
Impact of Perinatal Coexposure to Chlorpyrifos and a High-Fat Diet on Kisspeptin and GnRHR Presence and Reproductive Organs

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
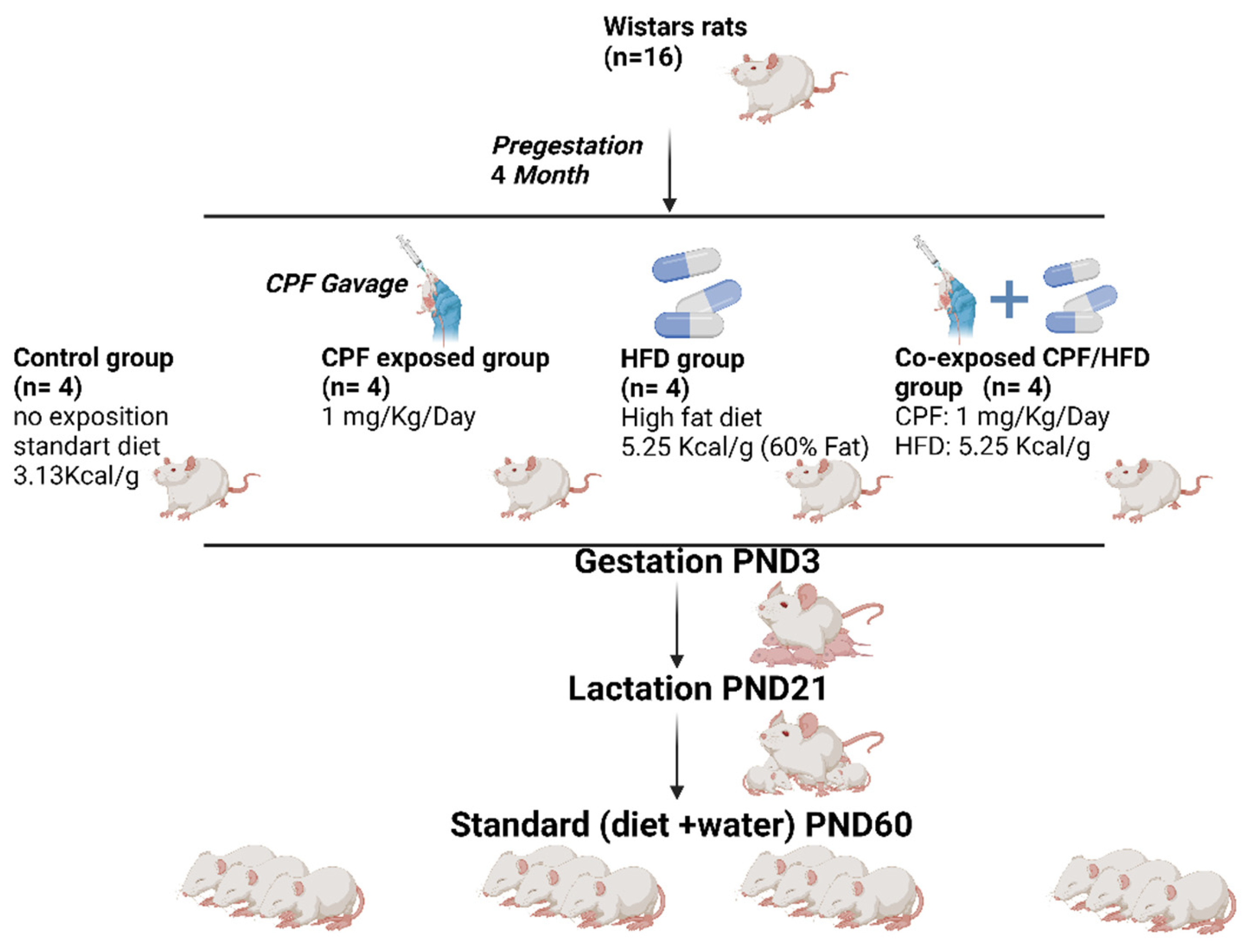
2.1. Chlorpyrifos Preparation and High-Fat Diet
2.2. Experimental Design
2.2.1. Animals Housing
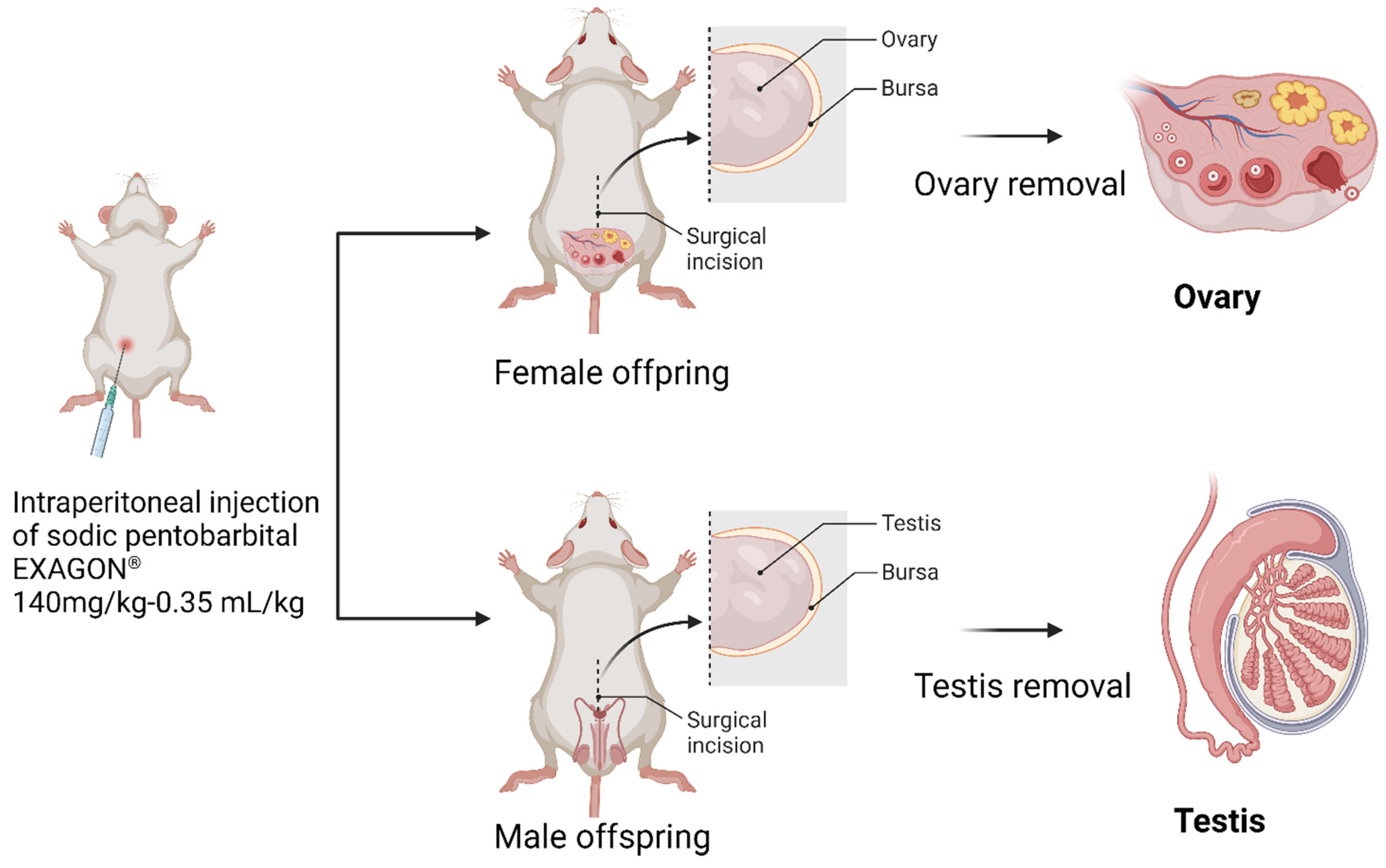
2.2.2. Ethical Approval and Euthanasia Method
2.3. Immunohistochemistry (Kisspeptin and GnRHR)
- -
- For Kisspeptin detection: primary rabbit polyclonal anti Kisspeptin antibody (1:300 dilution, product no. 251265; Abbiotec. Escondido, CA, USA)
- -
- For GnRHR detection: primary rabbit polyclonal to GnRHR (1:200 dilution; abcam, ab202848, Cambridge, UK)
2.4. Histological Analysis
2.5. In Silico Quantification of Expression
2.6. Statistical Analysis
3. Results
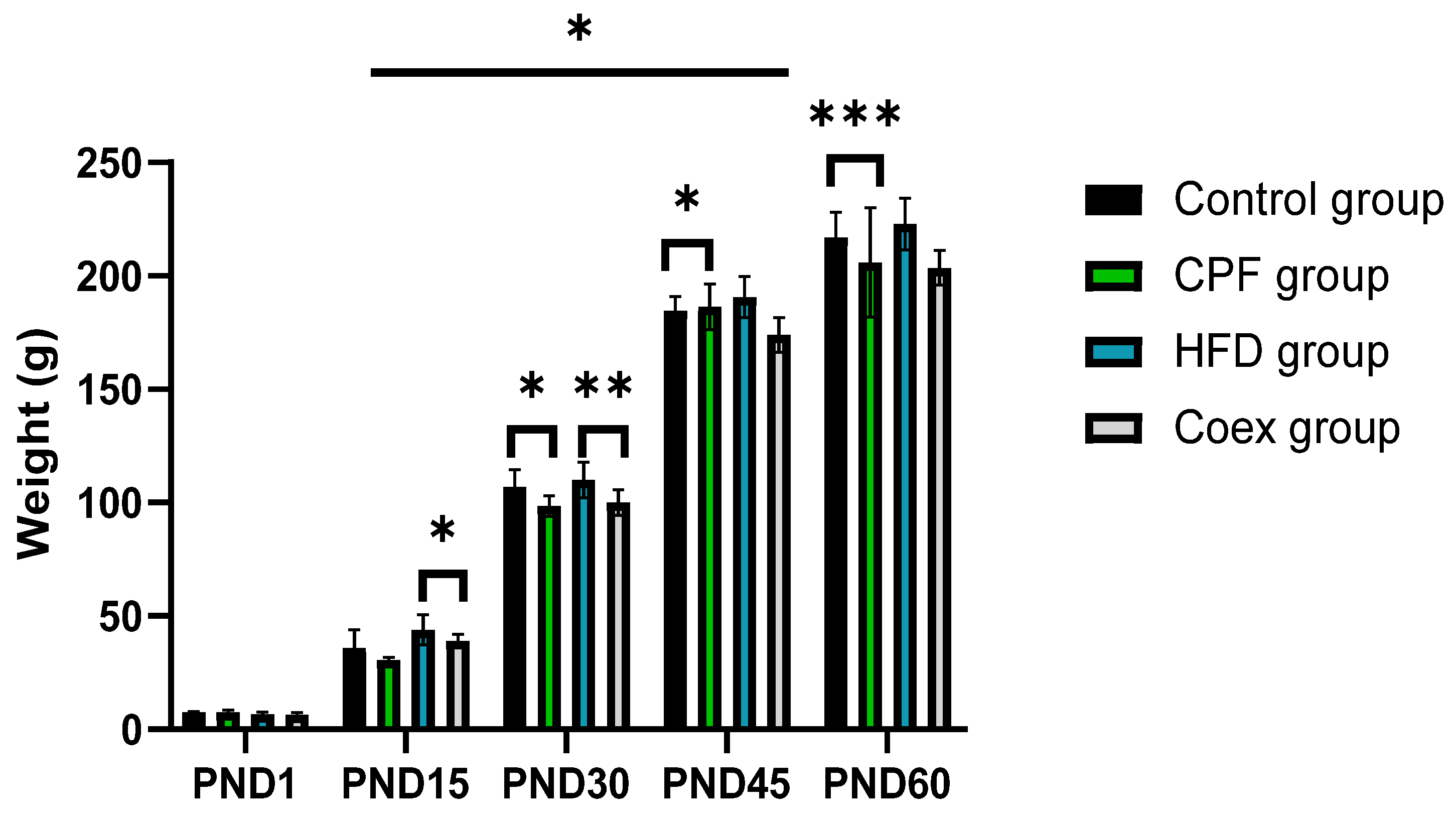
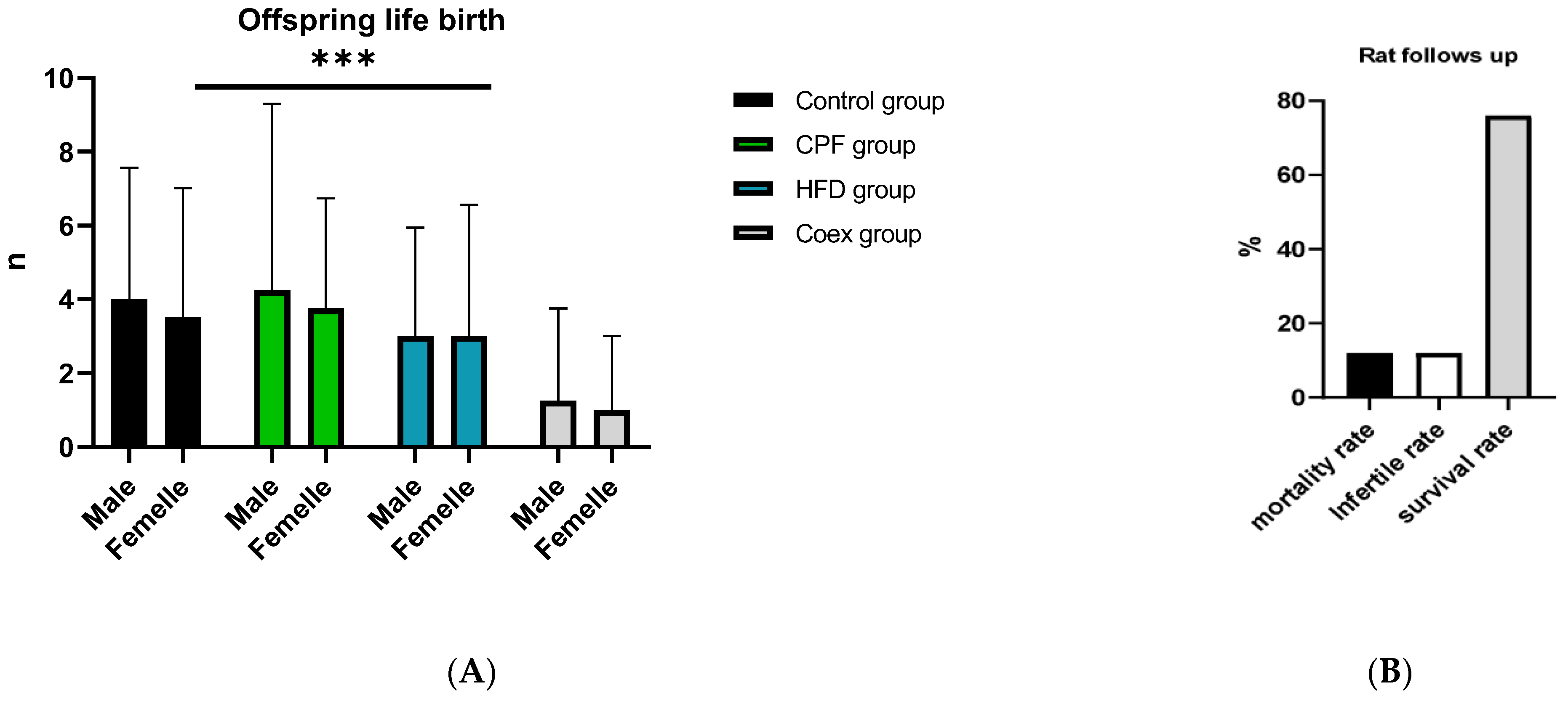
3.1. Impacts of Treatment on Pups’ Body Weight
3.2. Histological Analysis
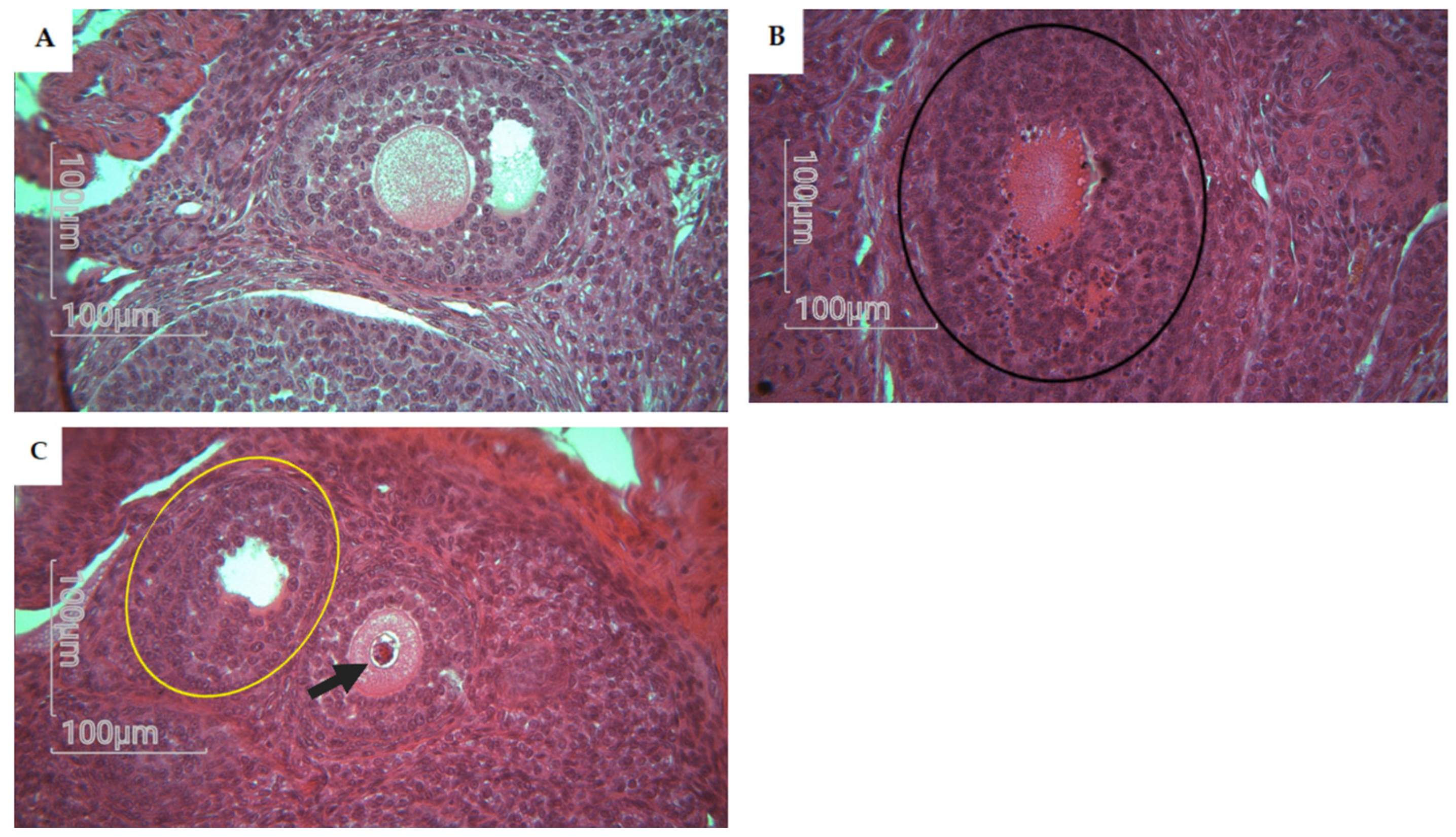
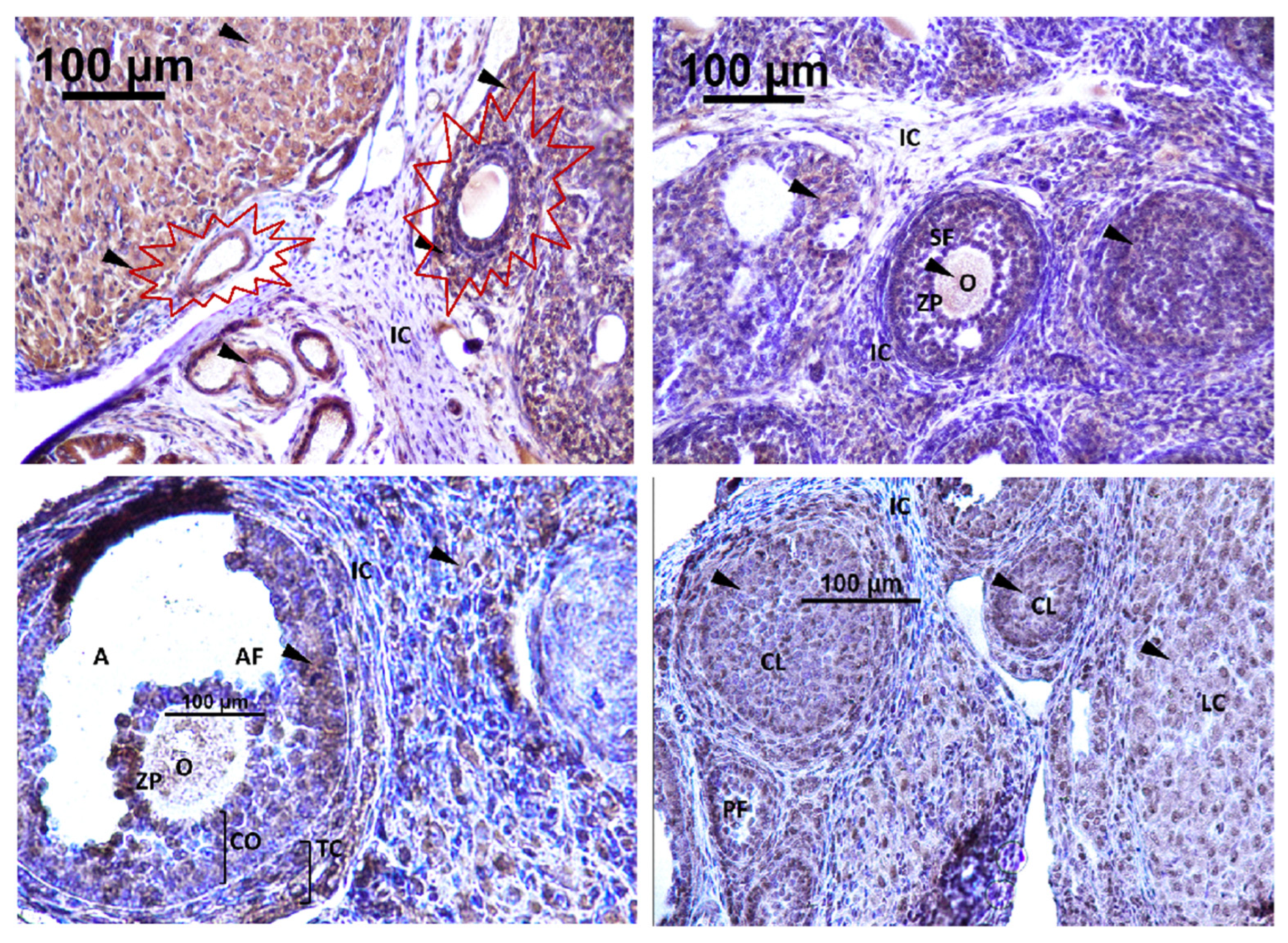
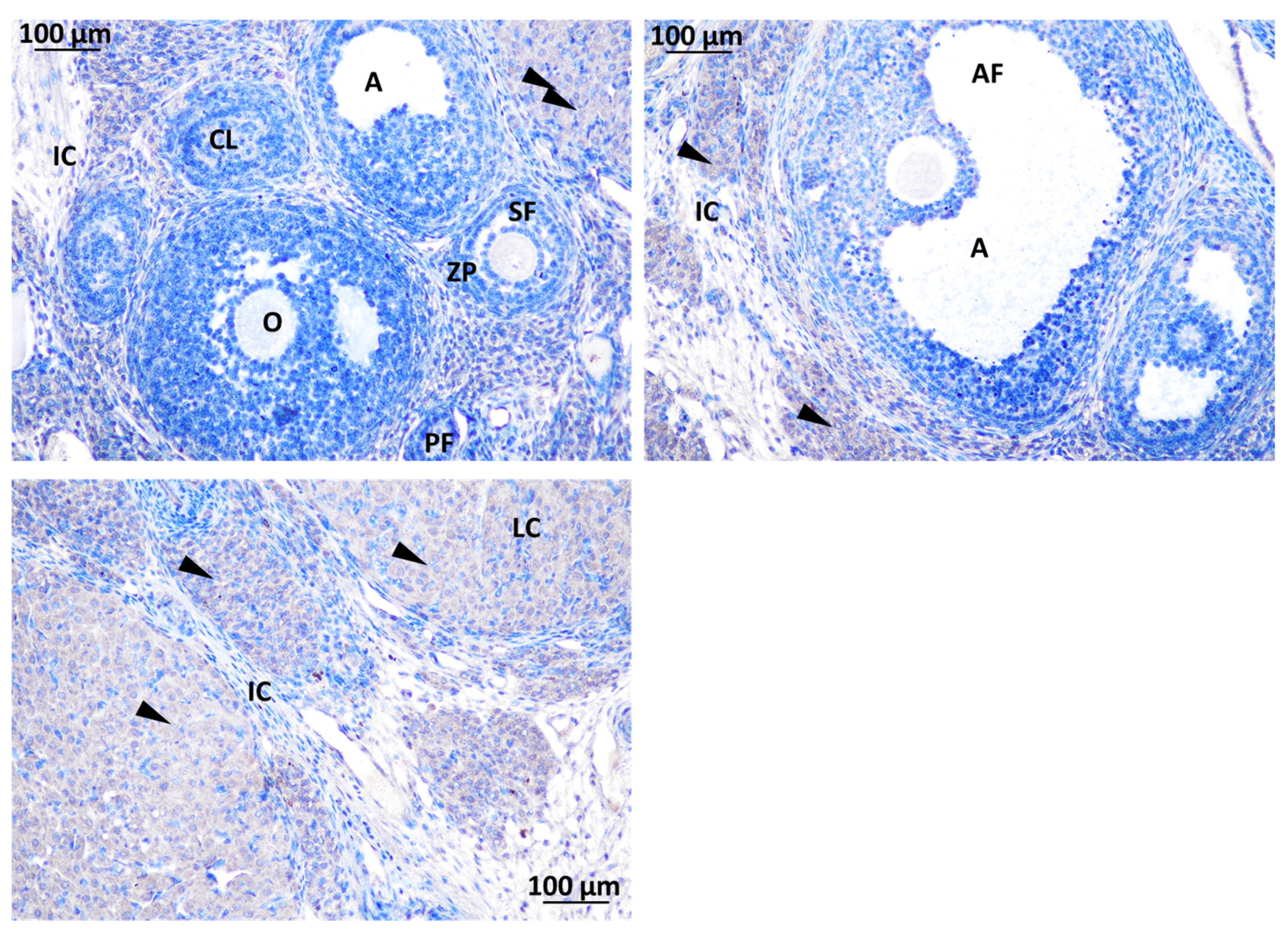
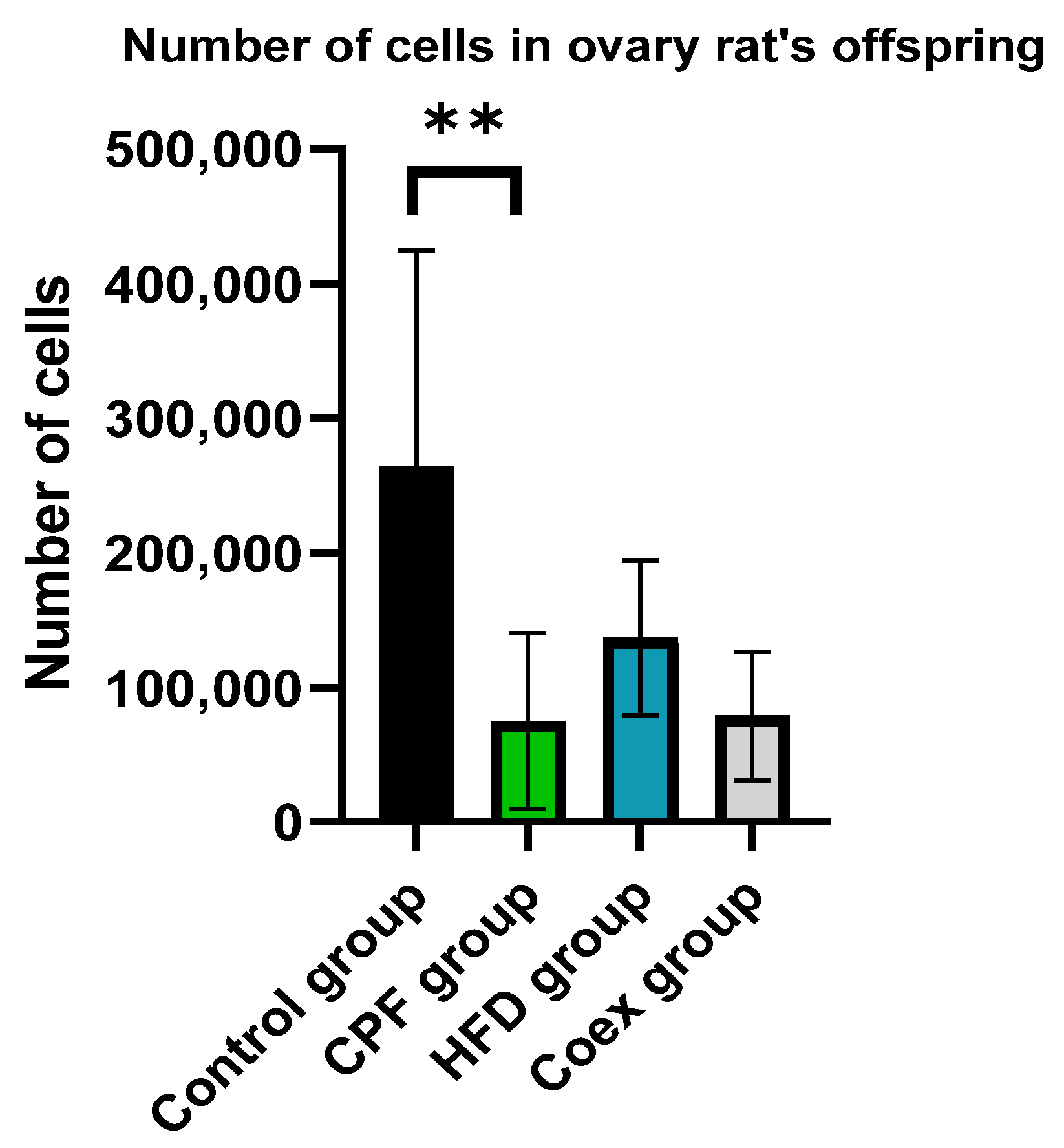
3.2.1. Ovary of Rat Offspring
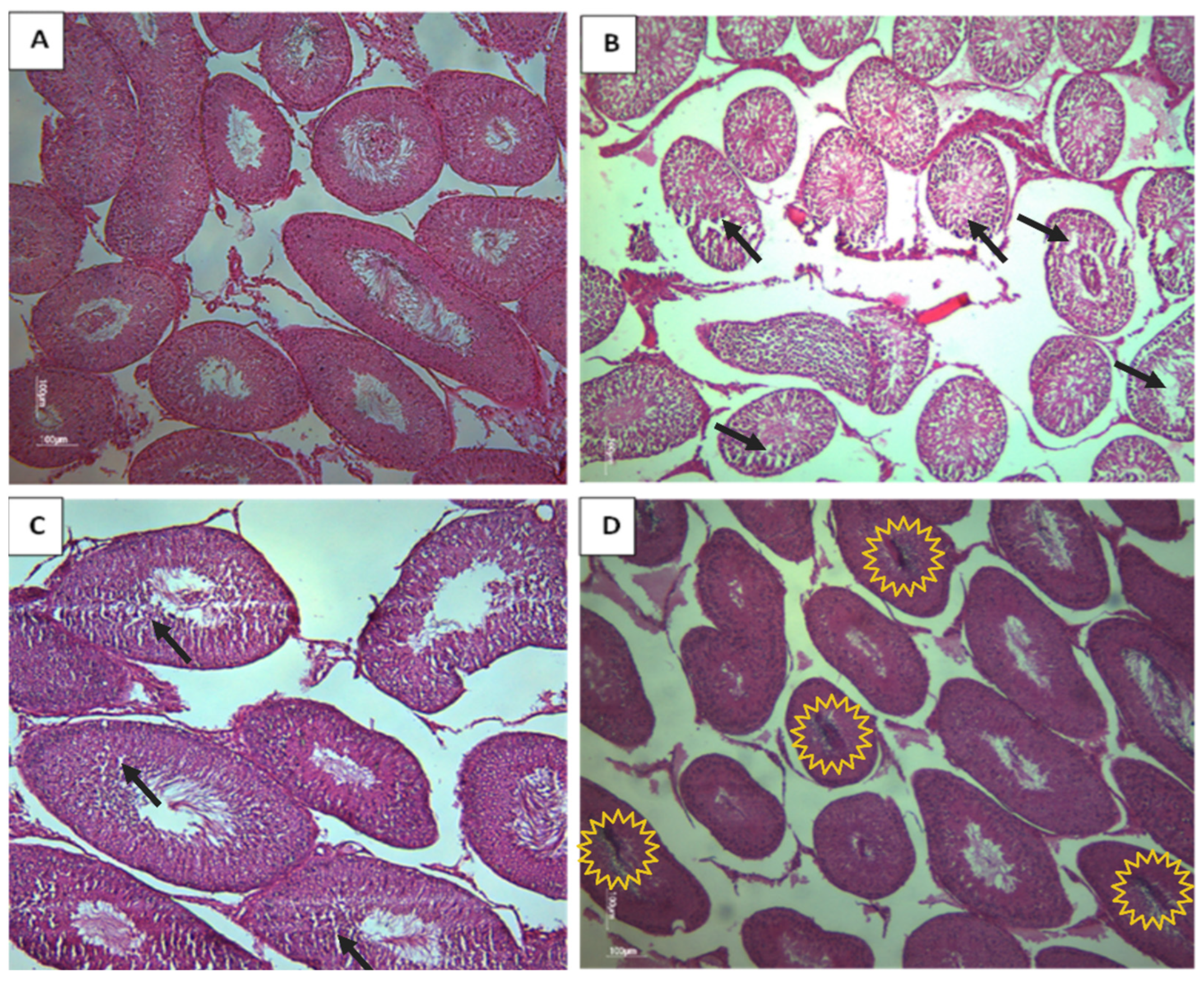
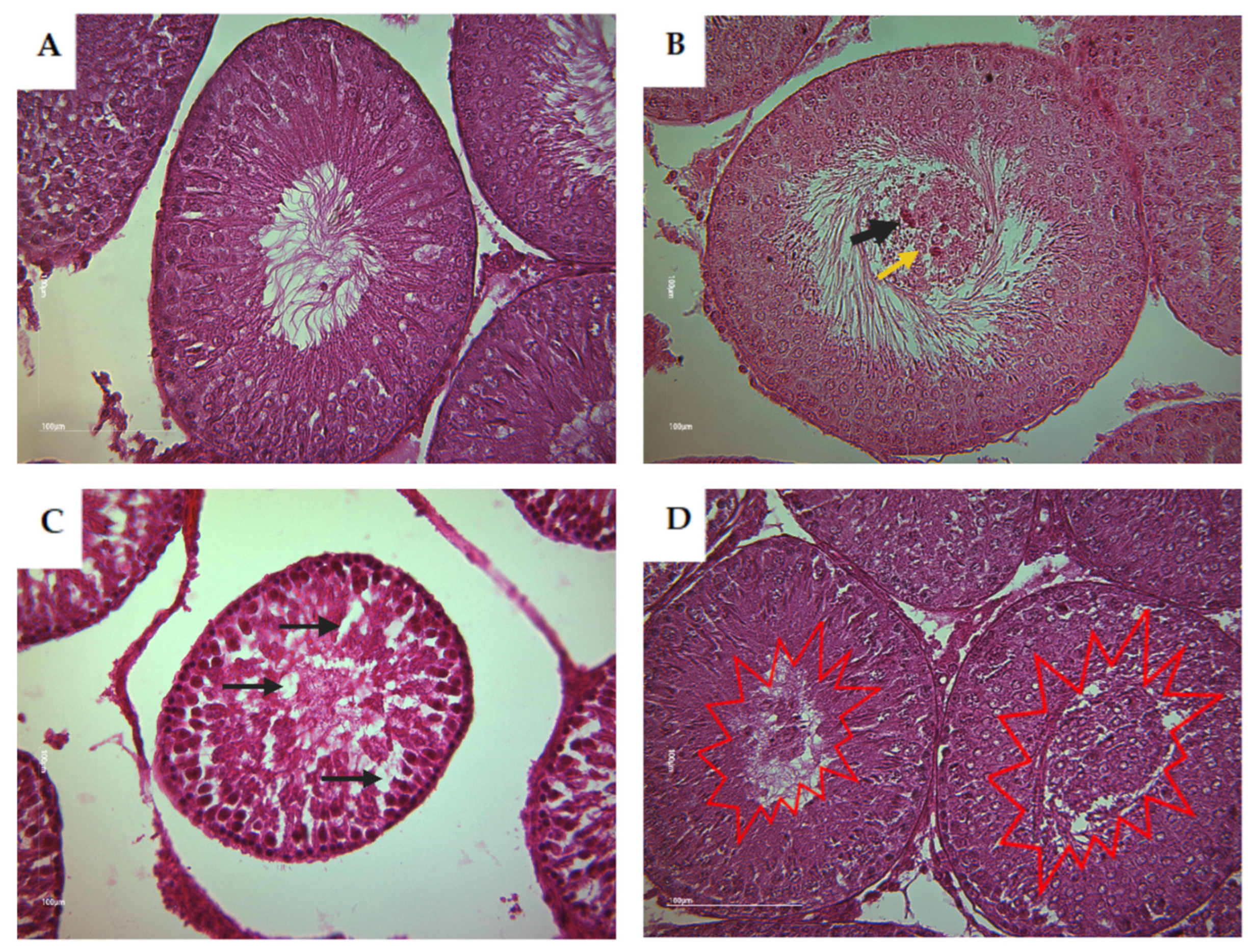
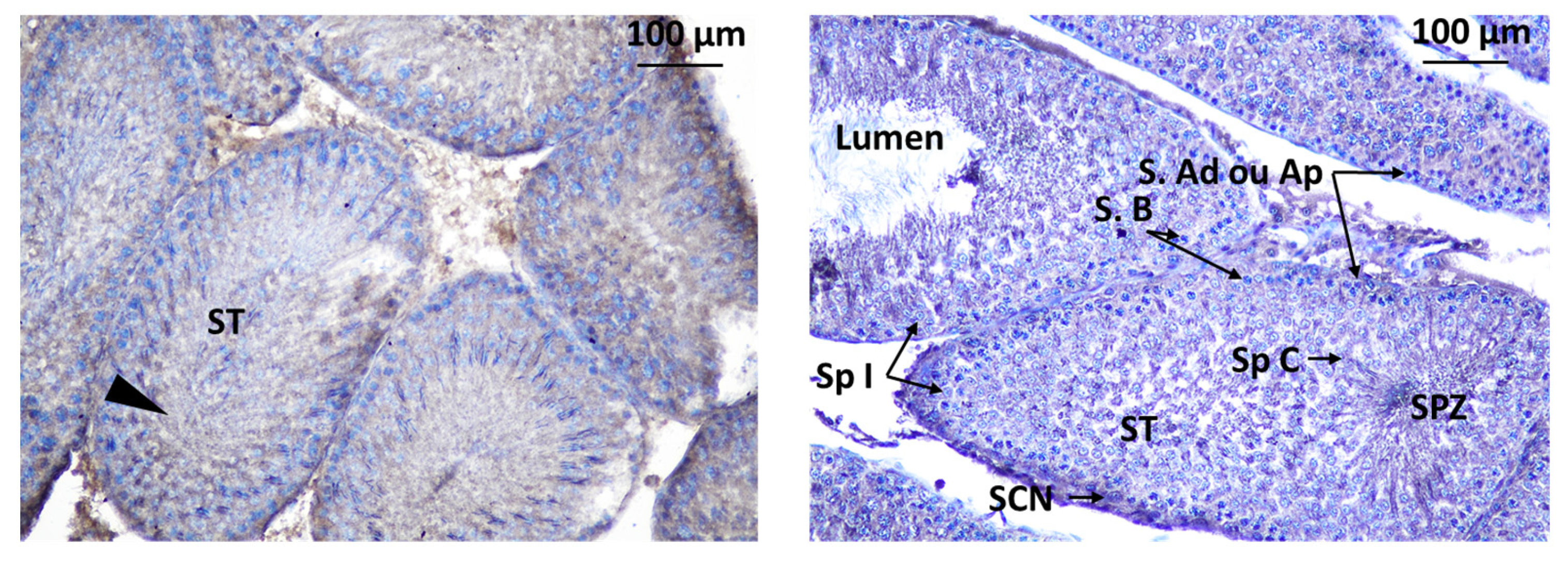
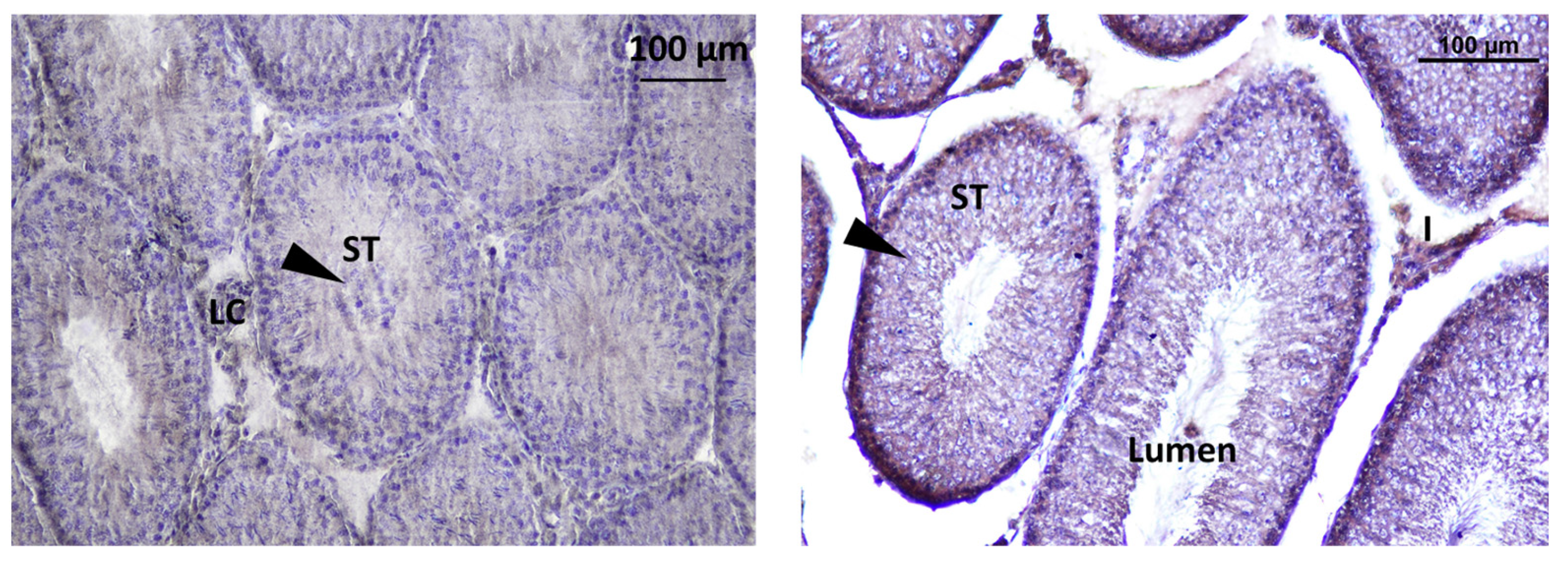
3.2.2. Testis of Rat Offspring
3.3. Kisspeptin and GnRHR Immunodetection
3.3.1. Localization of Immunoreactive Kisspeptin and GnRHR in the Ovary of Rat Offspring
3.3.2. Localization of Immunoreactive Kisspeptin and GnRHR in the Testis of Rat Offspring
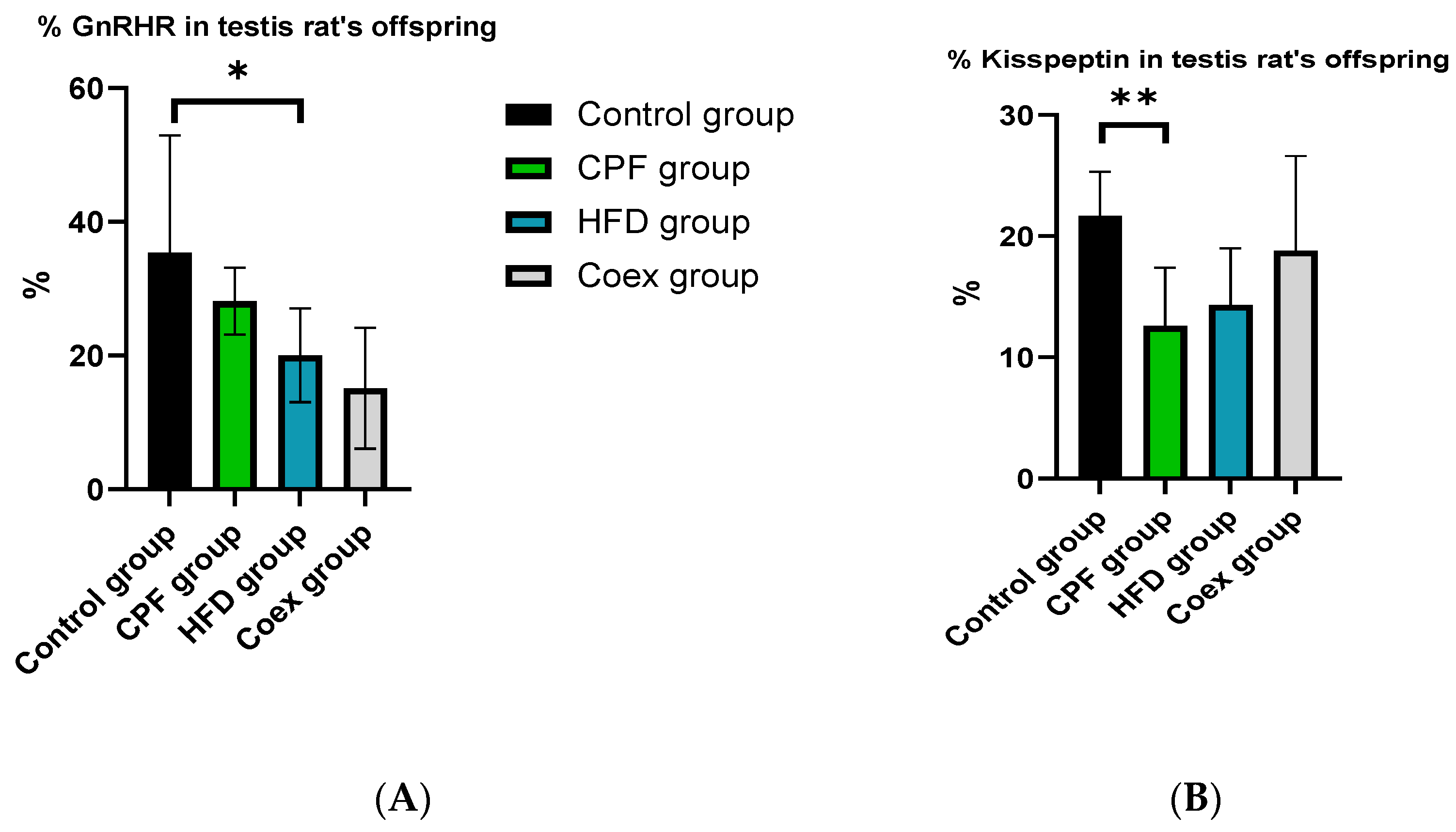
3.4. In Silico Quantification of the Expression of Kisspeptin and GnRHR
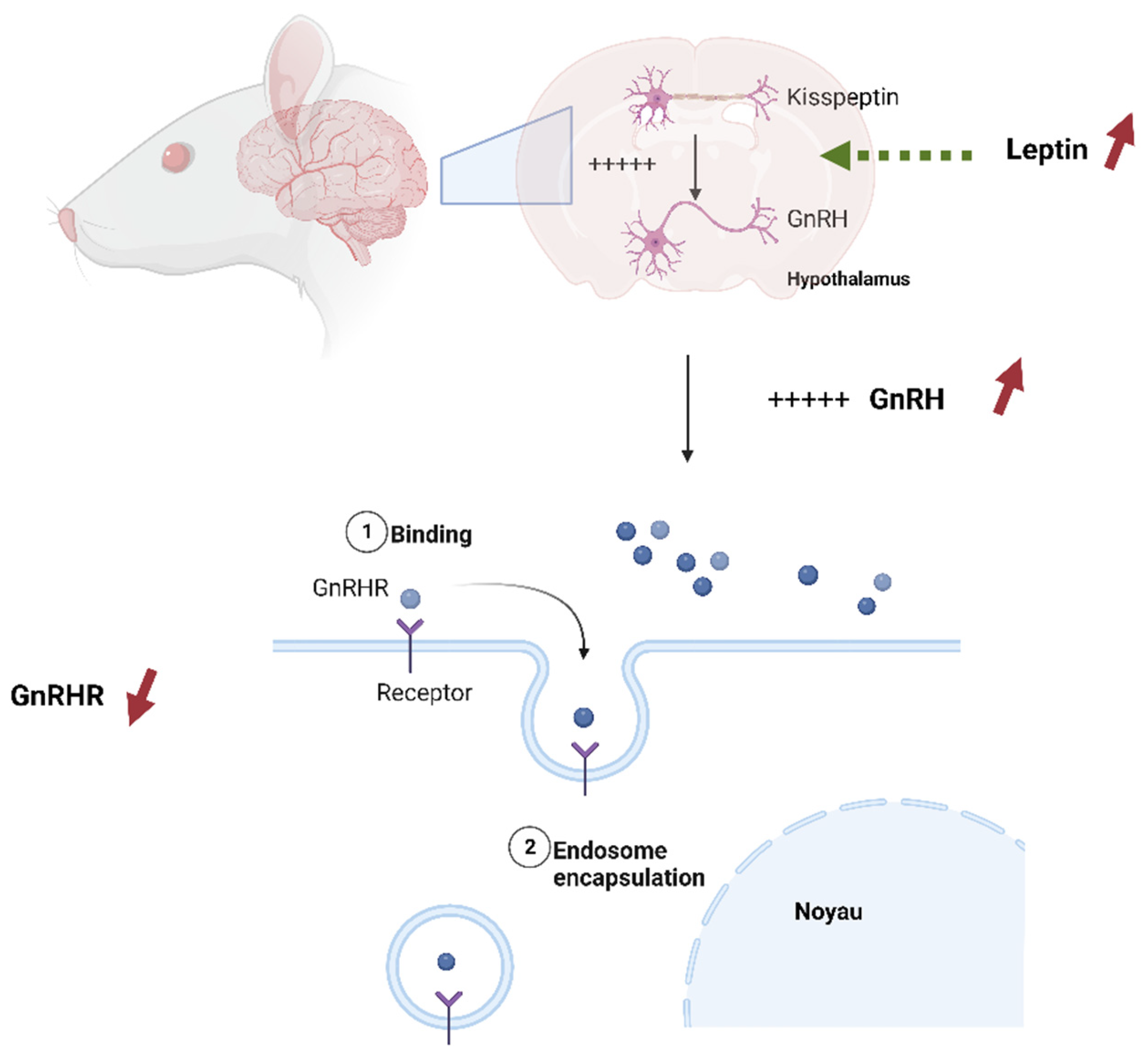
4. Discussion
5. Limitations and Perspectives
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cooper, T.G.; Noonan, E.; Von Eckardstein, S.; Auger, J.; Gordon Baker, H.W.; Behre, H.M.; Haugen, T.B.; Kruger, T.; Wang, C.; Mbizvo, M.T.; et al. World Health Organization reference values for human semen characteristics. Hum. Reprod. Update 2010, 16, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Baker, K.C.; Steiner, A.Z.; Hansen, K.R.; Barnhart, K.T.; Cedars, M.I.; Legro, R.S.; Diamond, M.P.; Krawetz, S.A.; Usadi, R.; Baker, V.L.; et al. Poor reproducibility of percentage of normally shaped sperm using the World Health Organization Fifth Edition strict grading criteria. FS Rep. 2022, 3, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 2015, 13, 37. [Google Scholar] [CrossRef] [PubMed]
- Infertility around the Globe: New Thinking on Gender, Reproductive Technologies and Global Movements in the 21st Century|Human Reproduction Update|Oxford Academic. Available online: https://academic.oup.com/humupd/article/21/4/411/683746 (accessed on 12 September 2022).
- Noppakun, K.; Juntarawijit, C. Association between pesticide exposure and obesity: A cross-sectional study of 20,295 farmers in Thailand. F1000Research 2022, 10, 445. [Google Scholar] [CrossRef]
- Djekkoun, N.; Lalau, J.-D.; Bach, V.; Depeint, F.; Khorsi-Cauet, H. Chronic oral exposure to pesticides and their consequences on metabolic regulation: Role of the microbiota. Eur. J. Nutr. 2021, 60, 4131–4149. [Google Scholar] [CrossRef] [PubMed]
- Segal, T.R.; Giudice, L.C. Before the beginning: Environmental exposures and reproductive and obstetrical outcomes. Fertil. Steril. 2019, 112, 613–621. [Google Scholar] [CrossRef]
- Lahimer, M.; Capelle, S.; Lefranc, E.; Cabry, R.; Montjean, D.; Bach, V.; Ajina, M.; Ben Ali, H.; Benkhalifa, M.; Khorsi-Cauet, H. Effect of pesticide exposure on human sperm characteristics, genome integrity, and methylation profile analysis. Environ. Sci. Pollut. Res. 2023, 30, 77560–77567. [Google Scholar] [CrossRef]
- Travaux et Implication de l’Anses sur les Perturbateurs Endocriniens. Anses—Agence Nationale de Sécurité Sanitaire de L’alimentation, de L’environnement et du Travail. 2019. Available online: https://www.anses.fr/fr/content/travaux-et-implication-de-lanses-sur-les-perturbateurs-endocriniens (accessed on 7 April 2023).
- USEPAO Overview of Endocrine Disruption. 2015. Available online: https://www.epa.gov/endocrine-disruption/overview-endocrine-disruption (accessed on 10 April 2023).
- US Environmental Protection Agency|US EPA. Available online: https://www.epa.gov/ (accessed on 25 February 2023).
- Akhtar, N.; Srivastava, M.K.; Raizada, R.B. Assessment of chlorpyrifos toxicity on certain organs in rat, Rattus norvegicus. J. Environ. Biol. 2009, 30, 1047–1053. [Google Scholar]
- Wołejko, E.; Łozowicka, B.; Jabłońska-Trypuć, A.; Pietruszyńska, M.; Wydro, U. Chlorpyrifos Occurrence and Toxicological Risk Assessment: A Review. Int. J. Environ. Res. Public Health 2022, 19, 12209. [Google Scholar] [CrossRef]
- Greer, J.B.; Magnuson, J.T.; Hester, K.; Giroux, M.; Pope, C.; Anderson, T.; Liu, J.; Dang, V.; Denslow, N.D.; Schlenk, D. Effects of Chlorpyrifos on Cholinesterase and Serine Lipase Activities and Lipid Metabolism in Brains of Rainbow Trout (Oncorhynchus mykiss). Toxicol. Sci. 2019, 172, 146–154. [Google Scholar] [CrossRef]
- Abou Diwan, M.; Lahimer, M.; Bach, V.; Gosselet, F.; Khorsi-Cauet, H.; Candela, P. Impact of Pesticide Residues on the Gut-Microbiota–Blood–Brain Barrier Axis: A Narrative Review. Int. J. Mol. Sci. 2023, 24, 6147. [Google Scholar] [CrossRef]
- Colović, M.B.; Krstić, D.Z.; Lazarević-Pašti, T.D.; Bondžić, A.M.; Vasić, V.M. Acetylcholinesterase inhibitors: Pharmacology and toxicology. Curr. Neuropharmacol. 2013, 11, 315–335. [Google Scholar] [CrossRef] [PubMed]
- Grandjean, P.; Barouki, R.; Bellinger, D.C.; Casteleyn, L.; Chadwick, L.H.; Cordier, S.; Etzel, R.A.; Gray, K.A.; Ha, E.-H.; Junien, C.; et al. Life-Long Implications of Developmental Exposure to Environmental Stressors: New Perspectives. Endocrinology 2015, 156, 3408–3415. [Google Scholar] [CrossRef] [PubMed]
- Djekkoun, N.; Depeint, F.; Guibourdenche, M.; El Khayat El Sabbouri, H.; Corona, A.; Rhazi, L.; Gay-Queheillard, J.; Rouabah, L.; Hamdad, F.; Bach, V.; et al. Chronic Perigestational Exposure to Chlorpyrifos Induces Perturbations in Gut Bacteria and Glucose and Lipid Markers in Female Rats and Their Offspring. Toxics 2022, 10, 138. [Google Scholar] [CrossRef] [PubMed]
- Mandy, M.; Nyirenda, M. Developmental Origins of Health and Disease: The relevance to developing nations. Int. Health 2018, 10, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Selevan, S.G.; Kimmel, C.A.; Mendola, P. Identifying critical windows of exposure for children’s health. Environ. Health Perspect. 2000, 108, 451–455. [Google Scholar]
- Safi-Stibler, S.; Gabory, A. Epigenetics and the Developmental Origins of Health and Disease: Parental environment signalling to the epigenome, critical time windows and sculpting the adult phenotype. Semin. Cell Dev. Biol. 2020, 97, 172–180. [Google Scholar] [CrossRef]
- Lacagnina, S. The Developmental Origins of Health and Disease (DOHaD). Am. J. Lifestyle Med. 2019, 14, 47–50. [Google Scholar] [CrossRef]
- Montjean, D.; Neyroud, A.-S.; Yefimova, M.G.; Benkhalifa, M.; Cabry, R.; Ravel, C. Impact of Endocrine Disruptors upon Non-Genetic Inheritance. Int. J. Mol. Sci. 2022, 23, 3350. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); Carrasco Cabrera, L.; Medina Pastor, P. The 2019 European Union report on pesticide residues in food. EFSA J. 2021, 19, e06491. [Google Scholar]
- Marques-Pinto, A.; Carvalho, D. Human infertility: Are endocrine disruptors to blame? Endocr. Connect. 2013, 2, R15–R29. [Google Scholar] [CrossRef] [PubMed]
- Padda, J.; Khalid, K.; Moosa, A.; Syam, M.; Kakani, V.; Imdad, U.; Ismail, D.; Cooper, A.C.; Jean-Charles, G. Role of Kisspeptin on Hypothalamic-Pituitary-Gonadal Pathology and Its Effect on Reproduction. Cureus 2021, 13, e17600. [Google Scholar] [CrossRef] [PubMed]
- Rønnekleiv, O.K.; Kelly, M.J. Kisspeptin Excitation of GnRH Neurons. Adv. Exp. Med. Biol. 2013, 784, 113–131. [Google Scholar] [PubMed]
- Prashar, V.; Arora, T.; Singh, R.; Sharma, A.; Parkash, J. Hypothalamic Kisspeptin Neurons: Integral Elements of the GnRH System. Reprod. Sci. 2023, 30, 802–822. [Google Scholar] [CrossRef]
- Han, S.-K.; Gottsch, M.L.; Lee, K.J.; Popa, S.M.; Smith, J.T.; Jakawich, S.K.; Clifton, D.K.; Steiner, R.A.; Herbison, A.E. Activation of Gonadotropin-Releasing Hormone Neurons by Kisspeptin as a Neuroendocrine Switch for the Onset of Puberty. J. Neurosci. 2005, 25, 11349–11356. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Xu, R.; Li, Y.; Zhou, Q.; Song, G.; Deng, Y.; Yan, Y. The effect of high-fat diet and exercise on KISS-1/GPR54 expression in testis of growing rats. Nutr. Metab. 2021, 18, 1. [Google Scholar] [CrossRef]
- Wang, J.-Y.; Hsu, M.-C.; Tseng, T.-H.; Wu, L.-S.; Yang, K.-T.; Chiu, C.-H. Kisspeptin expression in mouse Leydig cells correlates with age. J. Chin. Med. Assoc. 2015, 78, 249–257. [Google Scholar] [CrossRef][Green Version]
- Bhattacharya, M.; Babwah, A.V. Kisspeptin: Beyond the Brain. Endocrinology 2015, 156, 1218–1227. [Google Scholar] [CrossRef]
- Uenoyama, Y.; Pheng, V.; Tsukamura, H.; Maeda, K.I. The roles of Kisspeptin revisited: Inside and outside the hypothalamus. J. Reprod. Dev. 2016, 62, 537–545. [Google Scholar] [CrossRef]
- Rajin, T.; Patra, M.; Sheikh, P.A.; Singh, A.K.; Mishra, G.K.; Karikalan, M.; Singh, S.; Kumar, H.; Gaur, G.; Krishnaswamy, N. Expression of Kisspeptin and its receptor in different functional classes of ovarian follicle in the buffalo (Bubalus bubalis). Theriogenology 2022, 179, 87–96. [Google Scholar] [CrossRef]
- Tanyapanyachon, P.; Amelkina, O.; Chatdarong, K. The expression of Kisspeptin and its receptor in the domestic cat ovary and uterus in different stages of the ovarian cycle. Theriogenology 2018, 117, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Zeydabadi Nejad, S.; Ramezani Tehrani, F.; Zadeh-Vakili, A. The Role of Kisspeptin in Female Reproduction. Int. J. Endocrinol. Metab. 2017, 15, e44337. [Google Scholar] [CrossRef] [PubMed]
- Szeliga, A.; Meczekalski, B. Kisspeptin Modulation of Reproductive Function. Endocrines 2022, 3, 367–374. [Google Scholar] [CrossRef]
- Xie, Q.; Kang, Y.; Zhang, C.; Xie, Y.; Wang, C.; Liu, J.; Yu, C.; Zhao, H.; Huang, D. The Role of Kisspeptin in the Control of the Hypothalamic-Pituitary-Gonadal Axis and Reproduction. Front. Endocrinol. 2022, 13, 925206. [Google Scholar] [CrossRef]
- Seminara, S.B.; Messager, S.; Chatzidaki, E.E.; Thresher, R.R.; Acierno, J.S., Jr.; Shagoury, J.K.; Bo-Abbas, Y.; Kuohung, W.; Schwinof, K.M.; Hendrick, A.G.; et al. The GPR54 gene as a regulator of puberty. N. Engl. J. Med. 2003, 349, 1614–1627. [Google Scholar] [CrossRef]
- Abdelkareem, A.O.; Gebril, S.M.; AbdelHafez, F.F.; Terry, J.; Bedaiwy, M.A. Kisspeptin and Kisspeptin receptor immunoreactivity in euploid and aneuploid choriodecidual tissues of recurrent pregnancy losses. FS Sci. 2023, 4, 56–64. [Google Scholar] [CrossRef]
- Sengupta, A.; Chakrabarti, N.; Sridaran, R. Presence of Immunoreactive Gonadotropin Releasing Hormone (GnRH) and Its Receptor (GnRHR) in Rat Ovary During Pregnancy. Mol. Reprod. Dev. 2008, 75, 1031–1044. [Google Scholar] [CrossRef]
- Ciaramella, V.; Chianese, R.; Pariante, P.; Fasano, S.; Pierantoni, R.; Meccariello, R. Expression Analysis of Gnrh1 and Gnrhr1 in Spermatogenic Cells of Rat. Int. J. Endocrinol. 2015, 2015, e982726. [Google Scholar] [CrossRef]
- Palmon, A.; Ben Aroya, N.; Tel-Or, S.; Burstein, Y.; Fridkin, M.; Koch, Y. The gene for the neuropeptide gonadotropin-releasing hormone is expressed in the mammary gland of lactating rats. Proc. Natl. Acad. Sci. USA 1994, 91, 4994–4996. [Google Scholar] [CrossRef]
- Cheung, L.W.T.; Wong, A.S.T. Gonadotropin-releasing hormone: GnRH receptor signaling in extrapituitary tissues. FEBS J. 2008, 275, 5479–5495. [Google Scholar] [CrossRef]
- El Sabbouri, H.E.K.; Gay-Quéheillard, J.; Joumaa, W.H.; Delanaud, S.; Guibourdenche, M.; Darwiche, W.; Djekkoun, N.; Bach, V.; Ramadan, W. Does the perigestational exposure to chlorpyrifos and/or high-fat diet affect respiratory parameters and diaphragmatic muscle contractility in young rats? Food Chem. Toxicol. 2020, 140, 111322. [Google Scholar] [CrossRef] [PubMed]
- Elbetieha, A.; Da’as, S.I.; Khamas, W.; Darmani, H. Evaluation of the toxic potentials of cypermethrin pesticide on some reproductive and fertility parameters in the male rats. Arch. Environ. Contam. Toxicol. 2001, 41, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Dudás, J.; Dietl, W.; Romani, A.; Reinold, S.; Glueckert, R.; Schrott-Fischer, A.; Dejaco, D.; Chacko, L.J.; Tuertscher, R.; Schartinger, V.H.; et al. Nerve Growth Factor (NGF)—Receptor Survival Axis in Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2018, 19, 1771. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Sun, W.; Gao, F.; Ma, H.; Yuan, T.; Liu, Z.; Liu, H.; Hu, J.; Bai, J.; Zhang, X.; et al. Brain-Derived Estrogen Regulates Neurogenesis, Learning and Memory with Aging in Female Rats. Biology 2023, 12, 760. [Google Scholar] [CrossRef]
- Acker, C.I.; Nogueira, C.W. Chlorpyrifos acute exposure induces hyperglycemia and hyperlipidemia in rats. Chemosphere 2012, 89, 602–608. [Google Scholar] [CrossRef]
- Condette, C.J.; Khorsi-Cauet, H.; Morlière, P.; Zabijak, L.; Reygner, J.; Bach, V.; Gay-Quéheillard, J. Increased Gut Permeability and Bacterial Translocation after Chronic Chlorpyrifos Exposure in Rats. PLoS ONE 2014, 9, e102217. [Google Scholar] [CrossRef]
- Silva, M.H. Chlorpyrifos and Δ9 Tetrahydrocannabinol exposure and effects on parameters associated with the endocannabinoid system and risk factors for obesity. Curr. Res. Toxicol. 2021, 2, 296–308. [Google Scholar] [CrossRef]
- Tzoupis, H.; Nteli, A.; Androutsou, M.-E.; Tselios, T. Gonadotropin-Releasing Hormone and GnRH Receptor: Structure, Function and Drug Development. Curr. Med. Chem. 2020, 27, 6136–6158. [Google Scholar] [CrossRef]
- Miranda-Spooner, M.; Paccola, C.C.; Neves, F.M.O.; de Oliva, S.U.; Miraglia, S.M. Late reproductive analysis in rat male offspring exposed to nicotine during pregnancy and lactation. Andrology 2016, 4, 218–231. [Google Scholar] [CrossRef]
- Russell, L.D.; Ettlin, R.A.; Hikim, A.P.S.; Clegg, E.D. Histological and Histopathological Evaluation of the Testis. Int. J. Androl. 1993, 16, 83. [Google Scholar] [CrossRef]
- Omotoso, G.; Onanuga, I.; Ibrahim, R. Histological effects of permethrin insecticide on the testis of adult wistar rats. Ibnosina J. Med. Biomed. Sci. 2014, 6, 125–129. [Google Scholar] [CrossRef]
- Merviel, P.; Cabry, R.; Chardon, K.; Haraux, E.; Scheffler, F.; Mansouri, N.-B.; Devaux, A.; Chahine, H.; Bach, V.; Copin, H.; et al. Impact of oocytes with CLCG on ICSI outcomes and their potential relation to pesticide exposure. J. Ovarian Res. 2017, 10, 42. [Google Scholar] [CrossRef]
- Gupta, R.K.; Miller, K.P.; Babus, J.K.; Flaws, J.A. Methoxychlor Inhibits Growth and Induces Atresia of Antral Follicles through an Oxidative Stress Pathway. Toxicol. Sci. 2006, 93, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Corriero, A.; Zupa, R.; Mylonas, C.C.; Passantino, L. Atresia of ovarian follicles in fishes, and implications and uses in aquaculture and fisheries. J. Fish. Dis. 2021, 44, 1271–1291. [Google Scholar] [CrossRef] [PubMed]
- Gaytan, F.; Garcia-Galiano, D.; Dorfman, M.D.; Manfredi-Lozano, M.; Castellano, J.M.; Dissen, G.A.; Ojeda, S.R.; Tena-Sempere, M. Kisspeptin Receptor Haplo-insufficiency Causes Premature Ovarian Failure Despite Preserved Gonadotropin Secretion. Endocrinology 2014, 155, 3088–3097. [Google Scholar] [CrossRef]
- Meccariello, R. The Kisspeptin System in Male Reproduction. Endocrines 2022, 3, 168–174. [Google Scholar] [CrossRef]
- Izadi, V.; Saraf-Bank, S.; Azadbakht, L. Dietary intakes and leptin concentrations. ARYA Atheroscler. 2014, 10, 266–272. [Google Scholar]
- Childs, G.V.; Odle, A.K.; MacNicol, M.C.; MacNicol, A.M. The Importance of Leptin to Reproduction. Endocrinology 2020, 162, bqaa204. [Google Scholar] [CrossRef]
- Hiller-Sturmhöfel, S.; Bartke, A. The Endocrine System. Alcohol. Health Res. World 1998, 22, 153–164. [Google Scholar]
- Shankaran, H.; Wiley, H.S.; Resat, H. Receptor downregulation and desensitization enhance the information processing ability of signalling receptors. BMC Syst. Biol. 2007, 1, 48. [Google Scholar] [CrossRef]
- Mueller, J.K.; Heger, S. Endocrine disrupting chemicals affect the Gonadotropin releasing hormone neuronal network. Reprod. Toxicol. 2014, 44, 73–84. [Google Scholar] [CrossRef]
- Roepke, T.A.; Sadlier, N.C. Reproductive Toxicology: Impact of endocrine disruptors on neurons expressing GnRH or Kisspeptin and pituitary gonadotropins. Reproduction 2021, 162, F131–F145. [Google Scholar] [CrossRef]
- Dickerson, S.M.; Cunningham, S.L.; Gore, A.C. Prenatal PCBs disrupt early neuroendocrine development of the rat hypothalamus. Toxicol. Appl. Pharmacol. 2011, 252, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Feng, J.; Liang, C.; Song, G.; Yan, Y. Effects of high-fat diet and treadmill running on the hypothalamic Kiss-1-GPR54 signaling pathway in male growing rats. Hormones 2022, 21, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Li, H.; Zhang, D.; Lv, Y.; Lin, X.; Zhang, H. Expression of Kisspeptin/kiss1r System is Down-regulated in the Hypothalamic Arcuate Nucleus of Pubertal Male Rats with High-fat-diet. J. Reprod. Contracept. 2014, 25, 1–11. [Google Scholar]
- Alabduljabbar, S.; Zaidan, S.A.; Lakshmanan, A.P.; Terranegra, A. Personalized Nutrition Approach in Pregnancy and Early Life to Tackle Childhood and Adult Non-Communicable Diseases. Life 2021, 11, 467. [Google Scholar] [CrossRef]
- Yehuda, R.; Lehrner, A. Intergenerational transmission of trauma effects: Putative role of epigenetic mechanisms. World Psychiatry 2018, 17, 243–257. [Google Scholar] [CrossRef]
- Lee, E.B.; Dilower, I.; Marsh, C.A.; Wolfe, M.W.; Masumi, S.; Upadhyaya, S.; Rumi, M.A.K. Sexual Dimorphism in Kisspeptin Signaling. Cells 2022, 11, 1146. [Google Scholar] [CrossRef]
- Dugershaw, B.B.; Aengenheister, L.; Hansen, S.S.K.; Hougaard, K.S.; Buerki-Thurnherr, T. Recent insights on indirect mechanisms in developmental toxicity of nanomaterials. Part. Fibre Toxicol. 2020, 17, 31. [Google Scholar] [CrossRef]
- Sommer, M.; Mmari, K. Addressing Structural and Environmental Factors for Adolescent Sexual and Reproductive Health in Low- and Middle-Income Countries. Am. J. Public Health 2015, 105, 1973–1981. [Google Scholar] [CrossRef]
- Chargui, I.; Grissa, I.; Bensassi, F.; Hrira, M.Y.; Haouem, S.; Haouas, Z.; Bencheikh, H. Oxidative Stress, Biochemical and Histopathological Alterations in the Liver and Kidney of Female Rats Exposed to Low Doses of Deltamethrin (DM): A Molecular Assessment. Biomed. Environ. Sci. 2012, 25, 672–683. [Google Scholar] [PubMed]
- El-Din, M.A.E.-D.S.; Ghareeb, A.E.-W.E.; El-Garawani, I.M.; El-Rahman, H.A.A. Induction of apoptosis, oxidative stress, hormonal, and histological alterations in the reproductive system of thiamethoxam-exposed female rats. Environ. Sci. Pollut. Res. 2023, 30, 77917–77930. [Google Scholar] [CrossRef] [PubMed]
- Watermann, B.; Grote, K.; Gnass, K.; Kolodzey, H.; Thomsen, A.; Appel, K.; Candia-Carnevali, D.; Schulte-Oehlmann, U. Histological alterations in ovaries of pubertal female rats induced by triphenyltin. Exp. Toxicol. Pathol. 2008, 60, 313–321. [Google Scholar] [CrossRef]
- Grewal, K.K.; Sandhu, G.S.; Kaur, R.; Brar, R.S.; Sandhu, H.S. Toxic Impacts of Cypermethrin on Behavior and Histology of Certain Tissues of Albino Rats. Toxicol. Int. 2010, 17, 94–98. [Google Scholar]
- Lukowicz, C.; Ellero-Simatos, S.; Régnier, M.; Polizzi, A.; Lasserre, F.; Montagner, A.; Lippi, Y.; Jamin, E.L.; Martin, J.-F.; Naylies, C.; et al. Metabolic Effects of a Chronic Dietary Exposure to a Low-Dose Pesticide Cocktail in Mice: Sexual Dimorphism and Role of the Constitutive Androstane Receptor. Environ. Health Perspect. 2018, 126, 067007. [Google Scholar] [CrossRef] [PubMed]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
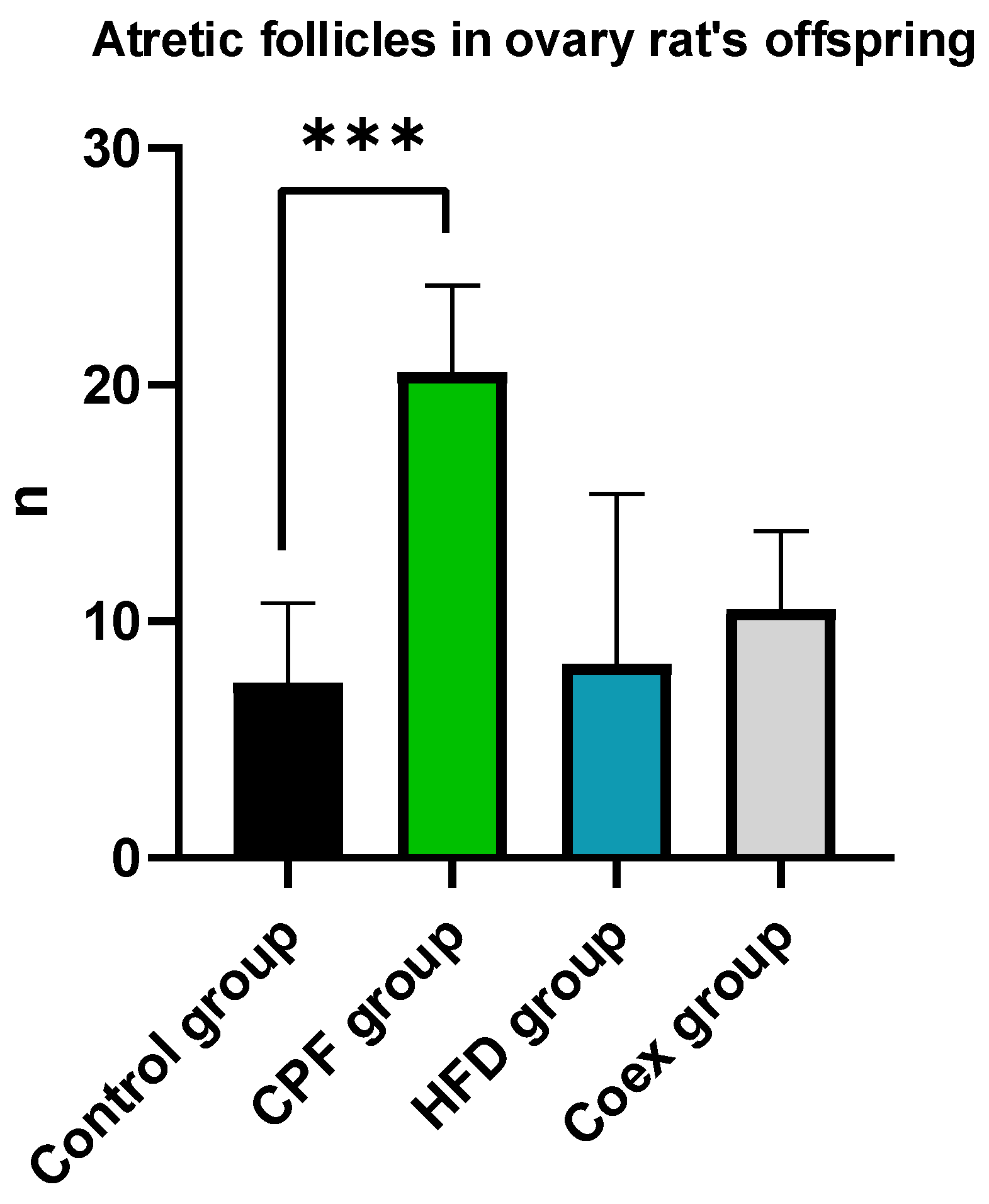{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean ± Sd (µm2) | F Value | p-Value | |
|---|---|---|---|
| Control group | 1107.445 ± 781.704 | ||
| CPF group | 1255.563 ± 636.659 | 0.2 | 0.6 |
| HFD group | 1360.484 ± 542.038 | 0.08 | 0.7 |
| Coexp group | 1082.892 ± 442.743 | 2.4 | 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lahimer, M.; Djekkoun, N.; Tricotteaux-Zarqaoui, S.; Corona, A.; Lafosse, I.; Ali, H.B.; Ajina, M.; Bach, V.; Benkhalifa, M.; Khorsi-Cauet, H. Impact of Perinatal Coexposure to Chlorpyrifos and a High-Fat Diet on Kisspeptin and GnRHR Presence and Reproductive Organs. Toxics 2023, 11, 789. https://doi.org/10.3390/toxics11090789
Lahimer M, Djekkoun N, Tricotteaux-Zarqaoui S, Corona A, Lafosse I, Ali HB, Ajina M, Bach V, Benkhalifa M, Khorsi-Cauet H. Impact of Perinatal Coexposure to Chlorpyrifos and a High-Fat Diet on Kisspeptin and GnRHR Presence and Reproductive Organs. Toxics. 2023; 11(9):789. https://doi.org/10.3390/toxics11090789
Chicago/Turabian StyleLahimer, Marwa, Narimane Djekkoun, Sophian Tricotteaux-Zarqaoui, Aurélie Corona, Isabelle Lafosse, Habib Ben Ali, Mounir Ajina, Véronique Bach, Moncef Benkhalifa, and Hafida Khorsi-Cauet. 2023. "Impact of Perinatal Coexposure to Chlorpyrifos and a High-Fat Diet on Kisspeptin and GnRHR Presence and Reproductive Organs" Toxics 11, no. 9: 789. https://doi.org/10.3390/toxics11090789
APA StyleLahimer, M., Djekkoun, N., Tricotteaux-Zarqaoui, S., Corona, A., Lafosse, I., Ali, H. B., Ajina, M., Bach, V., Benkhalifa, M., & Khorsi-Cauet, H. (2023). Impact of Perinatal Coexposure to Chlorpyrifos and a High-Fat Diet on Kisspeptin and GnRHR Presence and Reproductive Organs. Toxics, 11(9), 789. https://doi.org/10.3390/toxics11090789