


Mathematical Estimation of Endogenous Proline as a Bioindicator to Regulate the Stress of Trivalent Chromium on Rice Plants Grown in Different Nitrogenous Conditions


Abstract
:1. Introduction
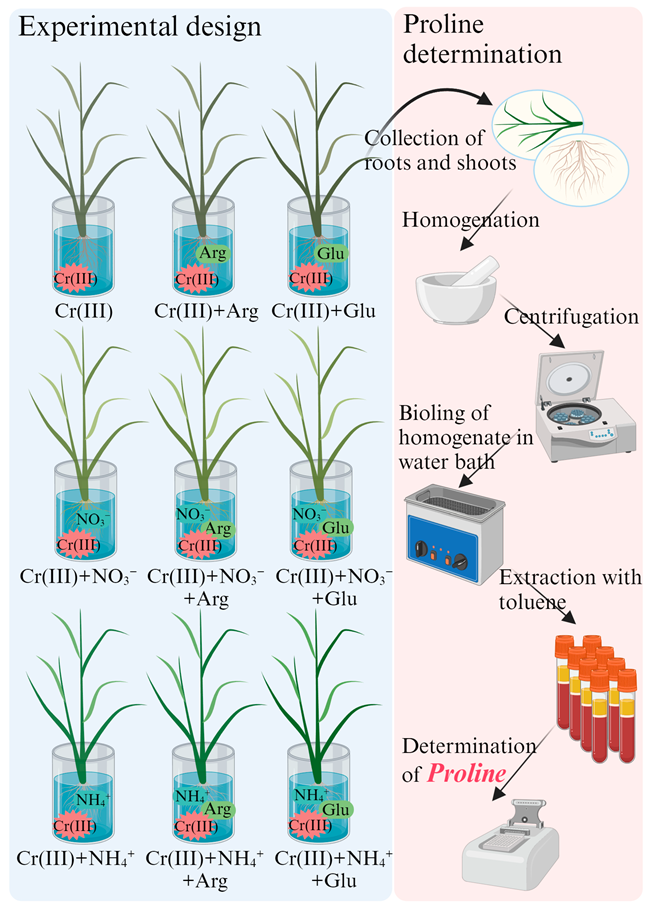
2. Methods and Materials
2.1. Rice Seedlings and Cr Treatment
2.2. Measurement of Relative Growth Rate
2.3. Measurement of Pro Content in Rice Seedlings
2.4. Modeling the “Mass Balance Matrix”
3. Results
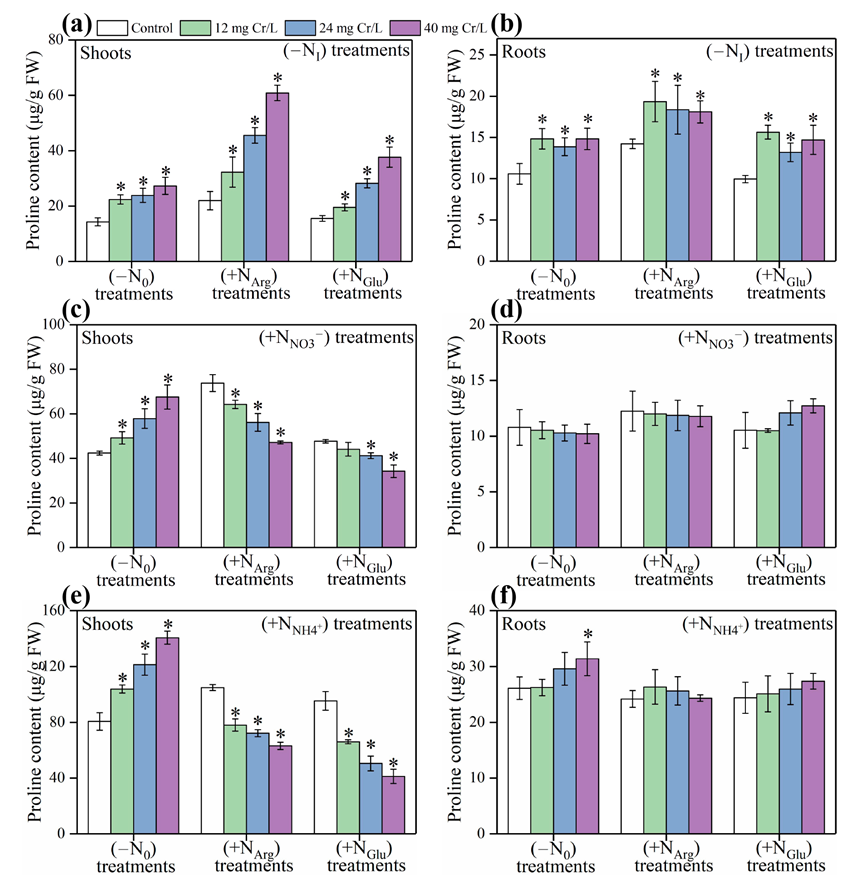
3.1. Pro Content in Rice Tissues under ‘Cr(III)+(−NI)’ Treatments
3.2. Pro Content in Rice Tissues under ‘Cr(III)+(+NNO3−)’ Treatments
3.3. Pro Content in Rice Tissues under ‘Cr(III)+(+NNH4+)’ Treatments
3.4. The Contribution of Organic and Inorganic N Application to Pro Content in Rice Seedlings
3.4.1. Pro Content in Shoots of Cr(III)-Treated Rice Seedlings
3.4.2. Pro Content in Roots of Cr(III)-Treated Rice Seedlings
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pan, X.; Ullah, A.; Feng, Y.X.; Tian, P.; Yu, X.Z. Proline-mediated activation of glyoxalase II improve methylglyoxal detoxification in Oryza sativa L. under chromium injury: Clarification via vector analysis of enzymatic activities and gene expression. Plant Physiol. Biochem. 2023, 201, 107867. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.X.; Yu, X.Z.; Mo, C.H.; Lu, C.J. Regulation network of sucrose metabolism in response to trivalent and hexavalent chromium in Oryza sativa. J. Agric. Food Chem. 2019, 67, 9738–9748. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.J.; Feng, Y.X.; Li, Y.H.; Lin, Y.J.; Yu, X.Z. Unraveling genes promoting ROS metabolism in subcellular organelles of Oryza sativa in response to trivalent and hexavalent chromium. Sci. Total Environ. 2020, 744, 140951. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Feng, Y.X.; Lin, Y.J.; Yu, X.Z. Mathematical quantification of interactive complexity of transcription factors involved in proline-mediated regulative strategies in Oryza sativa under chromium stress. Plant Physiol. Biochem. 2022, 182, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; Lopes, J.; Sousa, B.; Soares, C.; Valente, I.M.; Rodrigues, J.A.; Fidalgo, F.; Teixeira, J. Cr (VI)-induced oxidative damage impairs ammonia assimilation into organic forms in Solanum lycopersicum L. Plant Stress 2021, 2, 100034. [Google Scholar] [CrossRef]
- Li, C.Z.; Yang, L.; Lin, Y.J.; Zhang, H.; Rad, S.; Yu, X.Z. Assimilation of exogenous cyanide cross talk in Oryza sativa L. to the key nodes in nitrogen metabolism. Ecotoxicology 2020, 29, 1552–1564. [Google Scholar] [CrossRef]
- Chen, M.; Zhu, K.; Xie, J.; Liu, J.; Qiao, Z.; Tan, P.; Peng, F. Ammonium-nitrate mixtures dominated by NH4+-N promote the growth of pecan (Carya illinoinensis) through enhanced N uptake and assimilation. Front. Plant Sci. 2023, 14, 1186818. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Xu, M.Y.; Wang, S.Z.; Wu, L.D.; Sun, S.Y.; Su, T. Effects of exogenous L-Glutamine as a sole nitrogen source on physiological characteristics and nitrogen use efficiency of poplar. Plant Physiol. Biochem. 2022, 172, 1–13. [Google Scholar] [CrossRef]
- Hosseinifard, M.; Stefaniak, S.; Ghorbani Javid, M.; Soltani, E.; Wojtyla, Ł.; Garnczarska, M. Contribution of exogenous proline to abiotic stresses tolerance in plants: A review. Int. J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef]
- Meena, M.; Divyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Upadhyay, R.S. Regulation of L-proline biosynthesis, signal transduction, transport, accumulation and its vital role in plants during variable environmental conditions. Heliyon 2019, 5, e02952. [Google Scholar] [CrossRef]
- Spormann, S.; Nadais, P.; Sousa, F.; Pinto, M.; Martins, M.; Sousa, B.; Soares, C. Accumulation of proline in plants under contaminated soils-are we on the same page? Antioxidants 2023, 12, 666. [Google Scholar] [CrossRef]
- Li, Y.H.; Tian, P.; Li, C.Z.; Yu, X.Z. Elucidating comportment of the glutamate and ornithine pathway on proline accumulation in rice under different nitrogenous nutrition. Int. J. Environ. Sci. Technol. 2022, 19, 2993–3000. [Google Scholar] [CrossRef]
- Luo, L.; Zhang, Y.; Xu, G. How does nitrogen shape plant architecture? J. Exp. Bot. 2020, 71, 4415–4427. [Google Scholar] [CrossRef] [PubMed]
- Sangwan, P.; Kumar, V.; Joshi, U.N. Effect of chromium (VI) toxicity on enzymes of nitrogen metabolism in clusterbean (Cyamopsis tetragonoloba L.). Enzym. Res. 2014, 2014, 784036. [Google Scholar] [CrossRef]
- Huo, K.; Shangguan, X.; Xia, Y.; Shen, Z.; Chen, C. Excess copper inhibits the growth of rice seedlings by decreasing uptake of nitrate. Ecotoxicol. Environ. Saf. 2020, 190, 110105. [Google Scholar] [CrossRef]
- Chai, M.; Li, R.; Shen, X.; Tam, N.F.Y.; Zan, Q.; Li, R. Does ammonium nitrogen affect accumulation, subcellular distribution and chemical forms of cadmium in Kandelia obovata? Ecotoxicol. Environ. Saf. 2018, 162, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.Z.; Lin, Y.J.; Fan, W.J.; Lu, M.R. The role of exogenous proline in amelioration of lipid peroxidation in rice seedlings exposed to Cr (VI). Int. Biodeter. Biodegrad. 2017, 123, 106–112. [Google Scholar] [CrossRef]
- López-Bucio, J.S.; Ravelo-Ortega, G.; López-Bucio, J. Chromium in plant growth and development: Toxicity, tolerance and hormesis. Environ. Pollut. 2022, 312, 120084. [Google Scholar] [CrossRef] [PubMed]
- Danyal, Y.; Mahmood, K.; Ullah, S.; Rahim, A.; Raheem, G.; Khan, A.H.; Ullah, A. Phytoremediation of industrial effluents assisted by plant growth promoting bacteria. Environ. Sci. Pollut. Res. 2023, 30, 5296–5311. [Google Scholar] [CrossRef]
- Choudhary, S.P.; Kanwar, M.; Bhardwaj, R.; Gupta, B.D.; Gupta, R.K. Epibrassinolide ameliorates Cr (VI) stress via influencing the levels of indole-3-acetic acid, abscisic acid, polyamines and antioxidant system of radish seedlings. Chemosphere 2011, 84, 592–600. [Google Scholar] [CrossRef]
- Ullah, A.; Heng, S.; Munis, M.F.H.; Fahad, S.; Yang, X. Phytoremediation of heavy metals assisted by plant growth promoting (PGP) bacteria: A review. Environ. Exp. Bot. 2015, 117, 28–40. [Google Scholar] [CrossRef]
- Lin, Y.J.; Feng, Y.X.; Li, Y.H.; Yu, G.; Yu, X.Z. Fuzzy synthetic evaluation of the impact of plant growth regulators on the root phenotype traits of rice seedlings under thiocyanate stress. Plant Physiol. Biochem. 2021, 158, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Stein, H.; Honig, A.; Miller, G.; Erster, O.; Eilenberg, H.; Csonka, L.N.; Sazbados, L.; Koncz, C.; Zilberstein, A. Elevation of free proline and proline-rich protein levels by simultaneous manipulations of proline biosynthesis and degradation in plants. Plant Sci. 2011, 181, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Wu, L.; Wu, M.; Zhu, C.; Jin, Q.; Zhang, J. Abscisic acid mediated proline biosynthesis and antioxidant ability in roots of two different rice genotypes under hypoxic stress. BMC Plant Biol. 2020, 20, 198. [Google Scholar] [CrossRef] [PubMed]
- Cambui, C.A.; Svennerstam, H.; Gruffman, L.; Nordin, A.; Ganeteg, U.; Näsholm, T. Patterns of plant biomass partitioning depend on nitrogen source. PLoS ONE 2011, 6, e19211. [Google Scholar] [CrossRef] [PubMed]
- Carlisle, E.; Myers, S.; Raboy, V.; Bloom, A. The effects of inorganic nitrogen form and CO2 concentration on wheat yield and nutrient accumulation and distribution. Front. Plant Sci. 2012, 3, 195. [Google Scholar] [CrossRef]
- Trovato, M.; Funck, D.; Forlani, G.; Okumoto, S.; Amir, R. Amino acids in plants: Regulation and functions in development and stress defense. Front. Plant Sci. 2021, 12, 772810. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Winter, G.; Todd, C.D.; Trovato, M.; Forlani, G.; Funck, D. Physiological implications of arginine metabolism in plants. Front. Plant Sci. 2015, 6, 534. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, Y.; Zhang, Z.; Liu, X.; Li, C.; Ma, F. Arginine Increases Tolerance to Nitrogen Deficiency in Malus hupehensis via Alterations in Photosynthetic Capacity and Amino Acids Metabolism. Front. Plant Sci. 2022, 12, 772086. [Google Scholar] [CrossRef] [PubMed]
- Mondal, M.H.; Begum, W.; Nasrollahzadeh, M.; Ghorbannezhad, F.; Antoniadis, V.; Levizou, E.; Saha, B. A comprehensive review on chromium chemistry along with detection, speciation, extraction and remediation of hexavalent chromium in contemporary science and technology. Vietnam J. Chem. 2021, 59, 711–732. [Google Scholar]
- Gaur, V.S.; Singh, U.S.; Gupta, A.K.; Kumar, A. Understanding the differential nitrogen sensing mechanism in rice genotypes through expression analysis of high and low affinity ammonium transporter genes. Mol. Biol. Rep. 2012, 39, 2233–2241. [Google Scholar] [CrossRef] [PubMed]
- Thornton, B.; Robinson, D. Uptake and assimilation of nitrogen from solutions containing multiple N sources. Plant Cell Environ. 2005, 28, 813–821. [Google Scholar] [CrossRef]
- Cao, X.C.; Zhong, C.; Zhu, C.Q.; Zhu, L.F.; Zhang, J.H.; Wu, L.H.; Jin, Q.Y. Ammonium uptake and metabolism alleviate PEG-induced water stress in rice seedlings. Plant Physiol. Biochem. 2018, 132, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, N.; Umar, S.; Khan, N.A. Nitrogen availability regulates proline and ethylene production and alleviates salinity stress in mustard (Brassica juncea). J. Plant Physiol. 2015, 178, 84–91. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Treatments | Organic and Inorganic N Treatments Alone | Organic and Inorganic N Treatments in Combination | ||
|---|---|---|---|---|
| Shoot | Root | Shoot | Root | |
| Control | NNH4+ > NNO3− > NArg > NGlu | NArg > NNH4+ > NGlu > NNO3− | NNH4+ + NArg > NNH4+ + NGlu > NNO3− + NArg > NNO3− + NGlu | NNH4+ + NArg ≈ NNH4+ + NGlu > NNO3− + NArg ≈ NNO3− + NGlu |
| 12 mg Cr/L | NNH4+ > NNO3− > NArg > NGlu | NArg > NNH4+ > NGlu > NNO3− | NNH4+ + NArg > NNH4+ + NGlu > NNO3− + NArg > NNO3− + NGlu | NNH4+ + NArg ≈ NNH4+ + NGlu > NNO3− + NArg ≈ NNO3− + NGlu |
| 24 mg Cr/L | NNH4+ > NNO3− > NArg > NGlu | NArg > NNH4+ > NGlu > NNO3− | NNH4+ + NArg > NNO3− + NArg > NNH4+ + NGlu > NNO3− + NGlu | NNH4+ + NArg ≈ NNH4+ + NGlu > NNO3− + NArg ≈ NNO3− + NGlu |
| 40 mg Cr/L | NNH4+ > NNO3− > NArg > NGlu | NArg > NNH4+ > NGlu > NNO3− | NNH4+ + NArg > NNO3− + NArg > NNH4+ + NGlu > NNO3− + NGlu | NNH4+ + NArg ≈ NNH4+ + NGlu > NNO3− + NArg ≈ NNO3− + NGlu |
| In summary | NNH4+ > NNO3− > NArg > NGlu | NArg > NNH4+ > NGlu > NNO3− | 0 and 12 mg Cr/L treatment: NNH4+ + NArg > NNH4+ + NGlu > NNO3− + NArg > NNO3− + NGlu 24 and 40 mg Cr/L treatment: NNH4+ + NArg > NNO3− + NArg > NNH4+ + NGlu > NNO3− + NGlu | NNH4+ + NArg ≈ NNH4+ + NGlu > NNO3− + NArg ≈ NNO3− + NGlu |
| Treatments | (12) | (13) | (14) | (15) | |
|---|---|---|---|---|---|
| Shoot | Control | ||||
| 12 mgCr/L | |||||
| 24 mgCr/L | |||||
| 40 mgCr/L |
| Treatments | (16) | (17) | (18) | (19) | |
|---|---|---|---|---|---|
| Root | Control | ||||
| 12 mg Cr/L | |||||
| 24 mg Cr/L | |||||
| 40 mg Cr/L |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Feng, Y.; Tian, P.; Yu, X. Mathematical Estimation of Endogenous Proline as a Bioindicator to Regulate the Stress of Trivalent Chromium on Rice Plants Grown in Different Nitrogenous Conditions. Toxics 2023, 11, 803. https://doi.org/10.3390/toxics11100803
Li C, Feng Y, Tian P, Yu X. Mathematical Estimation of Endogenous Proline as a Bioindicator to Regulate the Stress of Trivalent Chromium on Rice Plants Grown in Different Nitrogenous Conditions. Toxics. 2023; 11(10):803. https://doi.org/10.3390/toxics11100803
Chicago/Turabian StyleLi, Chengzhi, Yuxi Feng, Peng Tian, and Xiaozhang Yu. 2023. "Mathematical Estimation of Endogenous Proline as a Bioindicator to Regulate the Stress of Trivalent Chromium on Rice Plants Grown in Different Nitrogenous Conditions" Toxics 11, no. 10: 803. https://doi.org/10.3390/toxics11100803
APA StyleLi, C., Feng, Y., Tian, P., & Yu, X. (2023). Mathematical Estimation of Endogenous Proline as a Bioindicator to Regulate the Stress of Trivalent Chromium on Rice Plants Grown in Different Nitrogenous Conditions. Toxics, 11(10), 803. https://doi.org/10.3390/toxics11100803