


Copper Effect on Microalgae: Toxicity and Bioremediation Strategies


 , ,
, ,  and
and 
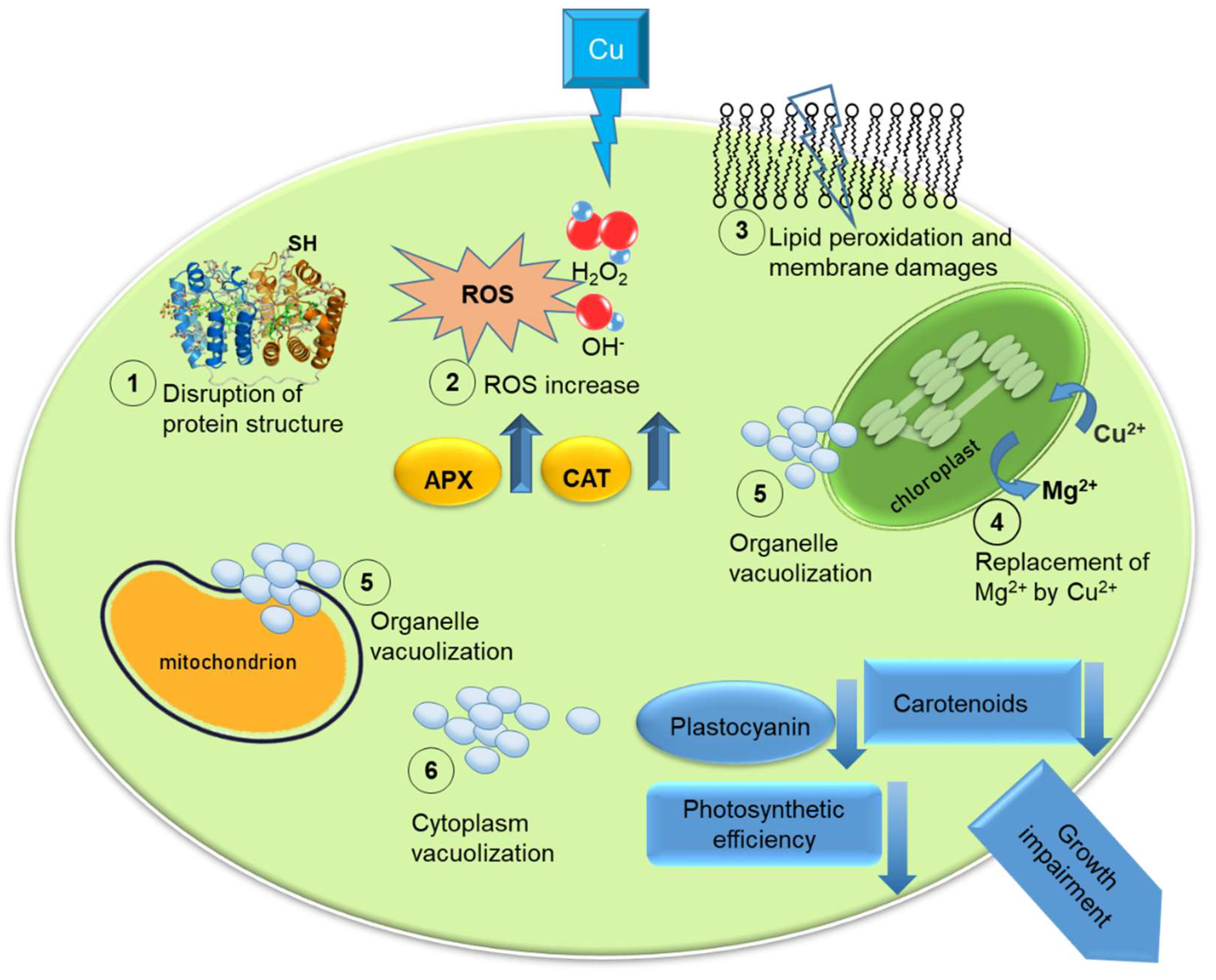
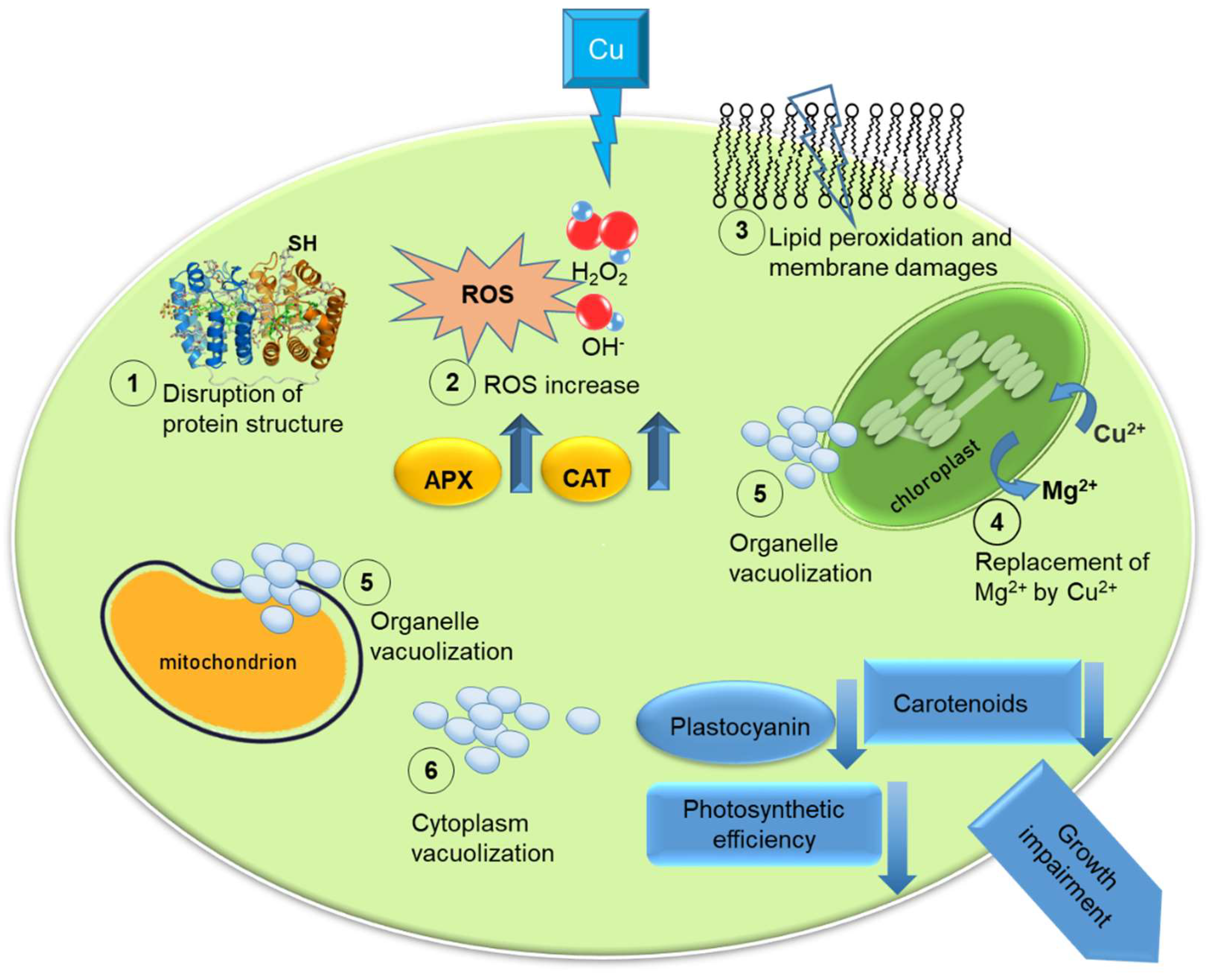{kind=link}
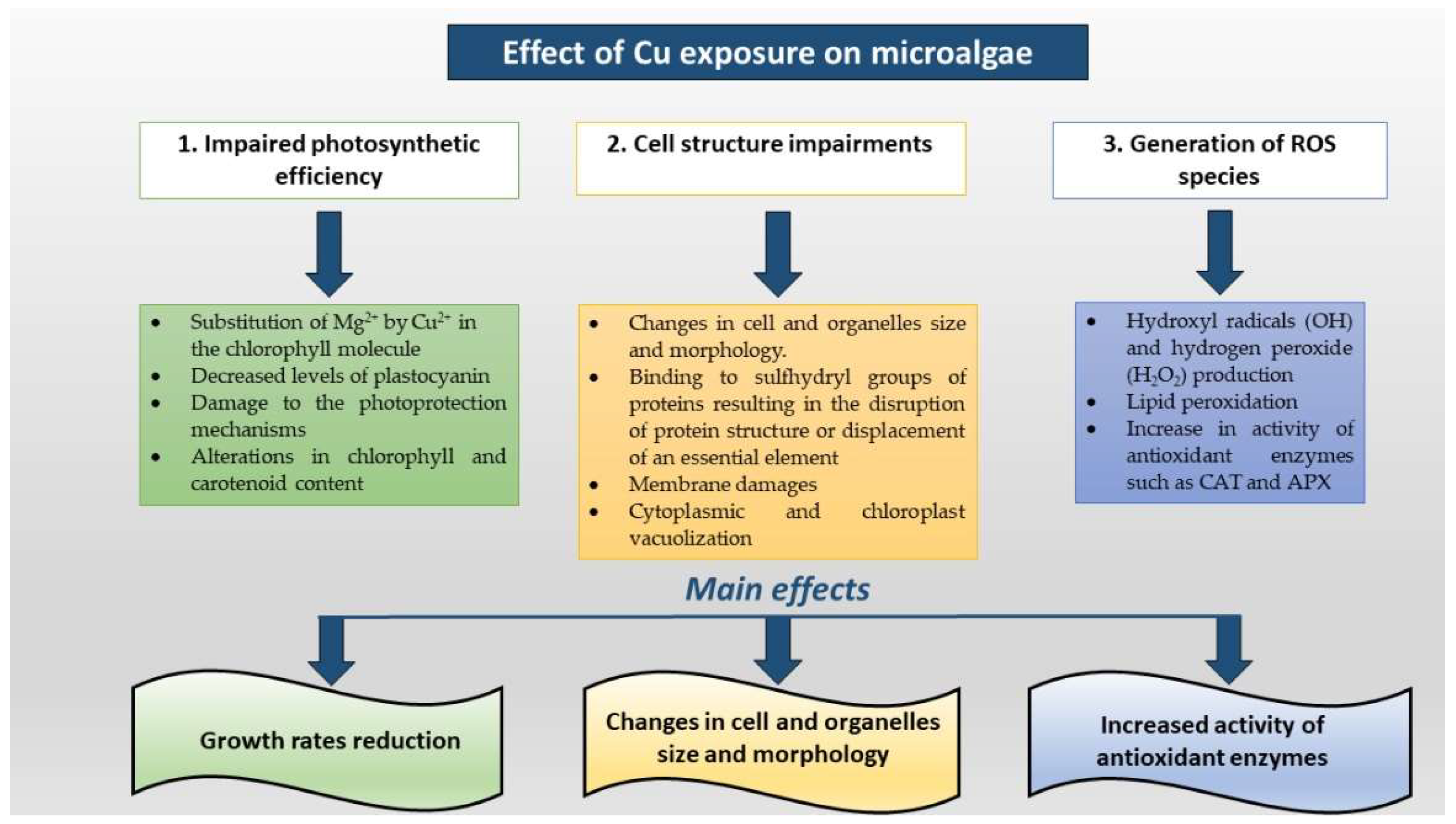
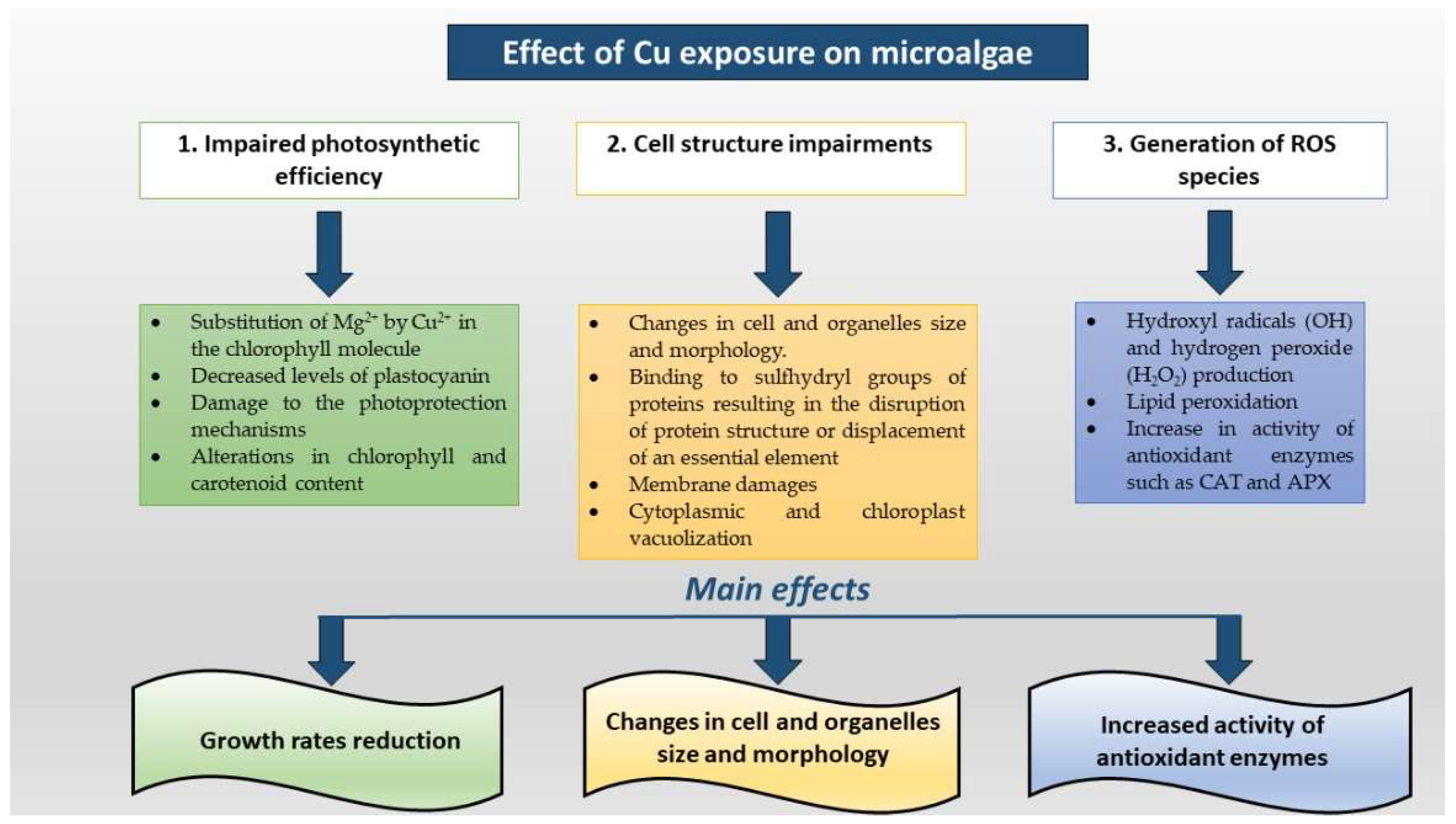{kind=link}
Abstract
:1. Introduction
2. The Role of Cu as Essential Metal in Microalgal Metabolism
3. Mechanisms of Adsorption/Compartmentalization
4. Detrimental Effects of Cu at High Concentrations
5. Generation of Reactive Oxygen Species
6. Phytochelatins and Metallothioneins Protective Role against Heavy Metal Toxicity
7. Synergistic Effect of Cu and Other Pollutants
8. Microalgal Potential for HM Removal, with a Special Focus on Cu: Possibilities and Constraints
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ali, H.; Khan, E. What are heavy metals? Long-standing controversy over the scientific use of the term ‘heavy metals’—Proposal of a comprehensive definition. Toxicol. Environ. Chem. 2018, 100, 6–19. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. Exp. Suppl. 2012, 101, 133–164. [Google Scholar] [CrossRef]
- He, Z.L.L.; Yang, X.E.; Stoffella, P.J. Trace elements in agroecosystems and impacts on the environment. J. Trace Elem. Med. Biol. 2005, 19, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.B.; Frid, C.; Attrill, M. Marine Pollution, 5th ed.; Oxford University Press: Oxford, UK; New York, NY, USA, 2001; 236p. [Google Scholar]
- Fu, Z.S.; Xi, S.H. The effects of heavy metals on human metabolism. Toxicol. Mech. Methods 2020, 30, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Rial, D.; Beiras, R. Prospective ecological risk assessment of sediment resuspension in an estuary. J. Environ. Monit. 2012, 14, 2137–2144. [Google Scholar] [CrossRef] [PubMed]
- Fichet, D.; Radenac, G.; Miramand, P. Experimental studies of impacts of harbour sediments resuspension to marine invertebrates larvae: Bioavailability of Cd, Cu, Pb and Zn and toxicity. Mar. Pollut. Bull. 1998, 36, 509–518. [Google Scholar] [CrossRef]
- Shull, D. Bioturbation. In Encyclopedia of Ocean Sciences, Vol 3: Ocean Dynamics, 3rd ed.; Cochran, J.K., Bokuniewicz, H.J., Yager, P.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 671–676. [Google Scholar]
- Cordoba-Tovar, L.; Marrugo-Negrete, J.; Baron, P.R.; Diez, S. Drivers of biomagnification of Hg, As and Se in aquatic food webs: A review. Environ. Res. 2022, 204, 112226. [Google Scholar] [CrossRef]
- Wu, P.P.; Zakem, E.J.; Dutkiewicz, S.; Zhang, Y.X. Biomagnification of Methylmercury in a Marine Plankton Ecosystem. Environ. Sci. Technol. 2020, 54, 5446–5455. [Google Scholar] [CrossRef]
- Xie, Q.; Gui, D.; Liu, W.; Wu, Y.P. Risk for Indo-Pacific humpback dolphins (Sousa chinensis) and human health related to the heavy metal levels in fish from the Pearl River Estuary, China. Chemosphere 2020, 240, 124844. [Google Scholar] [CrossRef]
- Vu, C.T.; Lin, C.; Yeh, G.; Villanueva, M.C. Bioaccumulation and potential sources of heavy metal contamination in fish species in Taiwan: Assessment and possible human health implications. Environ. Sci. Pollut. Res. 2017, 24, 19422–19434. [Google Scholar] [CrossRef]
- Köker, L.; Aydın, F.; Gaygusuz, Ö.; Akçaalan, R.; Çamur, D.; İlter, H.; Ayoğlu, F.N.; Altın, A.; Topbaş, M.; Albay, M. Heavy Metal Concentrations in Trachurus mediterraneus and Merlangius merlangus Captured from Marmara Sea, Turkey and Associated Health Risks. Environ. Manag. 2021, 67, 522–531. [Google Scholar] [CrossRef]
- De Mora, S.; Fowler, S.W.; Wyse, E.; Azemard, S. Distribution of heavy metals in marine bivalves, fish and coastal sediments in the Gulf and Gulf of Oman. Mar. Pollut. Bull. 2004, 49, 410–424. [Google Scholar] [CrossRef]
- Culha, S.T.; Culha, M.; Karayucel, I.; Celik, M.Y.; Isler, Y. Heavy metals in Mytilus galloprovincialis, suspended particulate matter and sediment from offshore submerged longline system, Black Sea. Int. J. Environ. Sci. Technol. 2017, 14, 385–396. [Google Scholar] [CrossRef]
- Mejdoub, Z.; Zaid, Y.; Hmimid, F.; Kabine, M. Assessment of metals bioaccumulation and bioavailability in mussels Mytilus galloprovincialis exposed to outfalls pollution in coastal areas of Casablanca. J. Trace Elem. Med. Biol. 2018, 48, 30–37. [Google Scholar] [CrossRef]
- Joksimovic, D.; Castelli, A.; Perosevic, A.; Djurovic, D.; Stankovic, S. Determination of trace metals in Mytilus galloprovincialis along the Boka Kotorska Bay, Montenegrin coast. J. Trace Elem. Med. Biol. 2018, 50, 601–608. [Google Scholar] [CrossRef]
- Kouali, H.; Chaouti, A.; Achtak, H.; Elkalay, K.; Dahbi, A. Trace metal contents in the mussel Mytilus galloprovincialis from Atlantic coastal areas in northwestern Morocco: Levels of contamination and assessment of potential risks to human health. Mar. Pollut. Bull. 2022, 179, 113680. [Google Scholar] [CrossRef]
- Mehri, A. Trace Elements in Human Nutrition (II)—An Update. Int. J. Prev. Med. 2020, 11, 2. [Google Scholar] [CrossRef]
- World Health Organization; International Atomic Energy Agency; Food and Agriculture Organization of the United Nations. Trace Elements in Human Nutrition and Health; World Health Organization: Geneva, Switzerland, 1996; ISBN 9241561734. [Google Scholar]
- Roveta, C.; Annibaldi, A.; Afghan, A.; Calcinai, B.; Di Camillo, C.G.; Gregorin, C.; Illuminati, S.; Pulido Mantas, T.; Truzzi, C.; Puce, S. Biomonitoring of Heavy Metals: The Unexplored Role of Marine Sessile Taxa. Appl. Sci. 2021, 11, 580. [Google Scholar] [CrossRef]
- Jovic, M.; Onjia, A.; Stankovic, S. Toxic metal health risk by mussel consumption. Environ. Chem. Lett. 2012, 10, 69–77. [Google Scholar] [CrossRef]
- Ramirez, R. The gastropod Osilinus atrata as a bioindicator of Cd, Cu, Pb and Zn contamination in the coastal waters of the Canary Islands. Chem. Ecol. 2013, 29, 208–220. [Google Scholar] [CrossRef]
- Tsang, T.; Davis, C.I.; Brady, D.C. Copper biology. Curr. Biol. CB 2021, 31, R421–R427. [Google Scholar] [CrossRef] [PubMed]
- Scheiber, I.; Dringen, R.; Mercer, J.F. Copper: Effects of deficiency and overload. Met. Ions Life Sci. 2013, 13, 359–387. [Google Scholar] [CrossRef]
- Franchitto, N.; Gandia-Mailly, P.; Georges, B.; Galinier, A.; Telmon, N.; Ducasse, J.L.; Rouge, D. Acute copper sulphate poisoning: A case report and literature review. Resuscitation 2008, 78, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Amuda, O.S.; Alade, A.O.; Hung, Y.T.; Wang, L.K.; Wang, M.H.S. Toxicity, Sources, and Control of Copper (Cu), Zinc (Zn), Molybdenum (Mo), Silver (Ag), and Rare Earth Elements in the Environment. Civ. Environ. Eng. Fac. Publ. 2016, 331, 1–25. [Google Scholar]
- Zolkefli, N.; Sharuddin, S.S.; Yusoff, M.Z.M.; Hassan, M.A.; Maeda, T.; Ramli, N. A Review of Current and Emerging Approaches for Water Pollution Monitoring. Water 2020, 12, 3417. [Google Scholar] [CrossRef]
- Nowicka, B. Heavy metal-induced stress in eukaryotic algae-mechanisms of heavy metal toxicity and tolerance with particular emphasis on oxidative stress in exposed cells and the role of antioxidant response. Environ. Sci. Pollut. Res. 2022, 29, 16860–16911. [Google Scholar] [CrossRef]
- Marella, T.K.; Saxena, A.; Tiwari, A. Diatom mediated heavy metal remediation: A review. Bioresour. Technol. 2020, 305, 123068. [Google Scholar] [CrossRef] [PubMed]
- Danouche, M.; El Ghachtouli, N.; El Arroussi, H. Phycoremediation mechanisms of heavy metals using living green microalgae: Physicochemical and molecular approaches for enhancing selectivity and removal capacity. Heliyon 2021, 7, e07609. [Google Scholar] [CrossRef]
- Goswami, R.K.; Agrawal, K.; Shah, M.P.; Verma, P. Bioremediation of heavy metals from wastewater: A current perspective on microalgae-based future. Lett. Appl. Microbiol. 2021. [Google Scholar] [CrossRef]
- Crichton, R.R.; Pierre, J.L. Old iron, young copper: From Mars to Venus. Biometals 2001, 14, 99–112. [Google Scholar] [CrossRef]
- Quigg, A. Micronutrients. In The Physiology of Microalgae, Vol 6: Developments in Applied Phycology; Borowitzka, M., Beardall, J., Raven, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 211–231. [Google Scholar]
- De la Cerda, B.; Castielli, O.; Durán, R.V.; Navarro, J.A.; Hervás, M.; De la Rosa, M.A. A proteomic approach to iron and copper homeostasis in cyanobacteria. Brief Funct. Genom. Proteom. 2007, 6, 322–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kropat, J.; Gallaher, S.D.; Urzica, E.I.; Nakamoto, S.S.; Strenkert, D.; Tottey, S.; Mason, A.Z.; Merchant, S.S. Copper economy in Chlamydomonas: Prioritized allocation and reallocation of copper to respiration vs. photosynthesis. Proc. Natl. Acad. Sci. USA 2015, 112, 2644–2651. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Cao, W.; Du, H.; Liu, W.H.; Li, P. Increasing Temperature Alters the Effects of Extracellular Copper on Thalassiosira Pseudonana Physiology and Transcription. J. Mar. Sci. Eng. 2021, 9, 816. [Google Scholar] [CrossRef]
- Maldonado, M.T.; Allen, A.E.; Chong, J.S.; Lin, K.; Leus, D.; Karpenko, N.; Harris, S.L. Copper-dependent iron transport in coastal and oceanic diatoms. Limnol. Oceanogr. 2006, 51, 1729–1743. [Google Scholar] [CrossRef]
- La Fontaine, S.; Quinn, J.M.; Nakamoto, S.S.; Page, M.D.; Gohre, V.; Mosely, J.L.; Kropat, J.; Merchant, S. Copper-dependent iron assimilation pathway in the model photosynthetic eukaryote Chlamydomonas reinhardtii. Eukaryot. Cell 2002, 1, 736–757. [Google Scholar] [CrossRef]
- Guo, J.; Lapi, S.; Ruth, T.J.; Maldonado, M.T. The effects of iron and copper availability on the copper stoichiometry on marine phytoplankton. J. Phycol. 2012, 48, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Annett, A.L.; Lapi, S.; Ruth, T.J.; Maldonado, M.T. The effects of Cu and Fe availability on the growth and Cu : C ratios of marine diatoms. Limnol. Oceanogr. 2008, 53, 2451–2461. [Google Scholar] [CrossRef]
- Peers, G.; Quesnel, S.A.; Price, N.M. Copper requirements for iron acquisition and growth of coastal and oceanic diatoms. Limnol. Oceanogr. 2005, 50, 1149–1158. [Google Scholar] [CrossRef]
- Silva, J.C.; Echeveste, P.; Lombardi, A.T. Higher biomolecules yield in phytoplankton under copper exposure. Ecotoxicol. Environ. Saf. 2018, 161, 57–63. [Google Scholar] [CrossRef]
- El Agawany, N.; Kaamoush, M.; El-Zeiny, A.; Ahmed, M. Effect of heavy metals on protein content of marine unicellular green alga Dunaliella tertiolecta. Environ. Monit. Assess. 2021, 193, 584. [Google Scholar] [CrossRef]
- Kong, L.; Price, N.M. Identification of copper-regulated proteins in an oceanic diatom, Thalassiosira oceanica 1005. Met. Integr. Biomet. Sci. 2020, 12, 1106–1117. [Google Scholar] [CrossRef] [PubMed]
- Rocha, G.S.; Parrish, C.C.; Espindola, E.L.G. Effects of copper on photosynthetic and physiological parameters of a freshwater microalga (Chlorophyceae). Algal Res. Biomass Biofuels Bioprod. 2021, 54, 102223. [Google Scholar] [CrossRef]
- Torres, M.A.; Barros, M.P.; Campos, S.C.G.; Pinto, E.; Rajamani, S.; Sayre, R.T.; Colepicolo, P. Biochemical biomarkers in algae and marine pollution: A review. Ecotoxicol. Environ. Saf. 2008, 71, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Das, N.; Vimala, R.; Karthika, P. Biosorption of heavy metals—An overview. Indian J. Biotechnol. 2008, 7, 159–169. [Google Scholar]
- He, J.S.; Chen, J.P. A comprehensive review on biosorption of heavy metals by algal biomass: Materials, performances, chemistry, and modeling simulation tools. Bioresour. Technol. 2014, 160, 67–78. [Google Scholar] [CrossRef]
- Levy, J.L.; Angel, B.M.; Stauber, J.L.; Poon, W.L.; Simpson, S.L.; Cheng, S.H.; Jolley, D.F. Uptake and internalisation of copper by three marine microalgae: Comparison of copper-sensitive and copper-tolerant species. Aquat. Toxicol. 2008, 89, 82–93. [Google Scholar] [CrossRef]
- Narula, P.; Mahajan, A.; Gurnani, C.; Kumar, V.; Mukhija, S. Microalgae as an Indispensable Tool against Heavy Metals Toxicity to Plants: A Review. Int. J. Pharm. Sci. Rev. Res. 2015, 31, 86–93. [Google Scholar]
- Quigg, A.; Reinfelder, J.R.; Fisher, N.S. Copper uptake kinetics in diverse marine phytoplankton. Limnol. Oceanogr. 2006, 51, 893–899. [Google Scholar] [CrossRef]
- Croot, P.L.; Karlson, B.; van Elteren, J.T.; Kroon, J.J. Uptake and efflux of Cu-64 by the marine cyanobacterium Synechococcus (WH7803). Limnol. Oceanogr. 2003, 48, 179–188. [Google Scholar] [CrossRef]
- Blaby-Haas, C.E.; Merchant, S.S. The ins and outs of algal metal transport. Biochim. Biophys. Acta 2012, 1823, 1531–1552. [Google Scholar] [CrossRef]
- Nassiri, Y.; Mansot, J.L.; Wery, J.; GinsburgerVogel, T.; Amiard, J.C. Ultrastructural and electron energy loss spectroscopy studies of sequestration mechanisms of Cd and Cu in the marine diatom Skeletonema costatum. Arch. Environ. Contam. Toxicol. 1997, 33, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.S.; Dillon, C.T.; Vogt, S.; Lai, B.; Stauber, J.; Jolley, D.F. Copper Uptake, Intracellular Localization, and Speciation in Marine Microalgae Measured by Synchrotron Radiation X-ray Fluorescence and Absorption Microspectroscopy. Environ. Sci. Technol. 2016, 50, 8827–8839. [Google Scholar] [CrossRef] [PubMed]
- Croot, P.L.; Moffett, J.W.; Brand, L.E. Production of extracellular Cu complexing ligands by eucaryotic phytoplankton in response to Cu stress. Limnol. Oceanogr. 2000, 45, 619–627. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Macias, M.D.; Correa-Murrieta, M.A.; Villegas-Peralta, Y.; Devora-Isiordia, G.E.; Alvarez-Sanchez, J.; Saldivar-Cabrales, J.; Sanchez-Duarte, R.G. Uptake of copper from acid mine drainage by the microalgae Nannochloropsis oculata. Environ. Sci. Pollut. Res. 2019, 26, 6311–6318. [Google Scholar] [CrossRef]
- Hamed, S.M.; Selim, S.; Klock, G.; AbdElgawad, H. Sensitivity of two green microalgae to copper stress: Growth, oxidative and antioxidants analyses. Ecotoxicol. Environ. Saf. 2017, 144, 19–25. [Google Scholar] [CrossRef]
- Gong, A.P.; Gu, W.M.; Zhao, Z.Y.; Shao, Y.N. Identification of heavy metal by testing microalgae using confocal Raman microspectroscopy technology. Appl. Opt. 2019, 58, 8396–8403. [Google Scholar] [CrossRef]
- Manimaran, K.; Karthikeyan, P.; Ashokkumar, S.; Prabu, V.A.; Sampathkumar, P. Effect of Copper on Growth and Enzyme Activities of Marine Diatom, Odontella mobiliensis. Bull. Environ. Contam. Toxicol. 2012, 88, 30–37. [Google Scholar] [CrossRef]
- Purbonegoro, T.; Suratno; Puspitasari, R.; Husna, N.A. Toxicity of copper on the growth of marine microalgae Pavlova sp. and its chlorophyll-a. In Proceedings of the 1st Global Colloquium on GeoSciences and Engineering (GCGE), Bandung, Indonesia, 18–19 October 2017. [Google Scholar]
- Neethu, K.V.; Saranya, K.S.; Krishna, N.G.A.; Praved, P.H.; Aneesh, B.P.; Nandan, S.B.; Marigoudar, S.R. Toxicity of copper on marine diatoms, Chaetoceros calcitrans and Nitzchia closterium from Cochin estuary, India. Ecotoxicology 2021, 30, 783–793. [Google Scholar] [CrossRef]
- Levy, J.L.; Stauber, J.L.; Jolley, D.F. Sensitivity of marine microalgae to copper: The effect of biotic factors on copper adsorption and toxicity. Sci. Total Environ. 2007, 387, 141–154. [Google Scholar] [CrossRef]
- Buayam, N.; Davey, M.P.; Smith, A.G.; Pumas, C. Effects of Copper and pH on the Growth and Physiology of Desmodesmus sp. AARLG074. Metabolites 2019, 9, 84. [Google Scholar] [CrossRef]
- Melegari, S.P.; Perreault, F.; Costa, R.H.R.; Popovic, R.; Matias, W.G. Evaluation of toxicity and oxidative stress induced by copper oxide nanoparticles in the green alga Chlamydomonas reinhardtii. Aquat. Toxicol. 2013, 142, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Hu, C.W.; Zhu, Q.; Chen, L.; Kong, Z.M.; Liu, Z.L. Copper and zinc induction of lipid peroxidation and effects on antioxidant enzyme activities in the microalga Pavlova viridis (Prymnesiophyceae). Chemosphere 2006, 62, 565–572. [Google Scholar] [CrossRef]
- Imlay, J.A. Pathways of oxidative damage. Annu. Rev. Microbiol. 2003, 57, 395–418. [Google Scholar] [CrossRef] [PubMed]
- Pinto, E.; Sigaud-Kutner, T.C.S.; Leitao, M.A.S.; Okamoto, O.K.; Morse, D.; Colepicolo, P. Heavy metal-induced oxidative stress in algae. J. Phycol. 2003, 39, 1008–1018. [Google Scholar] [CrossRef]
- Knauert, S.; Knauer, K. The role of reactive oxygen species in copper toxicity to two freshwater green algae1. J. Phycol. 2008, 44, 311–319. [Google Scholar] [CrossRef]
- Morelli, E.; Scarano, G. Copper-induced changes of non-protein thiols and antioxidant enzymes in the marine microalga Phaeodactylum tricomutum. Plant Sci. 2004, 167, 289–296. [Google Scholar] [CrossRef]
- Sabatini, S.E.; Juarez, A.B.; Eppis, M.R.; Bianchi, L.; Luquet, C.M.; de Molina, M.D.R. Oxidative stress and antioxidant defenses in two green microalgae exposed to copper. Ecotoxicol. Environ. Saf. 2009, 72, 1200–1206. [Google Scholar] [CrossRef]
- Wang, L.; Huang, X.L.; Sun, W.L.; Too, H.Z.; Laserna, A.K.C.; Li, S.F.Y. A global metabolomic insight into the oxidative stress and membrane damage of copper oxide nanoparticles and microparticles on microalga Chlorella vulgaris. Environ. Pollut. 2020, 258, 113647. [Google Scholar] [CrossRef]
- Kwon, D.H.; Cha, H.-J.; Lee, H.; Hong, S.-H.; Park, C.; Park, S.-H.; Kim, G.-Y.; Kim, S.; Kim, H.-S.; Hwang, H.-J.; et al. Protective Effect of Glutathione against Oxidative Stress-induced Cytotoxicity in RAW 264.7 Macrophages through Activating the Nuclear Factor Erythroid 2-Related Factor-2/Heme Oxygenase-1 Pathway. Antioxidants 2019, 8, 82. [Google Scholar] [CrossRef]
- Meister, A.; Anderson, M.E. Glutathione. Annu. Rev. Biochem. 1983, 52, 711–760. [Google Scholar] [CrossRef]
- Ahner, B.A.; Wei, L.P.; Oleson, J.R.; Ogura, N. Glutathione and other low molecular weight thiols in marine phytoplankton under metal stress. Mar. Ecol.-Prog. Ser. 2002, 232, 93–103. [Google Scholar] [CrossRef]
- Leal, M.F.C.; Vasconcelos, M.; van den Berg, C.M.G. Copper-induced release of complexing ligands similar to thiols by Emiliania huxleyi in seawater cultures. Limnol. Oceanogr. 1999, 44, 1750–1762. [Google Scholar] [CrossRef]
- Musgrave, W.B.; Yi, H.; Kline, D.; Cameron, J.C.; Wignes, J.; Dey, S.; Pakrasi, H.B.; Jez, J.M. Probing the origins of glutathione biosynthesis through biochemical analysis of glutamate-cysteine ligase and glutathione synthetase from a model photosynthetic prokaryote. Biochem. J. 2013, 450, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Cobbett, C.; Goldsbrough, P. Phytochelatins and metallothioneins: Roles in Heavy Metal Detoxification and Homeostasis. Annu. Rev. Plant Biol. 2002, 53, 159–182. [Google Scholar] [CrossRef]
- Hirata, K.; Tsuji, N.; Miyamoto, K. Biosynthetic regulation of phytochelatins, heavy metal-binding peptides. J. Biosci. Bioeng. 2005, 100, 593–599. [Google Scholar] [CrossRef]
- Howe, G.; Merchant, S. Heavy metal-activated synthesis of peptides in Chlamydomonas reinhardtii. Plant Physiol. 1992, 98, 127–136. [Google Scholar] [CrossRef]
- Peng, K.-T.; Zheng, C.-N.; Xue, J.; Chen, X.-Y.; Yang, W.-D.; Liu, J.-S.; Bai, W.; Li, H.-Y. Delta 5 Fatty Acid Desaturase Upregulates the Synthesis of Polyunsaturated Fatty Acids in the Marine Diatom Phaeodactylum tricornutum. J. Agric. Food Chem. 2014, 62, 8773–8776. [Google Scholar] [CrossRef]
- Aviles, C.; Loza-Tavera, H.; Terry, N.; Moreno-Sanchez, R. Mercury pretreatment selects an enhanced cadmium-accumulating phenotype in Euglena gracilis. Arch. Microbiol. 2003, 180, 1–10. [Google Scholar] [CrossRef]
- Mendoza-Cozatl, D.; Devars, S.; Loza-Tavera, H.; Moreno-Sanchez, R. Cadmium accumulation in the chloroplast of Euglena gracilis. Physiol. Plant. 2002, 115, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Balzano, S.; Sardo, A.; Blasio, M.; Chahine, T.B.; Dell’Anno, F.; Sansone, C.; Brunet, C. Microalgal Metallothioneins and Phytochelatins and Their Potential Use in Bioremediation. Front. Microbiol. 2020, 11, 517. [Google Scholar] [CrossRef] [PubMed]
- Ahner, B.A.; Kong, S.; Morel, F.M.M. Phytochelatin production in marine algae. 1. An interspecific comparison. Limnol. Oceanogr. 1995, 40, 649–657. [Google Scholar] [CrossRef]
- Ahner, B.A.; Morel, F.M.M. Phytochelatin production in marine algae. 2. Induction by various metals. Limnol. Oceanogr. 1995, 40, 658–665. [Google Scholar] [CrossRef]
- Tsuji, N.; Hirayanagi, N.; Iwabe, O.; Namba, T.; Tagawa, M.; Miyamoto, S.; Miyasaka, H.; Takagi, M.; Hirata, K.; Miyamoto, K. Regulation of phytochelatin synthesis by zinc and cadmium in marine green alga, Dunaliella tertiolecta. Phytochemistry 2003, 62, 453–459. [Google Scholar] [CrossRef]
- Kalinowska, R.; Pawlik-Skowrońska, B. Response of two terrestrial green microalgae (Chlorophyta, Trebouxiophyceae) isolated from Cu-rich and unpolluted soils to copper stress. Environ. Pollut. 2010, 158, 2778–2785. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Tan, N.G.; Fu, B.; Li, S.F. Metallomics and NMR-based metabolomics of Chlorella sp. reveal the synergistic role of copper and cadmium in multi-metal toxicity and oxidative stress. Met. Integr. Biomet. Sci. 2015, 7, 426–438. [Google Scholar] [CrossRef]
- Ziller, A.; Fraissinet-Tachet, L. Metallothionein diversity and distribution in the tree of life: A multifunctional protein. Met. Integr. Biomet. Sci. 2018, 10, 1549–1559. [Google Scholar] [CrossRef]
- Leszczyszyn, O.I.; Imam, H.T.; Blindauer, C.A. Diversity and distribution of plant metallothioneins: A review of structure, properties and functions. Met. Integr. Biomet. Sci. 2013, 5, 1146–1169. [Google Scholar] [CrossRef]
- Capdevila, M.; Atrian, S. Metallothionein protein evolution: A miniassay. J. Biol. Inorg. Chem. 2011, 16, 977–989. [Google Scholar] [CrossRef]
- Kumar, K.S.; Dahms, H.U.; Won, E.J.; Lee, J.S.; Shin, K.H. Microalgae—A promising tool for heavy metal remediation. Ecotoxicol. Environ. Saf. 2015, 113, 329–352. [Google Scholar] [CrossRef]
- Ziller, A.; Yadav, R.K.; Capdevila, M.; Reddy, M.S.; Vallon, L.; Marmeisse, R.; Atrian, S.; Palacios, O.; Fraissinet-Tachet, L. Metagenomics analysis reveals a new metallothionein family: Sequence and metal-binding features of new environmental cysteine-rich proteins. J. Inorg. Biochem. 2017, 167, 1–11. [Google Scholar] [CrossRef]
- Gutierrez, J.C.; Amaro, F.; Diaz, S.; de Francisco, P.; Cubas, L.L.; Martin-Gonzalez, A. Ciliate metallothioneins: Unique microbial eukaryotic heavy-metal-binder molecules. J. Biol. Inorg. Chem. 2011, 16, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.-C.; de Francisco, P.; Amaro, F.; Díaz, S.; Martín-González, A. Chapter 22—Structural and Functional Diversity of Microbial Metallothionein Genes. In Microbial Diversity in the Genomic Era; Das, S., Dash, H.R., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 387–407. [Google Scholar]
- Zahid, M.T.; Shakoori, F.R.; Zulfiqar, S.; Al-Ghanim, K.A.; Shakoori, A.R. Growth Characteristics, Metal Uptake and Expression Analysis of Copper Metallothionein in a Newly Reported Ciliate, Tetrahymena farahensis. Pak. J. Zool. 2018, 50, 1171–1181. [Google Scholar] [CrossRef]
- Balzano, S.; Sardo, A. Bioinformatic prediction of putative metallothioneins in non-ciliate protists. Biol. Lett. 2022, 18, 20220039. [Google Scholar] [CrossRef]
- Ruttkay-Nedecky, B.; Nejdl, L.; Gumulec, J.; Zitka, O.; Masarik, M.; Eckschlager, T.; Stiborova, M.; Adam, V.; Kizek, R. The Role of Metallothionein in Oxidative Stress. Int. J. Mol. Sci. 2013, 14, 6044–6066. [Google Scholar] [CrossRef]
- Chen, Z.Z.; Zhu, L.; Wilkinson, K.J. Validation of the Biotic Ligand Model in Metal Mixtures: Bioaccumulation of Lead and Copper. Environ. Sci. Technol. 2010, 44, 3580–3586. [Google Scholar] [CrossRef]
- Sanchez-Marin, P.; Fortin, C.; Campbell, P.G.C. Lead (Pb) and copper (Cu) share a common uptake transporter in the unicellular alga Chlamydomonas reinhardtii. Biometals 2014, 27, 173–181. [Google Scholar] [CrossRef]
- Huang, Z.Q.; Chen, B.Y.; Zhang, J.; Yang, C.L.; Wang, J.; Song, F.; Li, S.G. Absorption and speciation of arsenic by microalgae under arsenic-copper Co-exposure. Ecotoxicol. Environ. Saf. 2021, 213, 112024. [Google Scholar] [CrossRef]
- Zhu, X.; Zhao, W.; Chen, X.; Zhao, T.; Tan, L.; Wang, J. Growth inhibition of the microalgae Skeletonema costatum under copper nanoparticles with microplastic exposure. Mar. Environ. Res 2020, 158, 105005. [Google Scholar] [CrossRef]
- Wan, J.K.; Chu, W.L.; Kok, Y.Y.; Lee, C.S. Influence of polystyrene microplastic and nanoplastic on copper toxicity in two freshwater microalgae. Environ. Sci. Pollut. Res. Int. 2021, 28, 33649–33668. [Google Scholar] [CrossRef]
- Chen, B.Y.; Lee, C.H.; Hsueh, C.C. Dose-response assessment upon CO2 tolerance of indigenous microalgal isolates for biofuel production. In Proceedings of the 6th International Conference on Applied Energy (ICAE), Taipei, Taiwan, 30 May–2 June 2014; pp. 1047–1057. [Google Scholar]
- Nagase, H.; Yoshihara, K.-i.; Eguchi, K.; Okamoto, Y.; Murasaki, S.; Yamashita, R.; Hirata, K.; Miyamoto, K. Uptake pathway and continuous removal of nitric oxide from flue gas using microalgae. Biochem. Eng. J. 2001, 7, 241–246. [Google Scholar] [CrossRef]
- Mojiri, A.; Baharlooeian, M.; Zahed, M.A. The Potential of Chaetoceros muelleri in Bioremediation of Antibiotics: Performance and Optimization. Int. J. Environ. Res. Public Health 2021, 18, 977. [Google Scholar] [CrossRef] [PubMed]
- Albert, K.; Hsieh, P.Y.; Chen, T.H.; Hou, C.H.; Hsu, H.Y. Diatom-assisted biomicroreactor targeting the complete removal of perfluorinated compounds. J. Hazard. Mater. 2020, 384, 121491. [Google Scholar] [CrossRef] [PubMed]
- Minggat, E.; Roseli, W.; Tanaka, Y. Nutrient Absorption and Biomass Production by the Marine Diatom Chaetoceros muelleri: Effects of Temperature, Salinity, Photoperiod, and Light Intensity. J. Ecol. Eng. 2021, 22, 231–240. [Google Scholar] [CrossRef]
- Gao, F.; Li, C.; Yang, Z.H.; Zeng, G.M.; Mu, J.; Liu, M.; Cui, W. Removal of nutrients, organic matter, and metal from domestic secondary effluent through microalgae cultivation in a membrane photobioreactor. J. Chem. Technol. Biotechnol. 2016, 91, 2713–2719. [Google Scholar] [CrossRef]
- Scarponi, P.; Ghirardini, A.M.V.; Bravi, M.; Cavinato, C. Evaluation of Chlorella vulgaris and Scenedesmus obliquus growth on pretreated organic solid waste digestate. Waste Manag. 2021, 119, 235–241. [Google Scholar] [CrossRef]
- Stauber, J.L.; Davies, C.M. Use and limitations of microbial bioassays for assessing copper bioavailability in the aquatic environment. Environ. Rev. 2000, 8, 255–301. [Google Scholar] [CrossRef]
- Wang, L.; Zheng, B. Toxic effects of fluoranthene and copper on marine diatom Phaeodactylum tricornutum. J. Environ. Sci. 2008, 20, 1363–1372. [Google Scholar] [CrossRef]
- Oliveira, R.C.; Palmieri, M.C.; Garcia, O. Biosorption of Metals: State of the Art, General Features, and Potential Applications for Environmental and Technological Processes. In Progress in Biomass and Bioenergy Production; IntechOpen: London, UK, 2011; pp. 151–176. [Google Scholar]
- Vijayaraghavan, K.; Yun, Y.S. Bacterial biosorbents and biosorption. Biotechnol. Adv. 2008, 26, 266–291. [Google Scholar] [CrossRef]
- Wang, J.L.; Chen, C. Biosorbents for heavy metals removal and their future. Biotechnol. Adv. 2009, 27, 195–226. [Google Scholar] [CrossRef]
- Zhang, B.; Li, W.; Guo, Y.; Zhang, Z.; Shi, W.; Cui, F.; Lens, P.N.L.; Tay, J.-H. Microalgal-bacterial consortia: From interspecies interactions to biotechnological applications. Renew. Sustain. Energy Rev. 2020, 118, 109563. [Google Scholar] [CrossRef]
- Fallahi, A.; Rezvani, F.; Asgharnejad, H.; Khorshidi Nazloo, E.; Hajinajaf, N.; Higgins, B. Interactions of microalgae-bacteria consortia for nutrient removal from wastewater: A review. Chemosphere 2021, 272, 129878. [Google Scholar] [CrossRef]
- Ji, X.; Jiang, M.; Zhang, J.; Jiang, X.; Zheng, Z. The interactions of algae-bacteria symbiotic system and its effects on nutrients removal from synthetic wastewater. Bioresour. Technol. 2018, 247, 44–50. [Google Scholar] [CrossRef]
- Higgins, B.T.; Gennity, I.; Samra, S.; Kind, T.; Fiehn, O.; VanderGheynst, J.S. Cofactor symbiosis for enhanced algal growth, biofuel production, and wastewater treatment. Algal Res. 2016, 17, 308–315. [Google Scholar] [CrossRef] [Green Version]
- Perera, I.A.; Abinandan, S.; Panneerselvan, L.; Subashchandrabose, S.R.; Venkateswarlu, K.; Naidu, R.; Megharaj, M. Co-culturing of microalgae and bacteria in real wastewaters alters indigenous bacterial communities enhancing effluent bioremediation. Algal Res. 2022, 64, 102705. [Google Scholar] [CrossRef]
- Marín, D.; Posadas, E.; Cano, P.; Pérez, V.; Lebrero, R.; Muñoz, R. Influence of the seasonal variation of environmental conditions on biogas upgrading in an outdoors pilot scale high rate algal pond. Bioresour. Technol. 2018, 255, 354–358. [Google Scholar] [CrossRef]
- Loutseti, S.; Danielidis, D.B.; Economou-Amilli, A.; Katsaros, C.; Santas, R.; Santas, P. The application of a micro-algal/bacterial biofilter for the detoxification of copper and cadmium metal wastes. Bioresour. Technol. 2009, 100, 2099–2105. [Google Scholar] [CrossRef]
- Mubashar, M.; Naveed, M.; Mustafa, A.; Ashraf, S.; Shehzad Baig, K.; Alamri, S.; Siddiqui, M.H.; Zabochnicka-Świątek, M.; Szota, M.; Kalaji, H.M. Experimental Investigation of Chlorella vulgaris and Enterobacter sp. MN17 for Decolorization and Removal of Heavy Metals from Textile Wastewater. Water 2020, 12, 3034. [Google Scholar] [CrossRef]
- Zuñiga, C.; Li, T.; Guarnieri, M.T.; Jenkins, J.P.; Li, C.-T.; Bingol, K.; Kim, Y.-M.; Betenbaugh, M.J.; Zengler, K. Synthetic microbial communities of heterotrophs and phototrophs facilitate sustainable growth. Nat. Commun. 2020, 11, 3803. [Google Scholar] [CrossRef]
- Palacios, O.A.; López, B.R.; de-Bashan, L.E. Microalga Growth-Promoting Bacteria (MGPB): A formal term proposed for beneficial bacteria involved in microalgal–bacterial interactions. Algal Res. 2022, 61, 102585. [Google Scholar] [CrossRef]
- Yu, Q.; Li, P.; Li, B.; Zhang, C.; Zhang, C.; Ge, Y. Effects of algal–bacterial ratio on the growth and cadmium accumulation of Chlorella salina–Bacillus subtilis consortia. J. Basic Microbiol. 2022, 62, 518–529. [Google Scholar] [CrossRef]
- Li, Y.; Yang, X.; Geng, B. Preparation of Immobilized Sulfate-Reducing Bacteria-Microalgae Beads for Effective Bioremediation of Copper-Containing Wastewater. Water Air Soil Pollut. 2018, 229, 54. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavalletti, E.; Romano, G.; Palma Esposito, F.; Barra, L.; Chiaiese, P.; Balzano, S.; Sardo, A. Copper Effect on Microalgae: Toxicity and Bioremediation Strategies. Toxics 2022, 10, 527. https://doi.org/10.3390/toxics10090527
Cavalletti E, Romano G, Palma Esposito F, Barra L, Chiaiese P, Balzano S, Sardo A. Copper Effect on Microalgae: Toxicity and Bioremediation Strategies. Toxics. 2022; 10(9):527. https://doi.org/10.3390/toxics10090527
Chicago/Turabian StyleCavalletti, Elena, Giovanna Romano, Fortunato Palma Esposito, Lucia Barra, Pasquale Chiaiese, Sergio Balzano, and Angela Sardo. 2022. "Copper Effect on Microalgae: Toxicity and Bioremediation Strategies" Toxics 10, no. 9: 527. https://doi.org/10.3390/toxics10090527
APA StyleCavalletti, E., Romano, G., Palma Esposito, F., Barra, L., Chiaiese, P., Balzano, S., & Sardo, A. (2022). Copper Effect on Microalgae: Toxicity and Bioremediation Strategies. Toxics, 10(9), 527. https://doi.org/10.3390/toxics10090527