal barrier repair in ulcerative colitis via the mTOR/PBLD/AMOT signaling pathway. Biochim. Biophys. Acta Mol. Basis Dis. 2024, 1870, 167287. [Google Scholar] [CrossRef]
- Taylor, H.; Yerlioglu, D.; Phen, C.; Ballauff, A.; Nedelkopoulou, N.; Spier, I.; Loverdos, I.; Busoni, V.B.; Heise, J.; Dale, P.; et al. MTOR inhibitors reduce enteropathy, intestinal bleeding and colectomy rate in patients with juvenile polyposis of infancy with PTEN-BMPR1A deletion. Hum. Mol. Genet. 2021, 30, 1273–1282. [Google Scholar] [CrossRef]
- Pochard, C.; Gonzales, J.; Bessard, A.; Mahe, M.M.; Bourreille, A.; Cenac, N.; Jarry, A.; Coron, E.; Podevin, J.; Meurette, G.; et al. PGI2 inhibits intestinal epithelial permeability and apoptosis to alleviate colitis. Cell Mol. Gastroenterol. Hepatol. 2021, 12, 1037–1060. [Google Scholar] [CrossRef] [PubMed]
- Hynes, N.E.; Lane, H.A. ERBB receptors and cancer: The complexity of targeted inhibitors. Nat. Rev. Cancer 2005, 5, 341–354. [Google Scholar] [CrossRef]
- Mangiapane, L.R.; Nicotra, A.; Turdo, A.; Gaggianesi, M.; Bianca, P.; Di Franco, S.; Sardina, D.S.; Veschi, V.; Signore, M.; Beyes, S.; et al. PI3K-driven HER2 expression is a potential therapeutic target in colorectal cancer stem cells. Gut 2021, 71, 119–128. [Google Scholar] [CrossRef]
- Le, T.; Galmiche, L.; Levy, J.; Suwannarat, P.; Hellebrekers, D.M.E.I.; Morarach, K.; Boismoreau, F.; Theunissen, T.E.J.; Lefebvre, M.; Pelet, A.; et al. Dysregulation of the NRG1/ERBB pathway causes a developmental disorder with gastrointestinal dysmotility in humans. J. Clin. Investig. 2021, 131, e145837. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Akash, S.; Rahman, M.M.; Nowrin, F.T.; Akter, T.; Shohag, S.; Rauf, A.; Aljohani, A.S.M.; Simal-Gandara, J. Colon cancer and colorectal cancer: Prevention and treatment by potential natural products. Chem. Biol. Interact. 2022, 368, 110170. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Baek, K. E3 ligases and deubiquitinating enzymes regulating the MAPK signaling pathway in cancers. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188736. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Fang, Y.; Li, Y.; Au, R.; Cheng, C.; Li, W.; Xu, F.; Cui, Y.; Zhu, L.; Shen, H. A network pharmacology approach and experimental validation to investigate the anticancer mechanism of Qi-Qin-Hu-Chang formula against colitis-associated colorectal cancer through induction of apoptosis via JNK/p38 MAPK signaling pathway. J. Ethnopharmacol. 2024, 319, 117323. [Google Scholar] [CrossRef]
- Zhou, G.; Yang, J.; Song, P. Correlation of ERK/MAPK signaling pathway with proliferation and apoptosis of colon cancer cells. Oncol. Lett. 2018, 17, 2266–2270. [Google Scholar] [CrossRef]
- Humeau, J.; Pedro, J.M.B.; Vitale, I.; Ez, L.N.; Villalobos, C.; Kroemer, G.; Senovilla, L. Calcium signaling and cell cycle: Progression or death. Cell Calcium 2018, 70, 3–15. [Google Scholar] [CrossRef]
- Wu, J.; Shao, Y.; Huang, C.; Wang, Z.; Zhang, H.; Fu, Z. Genetic variants in the calcium signaling pathway participate in the pathogenesis of colorectal cancer through the tumor microenvironment. Front. Oncol. 2023, 13, 992326. [Google Scholar] [CrossRef]
- Lieu, C.; Kopetz, S. The src family of protein tyrosine kinases: A new and promising target for colorectal cancer therapy. Clin. Color. Cancer 2010, 9, 89–94. [Google Scholar] [CrossRef]
- Sen, B.; Saigal, B.; Parikh, N.; Gallick, G.; Johnson, F.M. Sustained src inhibition results in STAT3 activation and cancer cell survival via altered JAK-STAT3 binding. Cancer Res. 2009, 69, 1958–1965. [Google Scholar] [CrossRef]
- Biscardi, J.S.; Maa, M.; Tice, D.A.; Cox, M.E.; Leu, T.; Parsons, S.J. C-Src-mediated phosphorylation of the epidermal growth factor receptor on tyr845 and tyr1101 is associated with modulation of receptor function. J. Biol. Chem. 1999, 274, 8335–8343. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Degree | Closeness Centrality | Betweenness Centrality | Topological Coefficlent |
|---|---|---|---|---|
| SRC | 88 | 0.655677 | 0.10274 | 0.166128 |
| EGFR | 86 | 0.646209 | 0.070655 | 0.177292 |
| CASP3 | 85 | 0.646209 | 0.067594 | 0.176605 |
| MTOR | 69 | 0.610921 | 0.05981 | 0.191718 |
| ERBB2 | 69 | 0.606779 | 0.051657 | 0.192028 |
| NFKB1 | 69 | 0.604729 | 0.045221 | 0.19494 |
| MMP9 | 63 | 0.594684 | 0.040975 | 0.191118 |
| PTGS2 | 60 | 0.588815 | 0.037339 | 0.188631 |
| GSK3B | 59 | 0.583061 | 0.033291 | 0.196352 |
| PIK3CA | 57 | 0.573717 | 0.029618 | 0.2106307 |
| PRKACA | 54 | 0.571884 | 0.028589 | 0.152588 |
| PARP1 | 48 | 0.559375 | 0.027703 | 0.212075 |
| MAPK14 | 48 | 0.557632 | 0.019497 | 0.217721 |
| KDR | 46 | 0.5559 | 0.019177 | 0.217391 |
| ABL1 | 46 | 0.549079 | 0.019049 | 0.223273 |
| JAK2 | 45 | 0.549079 | 0.01485 | 0.238955 |
| IGF1R | 44 | 0.5474 | 0.014761 | 0.231476 |
| MAPK8 | 42 | 0.5474 | 0.01471 | 0.21956 |
| FYN | 42 | 0.545731 | 0.014464 | 0.230064 |
| PGR | 41 | 0.545731 | 0.014268 | 0.231276 |
| BRAF | 41 | 0.544072 | 0.013927 | 0.231997 |
| ICAM1 | 40 | 0.534328 | 0.01352 | 0.242438 |
| NR3C1 | 39 | 0.532738 | 0.012948 | 0.212789 |
| LYN | 38 | 0.531157 | 0.012811 | 0.219864 |
| PIK3CG | 37 | 0.529585 | 0.012061 | 0.226415 |
| PRKCA | 37 | 0.528023 | 0.011536 | 0.233894 |
| NTRK1 | 37 | 0.524926 | 0.01141 | 0.240453 |
| PIK3CD | 37 | 0.5249268 | 0.011282 | 0.241419 |
| SYK | 36 | 0.523391 | 0.011099 | 0.260243 |
| KIT | 36 | 0.523391 | 0.011074 | 0.263979 |
| Serial Number | Chemical Components | Degree |
|---|---|---|
| 1 | IPPD-Q | 73 |
| 2 | CPPD-Q | 68 |
| 3 | DPPD-Q | 67 |
| 4 | 6PPD-Q | 65 |
| 5 | 77PD | 21 |
| 6 | 7PPD | 19 |
| 7 | 44PD | 16 |
| 8 | 6PPD | 9 |
| 9 | CPPD | 5 |
| 10 | DNPD | 3 |
| Gene | PDB ID |
|---|---|
| SRC | 1A07 |
| EGFR | 1M14 |
| CASP3 | 1GFW |
| MTOR | 1AUE |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, H.; Tan, Y.; Ren, X.; Li, Z.; Liu, Y. Network Toxicology and Molecular Docking to Elucidate the Mechanisms of Intestinal Toxicity Induced by P-Phenylenediamine Antioxidants and Their Quinone Derivatives. Toxics 2025, 13, 480. https://doi.org/10.3390/toxics13060480
Zou H, Tan Y, Ren X, Li Z, Liu Y. Network Toxicology and Molecular Docking to Elucidate the Mechanisms of Intestinal Toxicity Induced by P-Phenylenediamine Antioxidants and Their Quinone Derivatives. Toxics. 2025; 13(6):480. https://doi.org/10.3390/toxics13060480
Chicago/Turabian StyleZou, Hui, Yumei Tan, Xiyi Ren, Zhu Li, and Yongxiang Liu. 2025. "Network Toxicology and Molecular Docking to Elucidate the Mechanisms of Intestinal Toxicity Induced by P-Phenylenediamine Antioxidants and Their Quinone Derivatives" Toxics 13, no. 6: 480. https://doi.org/10.3390/toxics13060480
APA StyleZou, H., Tan, Y., Ren, X., Li, Z., & Liu, Y. (2025). Network Toxicology and Molecular Docking to Elucidate the Mechanisms of Intestinal Toxicity Induced by P-Phenylenediamine Antioxidants and Their Quinone Derivatives. Toxics, 13(6), 480. https://doi.org/10.3390/toxics13060480