
Harmful Effects of Pyraclostrobin on the Fat Body and Pericardial Cells of Foragers of Africanized Honey Bee


 ,
,  ,
, 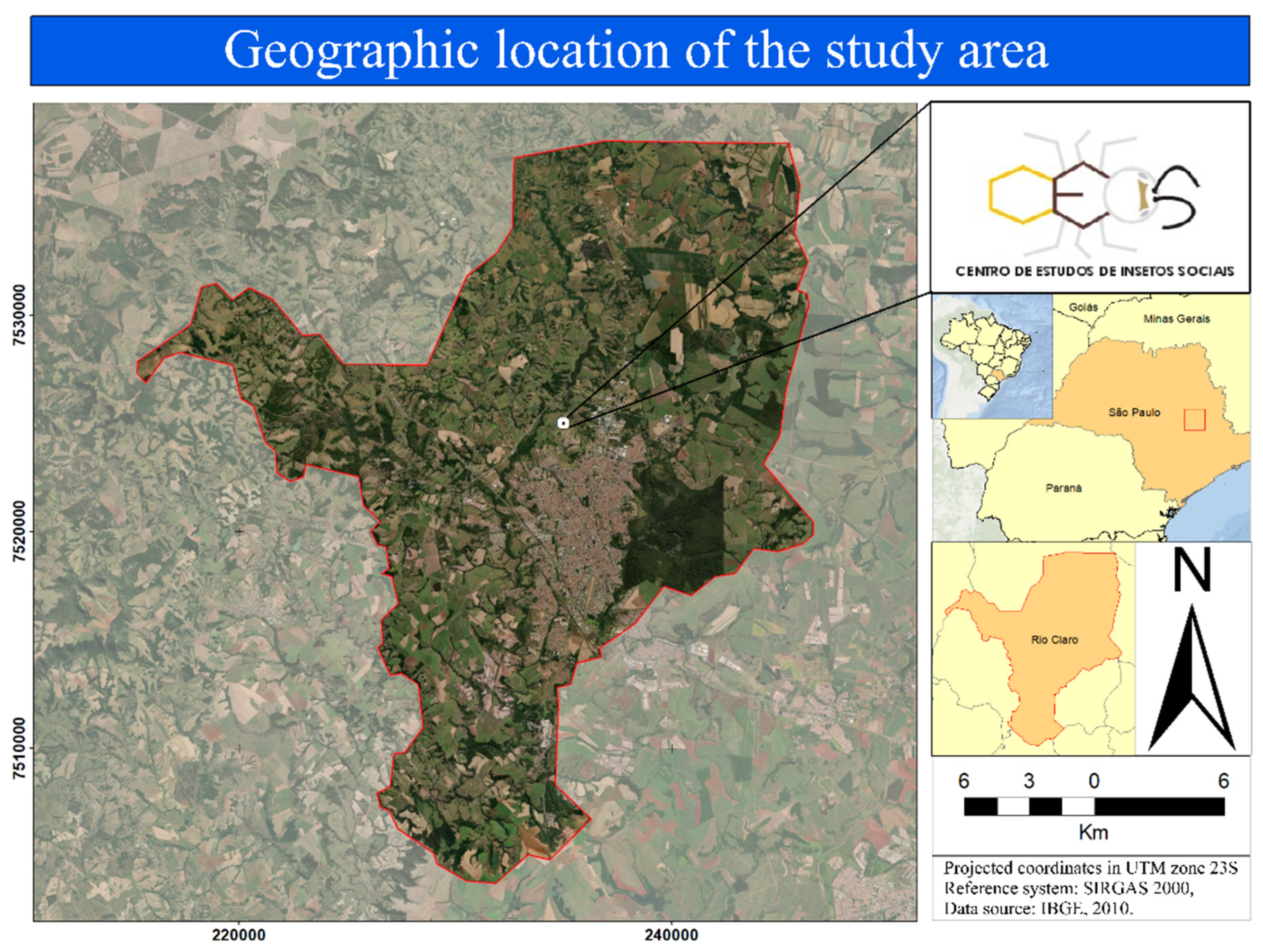{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Honey Bee Collection
2.2. Chemical
2.3. Experimental Design-Toxicological Bioassays
2.4. Histological Processing of the Fat Body
2.5. Qualitative Analysis of Fat Body and Pericardial Cells
2.6. Morphometry of Oenocytes and Pericardial Cells
2.7. Statistical Analysis
3. Results
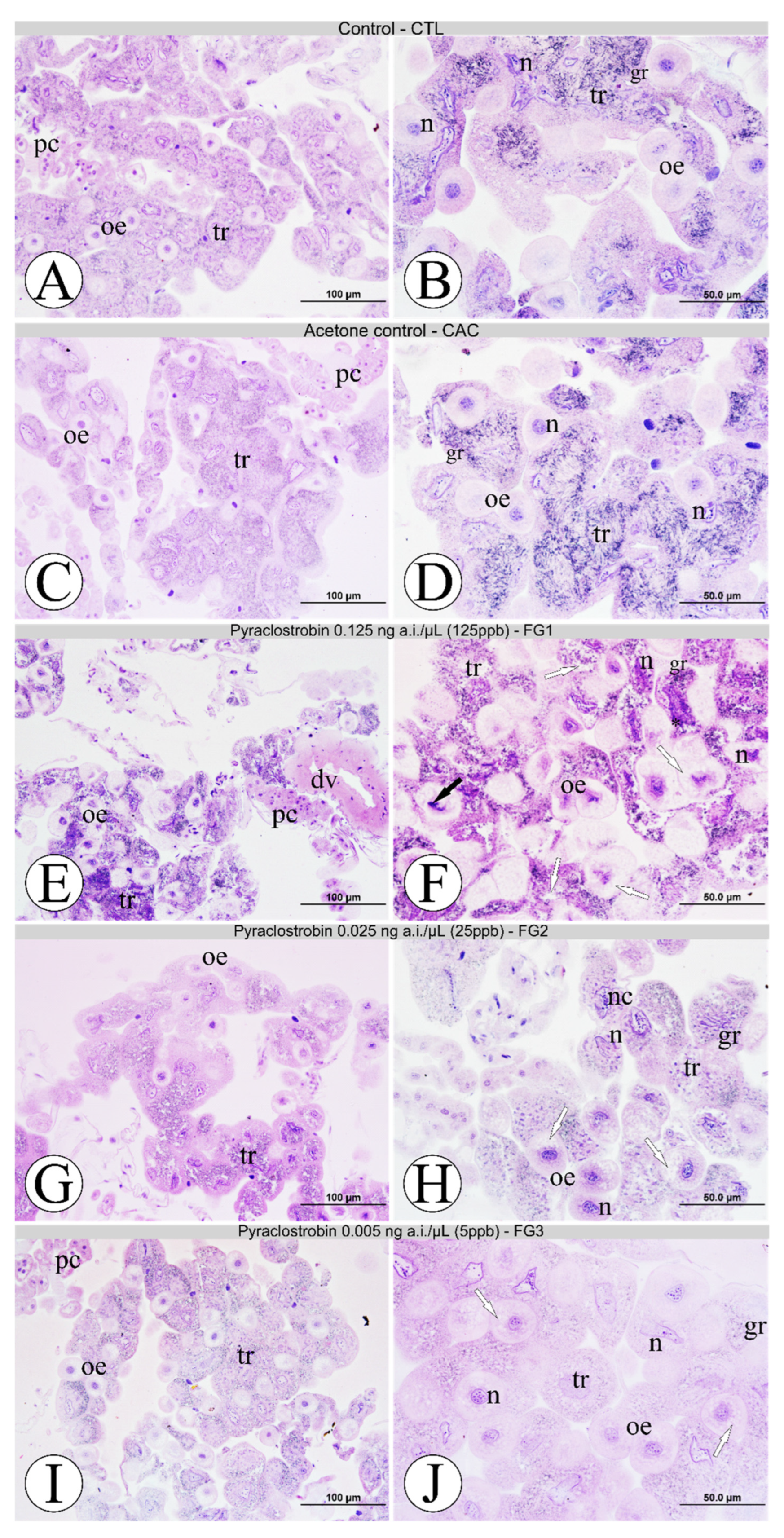
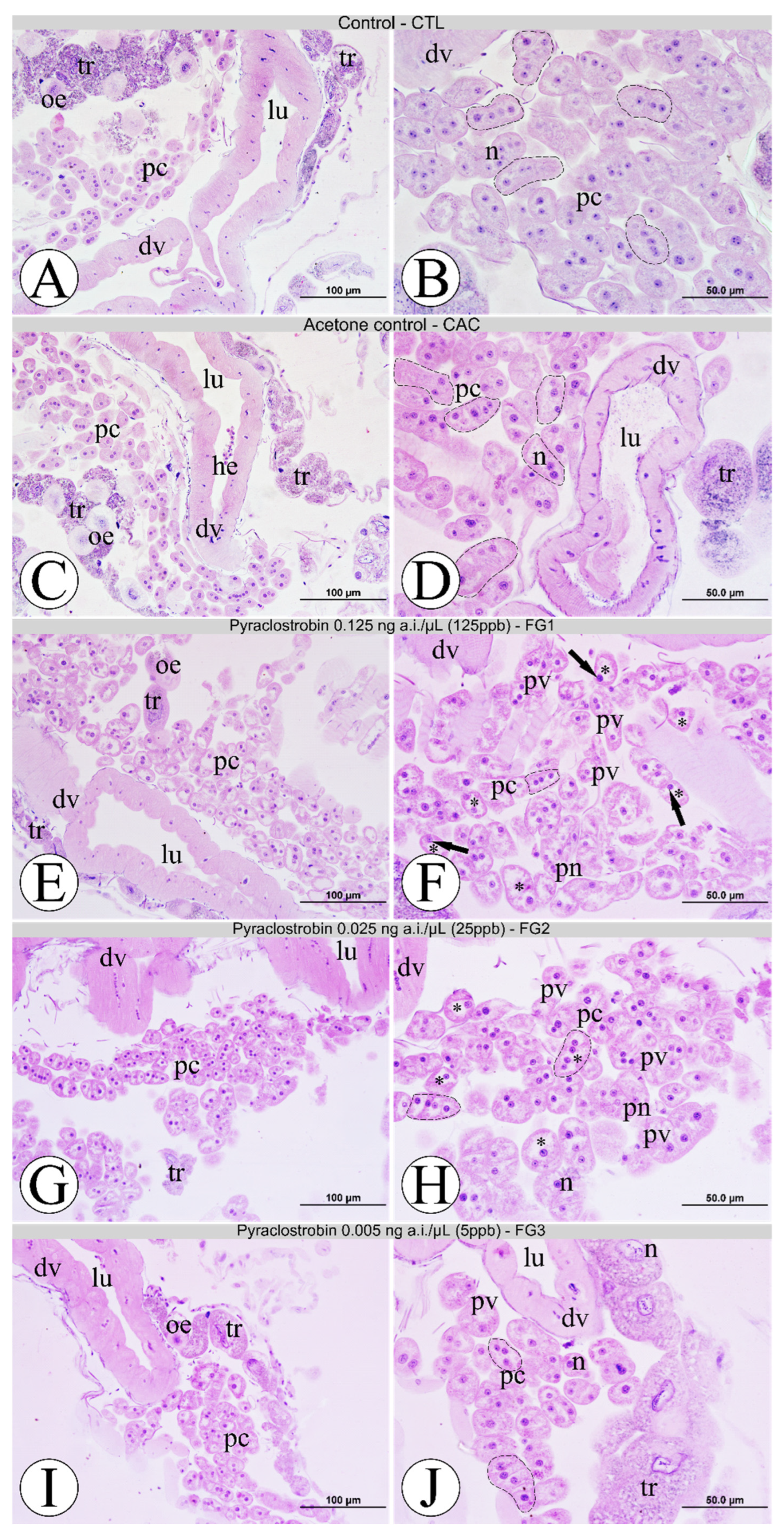
3.1. Morphological Analyses of Fat Body Cells
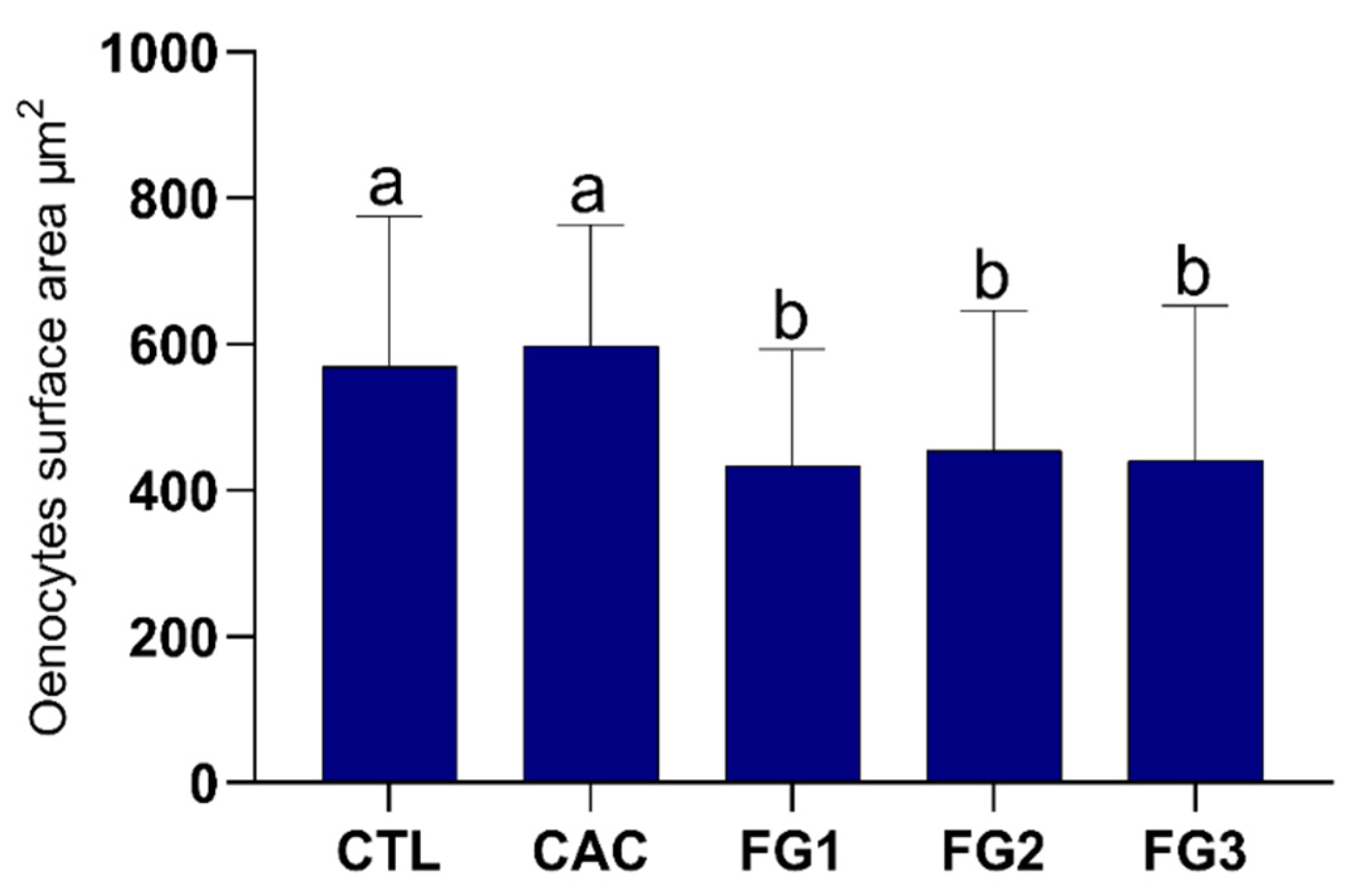
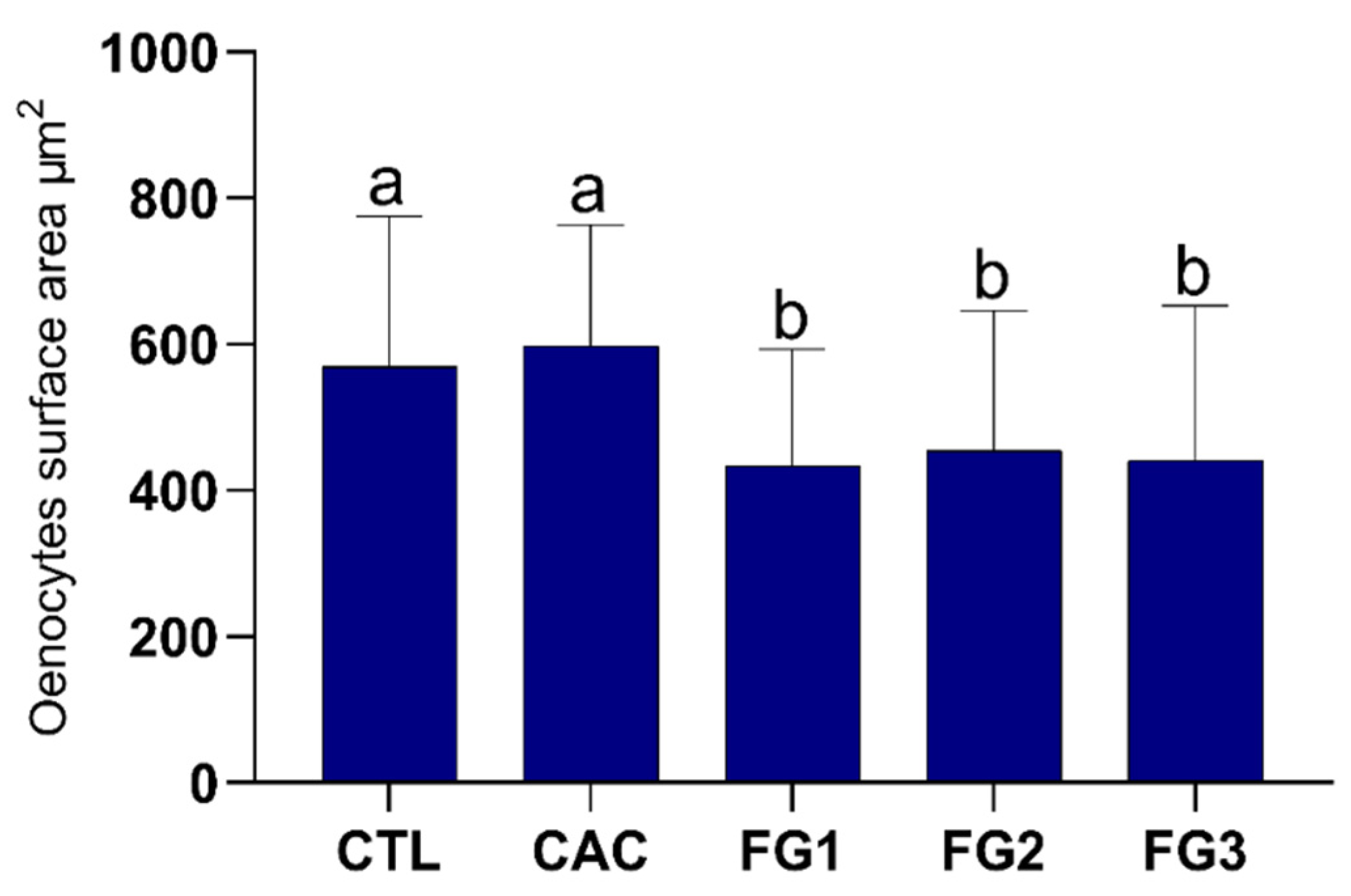
3.2. Morphometric Analyses of Oenocytes
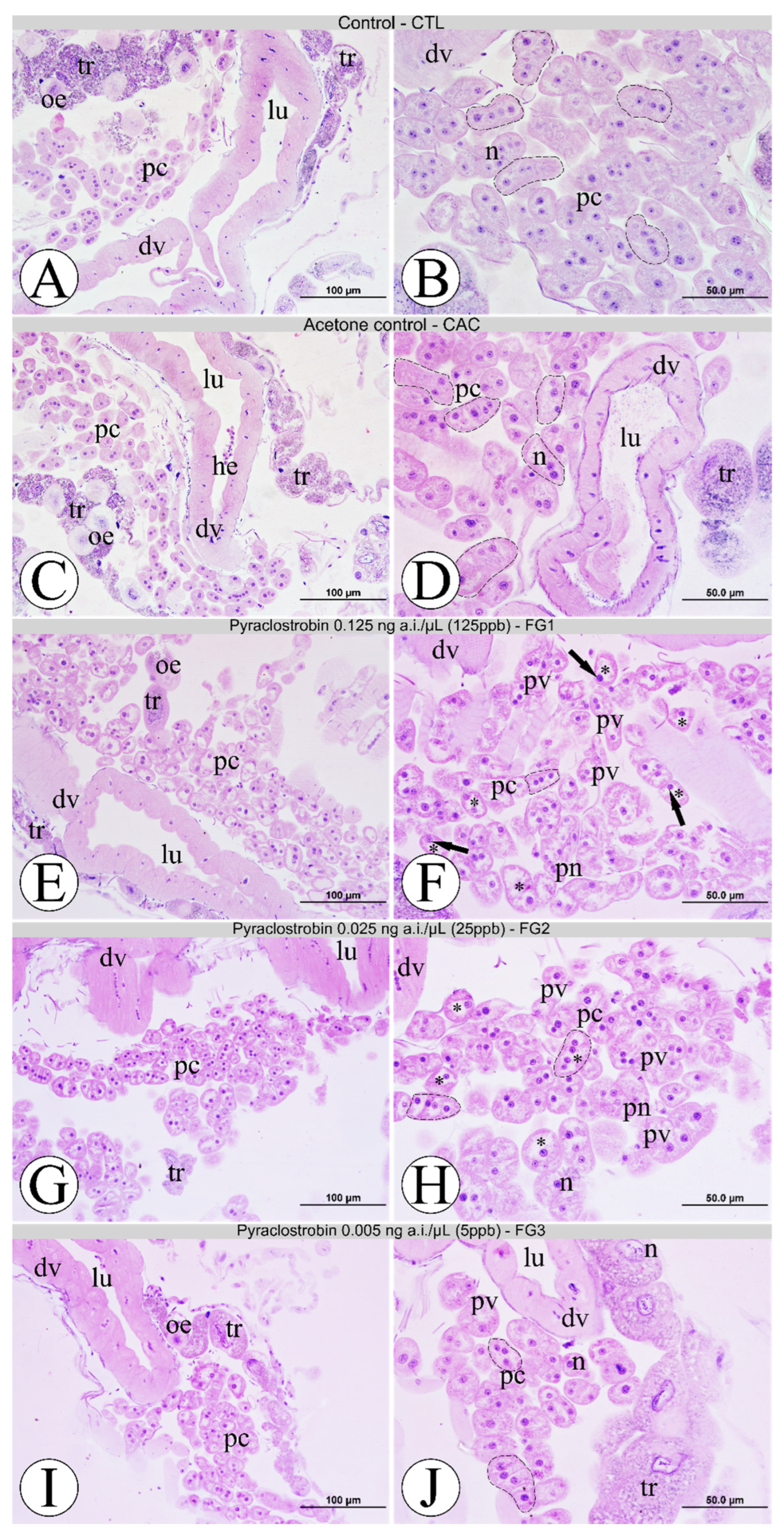
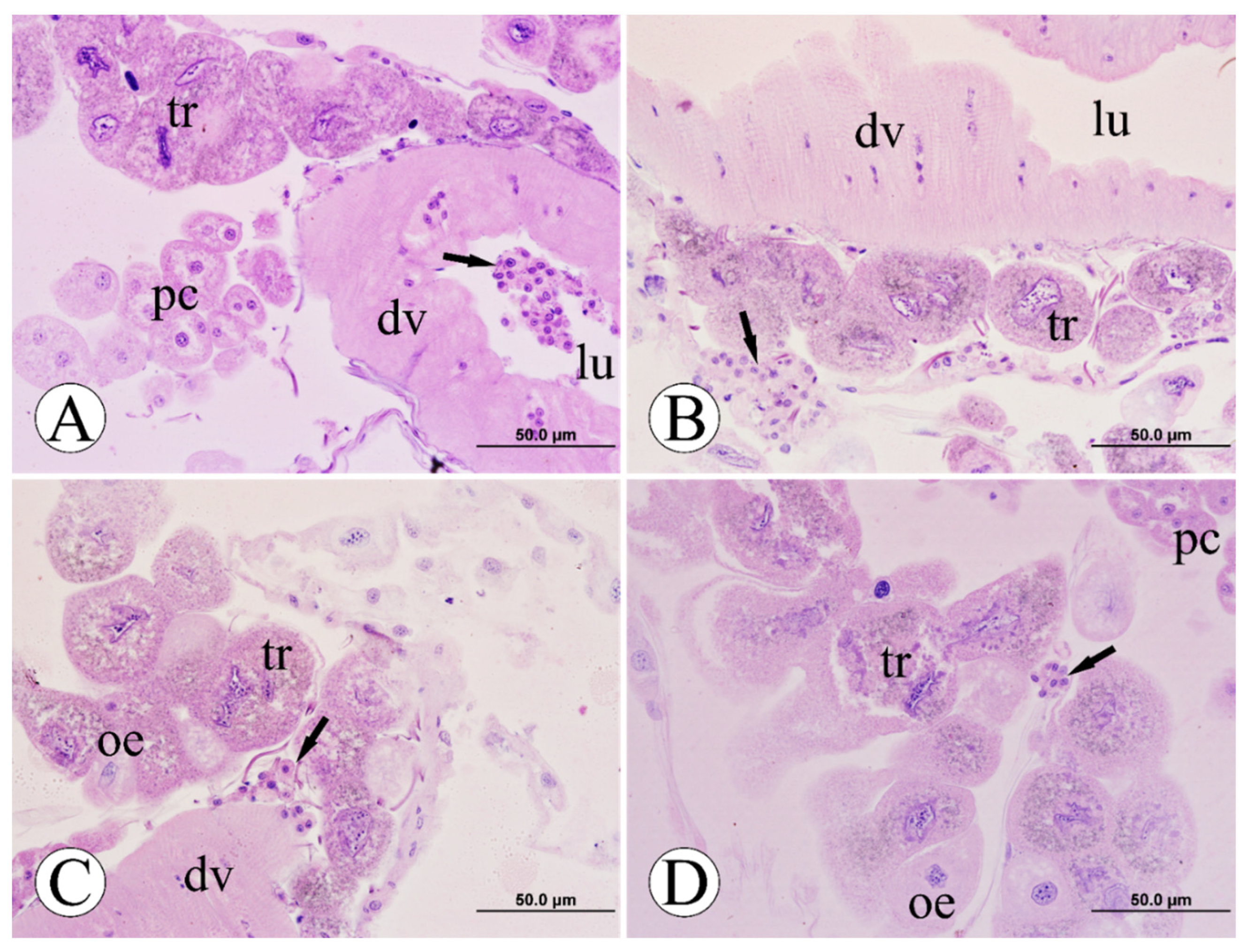
3.3. Morphological Analyses of Pericardial Cells
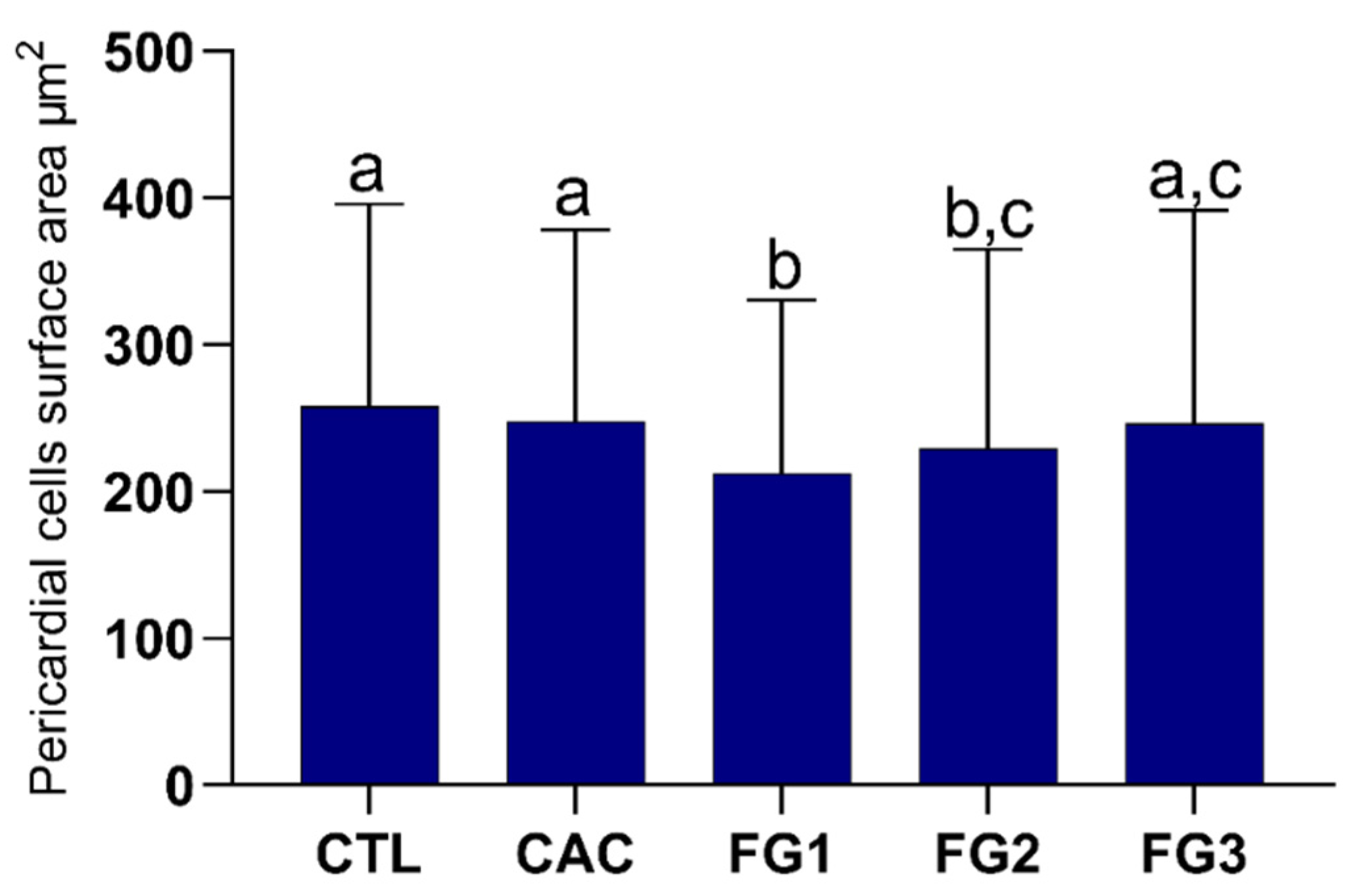
3.4. Morphometric Analyses of Pericardial Cells
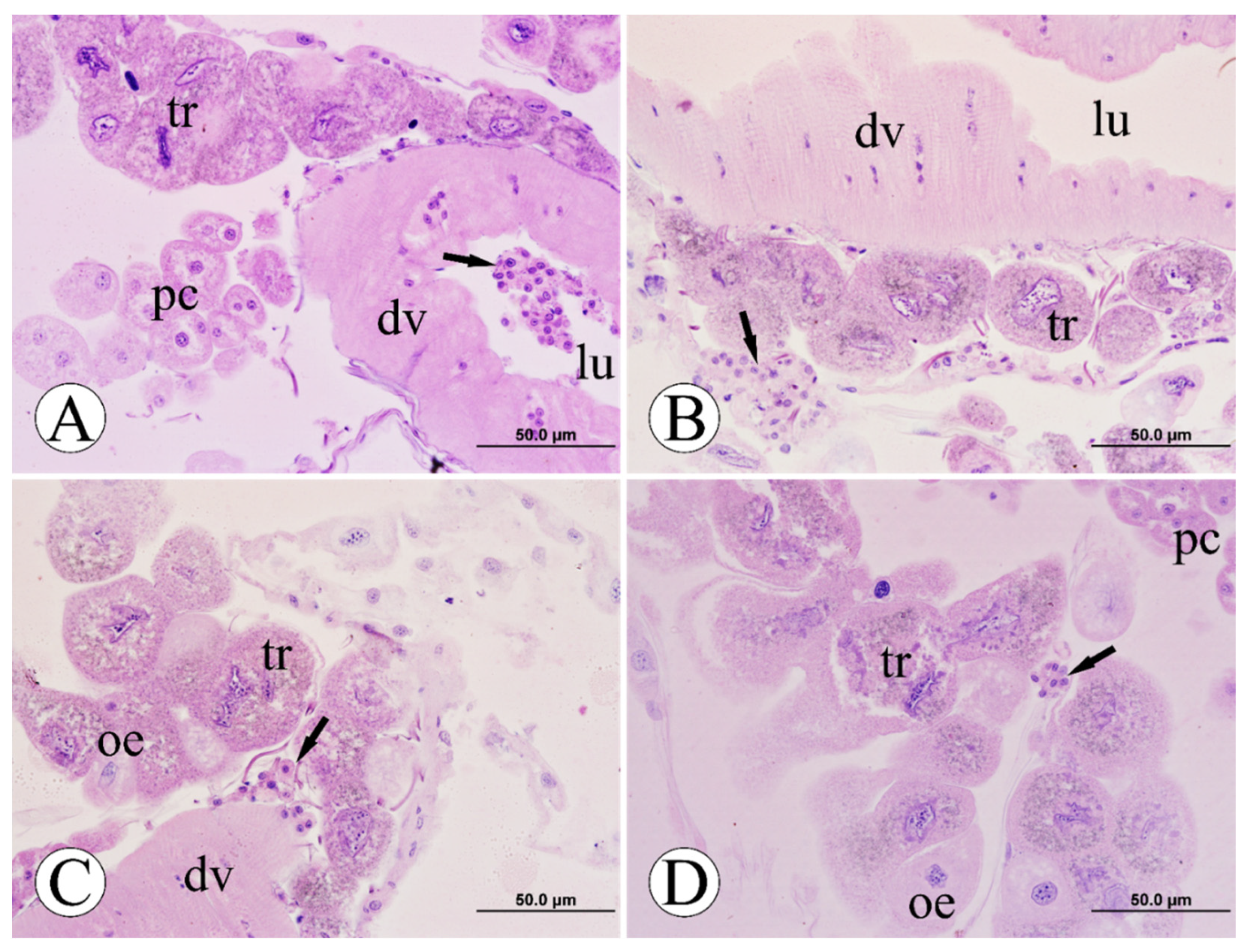
3.5. Immune System Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morse, R.A.; Calderone, N.W. The value of honey bees as pollinators of U.S. Crops in 2000. Bee Culture 2000, 128, 1–15. [Google Scholar]
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Calderone, N.W. Insect pollinated crops, insect pollinators and US agriculture: Trend analysis of aggregate data for the period 1992–2009. PLoS ONE 2012, 7, e37235. [Google Scholar] [CrossRef] [PubMed]
- Hung, K.-L.J.; Kingston, J.M.; Albrecht, M.; Holway, D.A.; Kohn, J.R. The worldwide importance of honey bees as pollinators in natural habitats. Proc. R. Soc. B 2018, 285, 20172140. [Google Scholar] [CrossRef]
- Matias, D.M.S.; Leventon, J.; Rau, A.-L.; Borgemesiter, C.; von Wehrden, H. A review of ecosystem service benefits from wild bees across social contexts. Ambio 2017, 46, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Katumo, D.M.; Liang, H.; Ochola, A.C.; Lv, M.; Wang, Q.-F.; Yang, C.-F. Pollinator diversity benefits natural and agricultural ecosystems, environmental health, and human welfare. Plant Divers. 2022. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.R.; Tarpy, D.R.; vanEngelsdorp, D.; Chauzat, M.-P.; Cox-Foster, D.L.; Delaplane, K.S.; Neumann, P.; Pettis, J.S.; Rogers, R.E.L.; Shutler, D. Colony Collapse Disorder in context. Bioessays 2010, 32, 845–846. [Google Scholar] [CrossRef]
- Hristov, P.; Shumkova, R.; Palova, N.; Neov, B. Factors Associated with Honey Bee Colony Losses: A Mini-Review. Vet. Sci. 2020, 7, 166. [Google Scholar] [CrossRef]
- Potts, S.G.; Roberts, S.P.; Dean, R.; Marris, G.; Brown, M.A.; Jones, R.; Neumann, P.; Settele, J. Declines of managed honey bees and beekeepers in Europe. J. Apic. Res. 2010, 49, 15–22. [Google Scholar] [CrossRef]
- McMenamin, A.J.; Genersch, E. Honey bee colony losses and associated viruses. Curr. Opin. Insect Sci. 2015, 8, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Drossart, M.; Gérard, M. Beyond the decline of wild bees: Optimizing conservation measures and bringing together the actors. Insects 2020, 11, 649. [Google Scholar] [CrossRef] [PubMed]
- Zattara, E.E.; Aizen, M.A. Worldwide occurrence records suggest a global decline in bee species richness. One Earth 2021, 4, 114–123. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Tanveer, M.; Sidhu, G.P.S.; Handa, N.; Kohli, S.K.; Yadav, P.; Bali, A.S.; Parihar, R.D.; et al. Worldwide pesticide usage and its impacts on ecosystem. SN Appl. Sci. 2019, 1, 1446. [Google Scholar] [CrossRef]
- Kaila, L.; Ketola, J.; Toivonen, M.; Loukola, O.; Hakala, K.; Raiskio, S.; Hurme, T.; Jalli, M. Pesticide residues in honeybee-collected pollen: Does the EU regulation protect honeybees from pesticides? Environ. Sci. Pollut. Res. 2022, 29, 18225–18244. [Google Scholar] [CrossRef] [PubMed]
- Gradish, A.E.; van der Steen, J.; Scott-Dupree, C.D.; Cabrera, A.R.; Cutler, G.C.; Goulson, D.; Klein, O.; Lehmann, D.M.; Lückmann, J.; O’Neill, B.; et al. Comparison of pesticide exposure in honey bees (Hymenoptera: Apidae) and bumble bees (Hymenoptera: Apidae): Implications for risk assessments. Environ. Entomol. 2019, 48, 12–21. [Google Scholar] [CrossRef]
- Pires, C.S.S.; Pereira, F.M.; Lopes, M.T.R.; Nocelli, R.C.F.; Malaspina, O.; Pettis, J.S.; Teixeira, E.W. Weakness and collapse of bee colonies in Brazil: Are there cases of CCD? Pesqui. Agropecu. Bras. 2016, 51, 422–442. [Google Scholar] [CrossRef]
- Assis, J.C.; Tadei, R.; Menezes-Oliveira, V.B.; Silva-Zacarin, E.C.M. Are native bees in Brazil at risk from the exposure to the neonicotinoid imidacloprid? Environ. Res. 2022, 212, 113127. [Google Scholar] [CrossRef]
- Benuszak, B.; Laurent, M.; Chauzat, M.P. The exposure of honey bees (Apis mellifera; Hymenoptera: Apidae) to pesticides: Room for improvement in research. Sci. Total Environ. 2017, 587–588, 423–438. [Google Scholar] [CrossRef]
- Cullen, M.G.; Thompson, L.J.; Carolan, J.C.; Stout, J.C.; Stanley, D.A. Fungicides, herbicides and bees: A systematic review of existing research and methods. PLoS ONE 2019, 14, e0225743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rondeau, S.; Raine, N.E. Fungicides and bees: A review of exposure and risk. Environ. Int. 2022, 165, 107311. [Google Scholar] [CrossRef] [PubMed]
- Tadei, R.; Domingues, C.E.C.; Malaquias, J.B.; Camilo, E.V.; Malaspina, O.; Silva-Zacarin, E.C.M. Late effect of larval co-exposure to the insecticide clothianidin and fungicide pyraclostrobin in Africanized Apis mellifera. Sci. Rep. 2019, 9, 3277. [Google Scholar] [CrossRef] [PubMed]
- Domingues, C.E.C.; Tadei, R.; Inoue, L.V.B.; Silva-Zacarin, E.C.M.; Malaspina, O. Effects of larval exposure to the fungicide pyraclostrobin on the post-embryonic development of Africanized Apis mellifera workers. Environ. Adv. 2021, 4, 100069. [Google Scholar] [CrossRef]
- Batista, A.C.; Domingues, C.E.C.; Costa, M.J.; Silva-Zacarin, E.C.M. Is a strobilurin fungicide capable of inducing histopathological effects on the midgut and Malpighian tubules of honey bees? J. Apic. Res. 2020, 59, 1–10. [Google Scholar] [CrossRef]
- Nicodemo, D.; Mingatto, F.E.; De Jong, D.; Bizerra, P.F.V.; Tavares, M.A.; Bellini, W.C.; Vicente, E.F.; Carvalho, A. Mitochondrial respiratory inhibition promoted by pyraclostrobin in fungi is also observed in honey bees. Environ. Toxicol. Chem. 2020, 39, 1267–1272. [Google Scholar] [CrossRef]
- Degrandi-Hoffman, G.; Chen, Y.; Dejong, E.W.; Chambers, M.L.; Hidalgo, G. Effects of oral exposure to fungicides on honey bee nutrition and virus levels. J. Econ. Entomol. 2015, 108, 2518–2528. [Google Scholar] [CrossRef]
- Carneiro, L.S.; Martínez, L.C.; Gonçalves, W.G.; Santana, L.M.; Serrao, J.E. The fungicide iprodione affects midgut cells of non-target honey bee Apis mellifera workers. Ecotoxicol. Environ. Saf. 2020, 189, 109991. [Google Scholar] [CrossRef]
- Brazilian Health Regulatory Agency (ANVISA in Portuguese). 2022. Available online: https://www.gov.br/anvisa/pt-br/setorregulado/regularizacao/agrotoxicos/monografias/monografias-autorizadas/p/4488json-file-1 (accessed on 1 August 2022).
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; Parr-Dobrzanski, B. The strobilurin fungicides. Pest Manag. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef]
- Domingues, C.E.C.; Inoue, L.V.B.; Silva-Zacarin, E.C.M.; Malaspina, O. Foragers of Africanized honeybee are more sensitive to fungicide pyraclostrobin than newly emerged bees. Environ. Pollut. 2020, 266, 115267. [Google Scholar] [CrossRef]
- Tadei, R.; Menezes-Oliveira, V.B.; Silva-Zacarin, E.C.M. Silent effect of the fungicide pyraclostrobin on the larval exposure of the non-target organism Africanized Apis mellifera and its interaction with the pathogen Nosema ceranae in adulthood. Environ. Pollut. 2020, 267, 115622. [Google Scholar] [CrossRef] [PubMed]
- Zioga, E.; Kelly, R.; White, B.; Stout, J.C. Plant protection product residues in plant pollen and nectar: A review of current knowledge. Environ. Res. 2020, 189, 109873. [Google Scholar] [CrossRef]
- Malaspina, O.; Silva-Zacarin, E.C.M. Cell markers for ecotoxicological studies in target organs of bees. Braz. J. Morphol. Sci. 2006, 23, 303–309. [Google Scholar]
- Skowronek, P.; Wójcik, L.; Strachecka, A. Fat Body—Multifunctional Insect Tissue. Insects 2021, 12, 547. [Google Scholar] [CrossRef] [PubMed]
- Mills, R.P.; King, R.C. The pericardial cells of Drosophila melanogaster. J. Cell Sci. 1965, 106, 261–268. [Google Scholar] [CrossRef]
- Abdalla, F.C.; Domingues, C.E.C. Hepato-nephrocitic system: A novel model of biomarkers for analysis of the ecology of stress in environmental biomonitoring. PLoS ONE 2015, 10, e0132349. [Google Scholar] [CrossRef]
- Domingues, C.E.C.; Inoue, L.V.B.; Silva-Zacarin, E.C.M.; Malaspina, O. Fungicide pyraclostrobin affects midgut morphophysiology and reduces survival of Brazilian native stingless bee Melipona scutellaris. Ecotoxicol. Environ. Saf. 2020, 206, 111395. [Google Scholar] [CrossRef] [PubMed]
- Pettis, J.S.; Lichtenberg, E.M.; Andree, M.; Stitzinger, J.; Rose, R.; vanEngelsdorp, D. Crop pollination exposes honey bees to pesticides which alters their susceptibility to the gut pathogen Nosema ceranae. PLoS ONE 2013, 8, e70182. [Google Scholar] [CrossRef]
- Yoder, J.A.; Jajack, A.J.; Rosselot, A.E.; Smith, T.J.; Yerke, M.C.; Sammataro, D. Fungicide contamination reduces beneficial fungi in bee bread based on an area-wide field study in honey bee, Apis mellifera, colonies. J. Toxicol. Environ. Health Part A 2013, 76, 587–600. [Google Scholar] [CrossRef]
- OECD (Organization for Economic Co-operation and Development). OECD Guidelines for the Testing of Chemicals; Honeybees, Acute Oral Toxicity Test N°213; OECD Publishing: Paris, France, 1998.
- Silva-Zacarin, E.C.M.; Chauzat, M.P.; Zeggane, S.; Drajnudel, P.; Schurr, F.; Faucon, J.P.; Malaspina, O.; Engler, J.A. Protocol for optimization of histological, histochemical and immunohistochemical analyses of larval tissues: Application in histopathology of honey bee. In Current Microscopy Contributions to Advances in Science and Technology, 1st ed.; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2012; Volume 5, pp. 696–703. [Google Scholar]
- Junqueira, L.C.U.; Junqueira, L.M.M.S. Técnicas Básicas de Citologia e Histologia, 1st ed.; Editora Santos: São Paulo, Brazil, 1983; pp. 1–123. [Google Scholar]
- Balsamo, P.J.; Domingues, C.E.C.; Silva-Zacarin, E.C.M.; Gregorc, A.; Irazusta, S.P.; Salla, R.F.; Costa, M.J.; Abdalla, F.C. Impact of sublethal doses of thiamethoxam and inoculation on the hepato-nephrocitic system in young Africanized. J. Apic. Res. 2020, 59, 350–361. [Google Scholar] [CrossRef]
- Seeley, T.D. Honey bee foragers as sensory units of their colonies. Behav. Ecol. Sociobiol. 1994, 34, 51–62. [Google Scholar] [CrossRef]
- More, S.; Bampidis, V.; Benford, D.; Bragard, C.; Halldorsson, T.; Hernández-Jerez, A.; Bennekou, S.H.; Koutsoumanis, K.; Machera, K.; Naegeli, H.; et al. A systems-based approach to the environmental risk assessment of multiple stressors in honey bees. EFSA J. 2021, 19, e06607. [Google Scholar] [CrossRef] [PubMed]
- Zaluski, R.; Justulin, L.A.; Orsi, R.D.O. Field-relevant doses of the systemic insecticide fipronil and fungicide pyraclostrobin impair mandibular and hypopharyngeal glands in nurse honeybees (Apis mellifera). Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Cruz-Landim, C. Abelhas: Morfologia e Funções de Sistemas, 1st ed.; UNESP: São Paulo, Brazil, 2009; pp. 1–416. [Google Scholar]
- de Oliveira, V.T.P.; Cruz-Landim, C. Morphology and function of insect fat body cells: A review. R. Bras. Bioci. 2003, 11, 195–205. [Google Scholar]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef]
- Domingues, C.E.C.; Abdalla, F.C.; Balsamo, P.J.; Pereira, B.V.R.; Hausen, M.A.; Costa, M.J.; Silva-Zacarin, E.C.M. Thiamethoxam and picoxystrobin reduce the survival and overload the hepato-nephrocitic system of the Africanized honeybee. Chemosphere 2017, 186, 994–1005. [Google Scholar] [CrossRef]
- Cousin, M.; Silva-Zacarin, E.C.M.; Kretzschmar, A.; El Maataoui, M.; Brunet, J.-L.; Belzunces, L.P. Size changes in honey bee larvae oenocytes induced by exposure to Paraquat at very low concentrations. PLoS ONE 2013, 8, e65693. [Google Scholar] [CrossRef] [PubMed]
- Roma, G.C.; Bueno, O.C.; Camargo-Mathias, M.I. Morphophysiological analysis of the insect fat body: A review. Micron 2010, 41, 395–401. [Google Scholar] [CrossRef]
- Fife, H.G.; Palli, S.R.; Locke, M. A function for pericardial cells in an insect. Insect Biochem. 1987, 17, 829–840. [Google Scholar] [CrossRef]
- Balsamo, P.J.; Nogueira, F.L.A.; Ceschi-Bertoli, L.; Salla, R.F.; Maia, F.M.C.; Irazusta, S.P.; Boeing, G.A.N.S.; Abdalla, F.C. Effect of a 110 ppb mercury exposition on neotropical bumble bee workers, Bombus atratus: In situ localization of Hsp70 and Hsp90 and general morphological changes of hepato-nephrocitic cells. J. Apic. Res. 2022, 61, 1–9. [Google Scholar] [CrossRef]
- Nogueira, F.L.A.; Balsamo, P.J.; Costa, M.J.; Abdalla, F.C. Exposure to Mercury at Trace Concentrations Leads to Collapse of the Hepato-Nephrocitic System in Two Neotropical Species of Bumblebee. Am. J. Agric. Biol. Sci. 2019, 14, 1–10. [Google Scholar] [CrossRef]
- Tosi, S.; Sfeir, C.; Camesecchi, E.; vanEngelsdorp, D.; Chauzat, M.-P. Lethal, sublethal, and combined effects of pesticides on bees: A meta-analysis. Sci. Total Environ. 2022, 844, 156857. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inoue, L.V.B.; Domingues, C.E.C.; Gregorc, A.; Silva-Zacarin, E.C.M.; Malaspina, O. Harmful Effects of Pyraclostrobin on the Fat Body and Pericardial Cells of Foragers of Africanized Honey Bee. Toxics 2022, 10, 530. https://doi.org/10.3390/toxics10090530
Inoue LVB, Domingues CEC, Gregorc A, Silva-Zacarin ECM, Malaspina O. Harmful Effects of Pyraclostrobin on the Fat Body and Pericardial Cells of Foragers of Africanized Honey Bee. Toxics. 2022; 10(9):530. https://doi.org/10.3390/toxics10090530
Chicago/Turabian StyleInoue, Lais V. B., Caio E. C. Domingues, Aleš Gregorc, Elaine C. M. Silva-Zacarin, and Osmar Malaspina. 2022. "Harmful Effects of Pyraclostrobin on the Fat Body and Pericardial Cells of Foragers of Africanized Honey Bee" Toxics 10, no. 9: 530. https://doi.org/10.3390/toxics10090530
APA StyleInoue, L. V. B., Domingues, C. E. C., Gregorc, A., Silva-Zacarin, E. C. M., & Malaspina, O. (2022). Harmful Effects of Pyraclostrobin on the Fat Body and Pericardial Cells of Foragers of Africanized Honey Bee. Toxics, 10(9), 530. https://doi.org/10.3390/toxics10090530