
Developmental Toxic Effects of Thiram on Developing Zebrafish (Danio rerio) Embryos

 ,
,  ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Zebrafish Maintenance and Embryo Collection
2.3. Toxicity Assay
2.4. Deformities Scoring and Calculation
2.5. Heart Rate Analysis
2.6. Body Length Survey
2.7. Spine Deformities Scoring
2.8. Data Analysis
3. Results
3.1. Acute Toxicity of Thiram
3.2. Developmental Toxicity of Thiram
3.3. Effects of Thiram on ZF Cardiac Development and Function
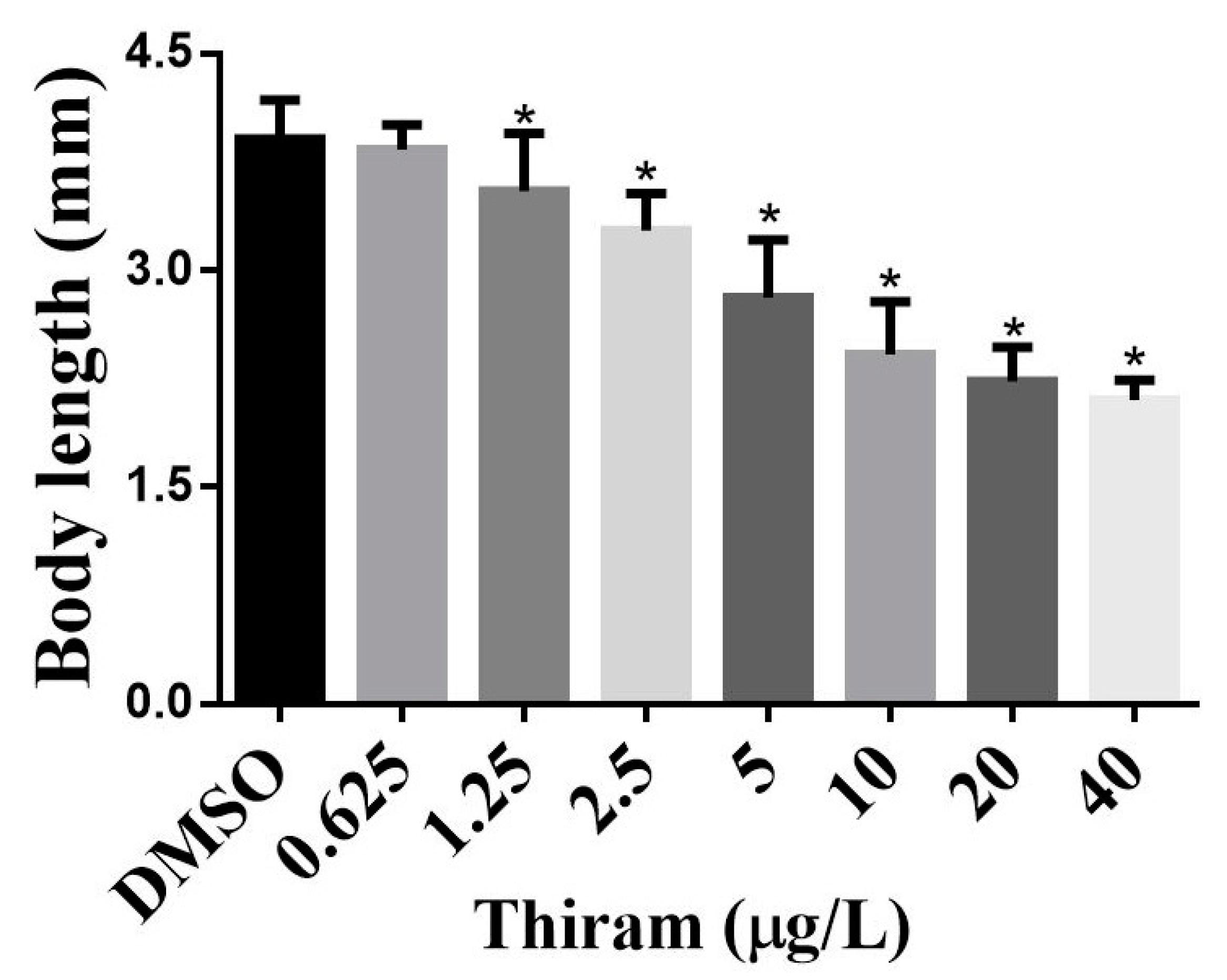
3.4. Effects of Thiram on Overall ZF Growth
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zubrod, J.P.; Bundschuh, M.; Arts, G.; Brühl, C.A.; Imfeld, G.; Knäbel, A.; Payraudeau, S.; Rasmussen, J.J.; Rohr, J.; Scharmüller, A.; et al. Fungicides: An overlooked pesticide class? Environ. Sci. Technol. 2019, 53, 3347–3365. [Google Scholar] [CrossRef] [PubMed]
- US EPA. Reregistration Eligibility Decision Facts, Thiram. 2004. Available online: https://www3.epa.gov/pesticides/chem_search/reg_actions/reregistration/fs_PC-079801_1-Sep-04.pdf (accessed on 15 January 2022).
- Van Boxtel, A.L.; Pieterse, B.; Cenijn, P.; Kamstra, J.H.; Brouwer, A.; van Wieringen, W.; de Boer, J.; Legler, J. Dithiocarbamates induce craniofacial abnormalities and downregulate sox9a during zebrafish development. Toxicol. Sci. 2010, 117, 209–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robens, J.F.J.T. Teratologic studies of carbaryl, diazinon, norea, disulfiram, and thiram in small laboratory animals. Toxicol. Appl. Pharmacol. 1969, 15, 152–163. [Google Scholar] [CrossRef]
- Park, H.; You, H.H.; Song, G. Multiple toxicity of propineb in developing zebrafish embryos: Neurotoxicity, vascular toxicity, and notochord defects in normal vertebrate development. Cop. Biochem. Physiol. C. Toxicol. Pharmacol. 2021, 243, 108993. [Google Scholar] [CrossRef] [PubMed]
- Nageswara, R.; Srinivasa, N.; Jagadish, N.M. Thiram a fungicide induced toxicity on glycogen and blood glucose level of freshwater fish Cyprinus carpio (Hamilton). Int. J. Fish. Aquat. 2017, 5, 93–96. [Google Scholar]
- Heikkila, R.E.; Cabbat, F.S.; Cohen, G. In vivo inhibition of superoxide dismutase in mice by diethyldithiocarbamate. J. Biol. Chem. 1976, 251, 2182–2185. [Google Scholar] [CrossRef]
- Cao, F.; Souders II, C.L.; Li, P.; Pang, S.; Liang, X.; Qiu, L.; Martyniuk, C.J. Developmental neurotoxicity of maneb: Notochord defects, mitochondrial dysfunction and hypoactivity in zebrafish (Danio rerio) embryos and larvae. Ecotoxicol. Environ. Saf. 2019, 170, 227–237. [Google Scholar] [CrossRef]
- Tilton, F.; La Du, J.K.; Vue, M.; Alzarban, N.; Tanguay, R.L. Dithiocarbamates have a common toxic effect on zebrafish body axis formation. Toxicol. Appl. Pharmacol. 2006, 216, 55–68. [Google Scholar] [CrossRef]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Occupational Exposures in Insecticide Application, and Some Pesticides. Lyon (FR): International Agency for Research on Cancer. (IARC Monographs on the Evaluation of Carcinogenic Risks to Humans, No. 53.) Thiram. 1991. Available online: https://www.ncbi.nlm.nih.gov/books/NBK499628/. (accessed on 20 March 2022).
- USGS. Estimated Annual Agricultural Pesticide Use, Pesticide Use Maps—Thiram. Available online: https://water.usgs.gov/nawqa/pnsp/usage/maps/show_map.php?year=2017&map=THIRAM&hilo=L&disp=Thiram (accessed on 20 January 2022).
- Imports of technicals for agrochemicals and raw materials for technical. In Agrochemical Yearbook; Korean Crop Protection Association: Seoul, Korea, 2018; pp. 402–403.
- Maznah, Z.; Halimah, M.; Ismail, B.S. Evaluation of the persistence and leaching behaviour of thiram fungicide in soil, water and oil palm leaves. Bull Environ. Contam. Toxicol. 2018, 100, 677–682. [Google Scholar] [CrossRef]
- Kaskevich, L.; Bezuglyĭ, V.P. Clinical aspects of chronic intoxications induced by TMTD. Vrachebnoe Delo 1973, 6, 128–130. [Google Scholar]
- Cherpak, V.; Bezuglyĭ, V.; Kaskevich, L.J.V.D. Health and hygienic characteristics of the working conditions and state of health of persons working with tetramethylthiuramdisulfide (TMTD). Vrachebnoe Delo 1971, 10, 136–139. [Google Scholar] [PubMed]
- Hayes, W.J.; Laws, E.R. Handbook of Pesticide Toxicology; Academic Press: Cambridge, MA, USA, 1991. [Google Scholar]
- Matthiaschk, G.J.A.T. The influence of L-cysteine on the teratogenicity caused by thiram (TMTD) in MMRI mice. Arch. Toxicol. 1973, 30, 251. [Google Scholar] [CrossRef]
- Liu, K.; Li, Y.; Iqbal, M.; Tang, Z.; Zhang, H. Thiram exposure in environment: A critical review on cytotoxicity. Chemosphere 2022, 295, 133928. [Google Scholar] [CrossRef] [PubMed]
- Pesticide Properties DataBase, Thiram (Ref: ENT 987). Available online: http://sitem.herts.ac.uk/aeru/ppdb/en/Reports/642.htm (accessed on 20 December 2021).
- Belaid, C.; Sbartai, I. Assessing the effects of Thiram to oxidative stress responses in a freshwater bioindicator cladoceran (Daphnia magna). Chemosphere 2021, 268, 128808. [Google Scholar] [CrossRef]
- Chen, X.; Fang, M.; Chernick, M.; Wang, F.; Yang, J.; Yu, Y.; Zheng, N.; Teraoka, H.; Nanba, S.; Hiraga, T.; et al. The case for thyroid disruption in early life stage exposures to thiram in zebrafish (Danio rerio). Gen. Comp. Endocrinol. 2019, 271, 73–81. [Google Scholar] [CrossRef]
- Bailey, J.; Oliveri, A.; Levin, E.D. Zebrafish model systems for developmental neurobehavioral toxicology. Birth Defects Res. C. Embryo Today 2013, 99, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Vasamsetti, B.M.K.; Chon, K.; Kim, J.; Oh, J.-A.; Yoon, C.-Y.; Park, H.-H. Transcriptome-Based Identification of Genes Responding to the Organophosphate Pesticide Phosmet in Danio rerio. Genes 2021, 12, 1738. [Google Scholar] [CrossRef]
- Horzmann, K.A.; Freeman, J.L. Making waves: New developments in toxicology with the zebrafish. Toxicol. Sci. 2018, 163, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Glaberman, S.; Padilla, S.; Barron, M.G. Evaluating the zebrafish embryo toxicity test for pesticide hazard screening. Environ. Toxicol. Chem. 2017, 36, 1221–1226. [Google Scholar] [CrossRef]
- Haque, E.; Ward, A.C. Zebrafish as a model to evaluate nanoparticle toxicity. Nanomaterials 2018, 8, 561. [Google Scholar] [CrossRef] [Green Version]
- Cassar, S.; Adatto, I.; Freeman, J.L.; Gamse, J.T.; Iturria, I.; Lawrence, C.; Muriana, A.; Peterson, R.T.; Van Cruchten, S.; Zon, L.I. Use of zebrafish in drug discovery toxicology. Chem. Res. Toxicol. 2019, 33, 95–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasamsetti, B.M.K.; Kim, N.S.; Chon, K.; Park, H.-H. Developmental toxic effects of phosmet on zebrafish (Danio rerio) embryos. Korean J. Pestic. Sci. 2020, 24, 343–351. [Google Scholar] [CrossRef]
- Vasamsetti, B.M.K.; Kim, N.-S.; Chon, K.; Park, H.-H. Teratogenic and developmental toxic effects of etridiazole on zebrafish (Danio rerio) embryos. Appl. Biol. Chem. 2020, 63, 80. [Google Scholar] [CrossRef]
- OECD. Test No. 236: Fish Embryo Acute Toxicity (FET) Test; OECD: Paris, France, 2013. [Google Scholar]
- Cao, F.; Souders II, C.L.; Li, P.; Adamovsky, O.; Pang, S.; Qiu, L.; Martyniuk, C.J. Developmental toxicity of the fungicide ziram in zebrafish (Danio rerio). Chemosphere 2019, 214, 303–313. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Sant, K.E.; Timme-Laragy, A.R. Zebrafish as a model for toxicological perturbation of yolk and nutrition in the early embryo. Curr. Environ. Health Rep. 2018, 5, 125–133. [Google Scholar] [CrossRef]
- Helvik, J.V.; Walther, B.T. Development of hatchability in halibut (Hippoglossus hippoglossus) embryos. Int. J. Dev. Biol. 2002, 37, 487–490. [Google Scholar]
- Trikić, M.Z.; Monk, P.; Roehl, H.; Partridge, L.J. Regulation of zebrafish hatching by tetraspanin cd63. PLoS ONE 2011, 6, e19683. [Google Scholar] [CrossRef] [Green Version]
- Sogard, S. Size-selective mortality in the juvenile stage of teleost fishes: A review. Bull. Mar. Sci. 1997, 60, 1129–1157. [Google Scholar]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef] [Green Version]
- Lulla, A.; Barnhill, L.; Bitan, G.; Ivanova, M.I.; Nguyen, B.; O’Donnell, K.; Stahl, M.C.; Yamashiro, C.; Klärner, F.-G.; Schrader, T.; et al. Neurotoxicity of the Parkinson disease-associated pesticide ziram is synuclein-dependent in zebrafish embryos. Environ. Health Perspect. 2016, 124, 1766–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, D.R.; Samsa, L.A.; Qian, L.; Liu, J. Advances in the study of heart development and disease using zebrafish. J. Cardiovasc. Dev. Dis. 2016, 3, 13. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Thiram (μg/L) | Mortality (%) | No Tail Detachment (%) | Abnormal Somites (%) | Abnormal Eye Pigmentation (%) | Abnormal Tail Morphology (%) | Abnormal Tail Blood Flow (%) | |
| 24 hpf | 0 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.83 ± 2.04 | |||
| 0.625 | 0.83 ± 2.04 | 0.00 ± 0.00 | 0.00 ± 0.00 | ||||
| 1.25 | 4.17 ± 4.92 | 5.27 ± 3.34 | 11.01 ± 13.14 | ||||
| 2.5 | 5.83 ± 4.92 | 0.88 ± 2.15 | 43.69 ± 10.18 | ||||
| 5 | 2.50 ± 2.74 | 0.83 ± 2.04 | 70.26 ± 29.09 | ||||
| 10 | 12.50 ± 11.73 | 1.28 ± 3.14 | 69.87 ± 26.20 | ||||
| 20 | 16.67 ± 6.06 | 1.91 ± 2.96 | 88.41 ± 9.98 | ||||
| 40 | 19.17 ± 9.70 | 30.70 ± 15.46 | 93.52 ± 15.58 | ||||
| 48 hpf | 0 | 1.67 ± 2.58 | 0.83 ± 2.04 | 0.00 ± 0.00 | 0.00 ± 0.00 | ||
| 0.625 | 0.83 ± 2.04 | 0.83 ± 2.04 | 0.00 ± 0.00 | 1.67 ± 4.08 | |||
| 1.25 | 8.33 ± 2.04 | 5.36 ± 4.71 | 2.68 ± 2.94 | 8.50 ± 8.88 | |||
| 2.5 | 7.50 ± 5.24 | 29.28 ± 15.13 | 9.34 ± 8.85 | 33.09 ± 22.58 | |||
| 5 | 7.50 ± 6.12 | 38.52 ± 16.28 | 21.16 ± 15.32 | 46.85 ± 18.56 | |||
| 10 | 20.83 ± 10.21 | 87.29 ± 13.49 | 89.53 ± 4.65 | 92.03 ± 4.75 | |||
| 20 | 45.00 ± 10.49 | 100.00 ± 0.00 | 98.72 ± 3.14 | 98.72 ± 3.14 | |||
| 40 | 60.83 ± 9.17 | 100.00 ± 0.00 | 97.62 ± 5.83 | 97.62 ± 5.83 | |||
| Thiram (μg/L) | Mortality (%) | Unhatched Eggs (%) | Pericardial Edema (%) | Yolk Sac Edema (%) | |||
| 72 hpf | 0 | 1.67 ± 2.58 | 7.63 ± 10.49 | 3.33 ± 4.08 | 2.50 ± 2.74 | ||
| 0.625 | 0.83 ± 2.04 | 3.38 ± 4.11 | 3.38 ± 4.11 | 4.25 ± 5.04 | |||
| 1.25 | 8.33 ± 4.08 | 43.11 ± 25.51 | 14.89 ± 14.89 | 8.34 ± 6.82 | |||
| 2.5 | 8.33 ± 5.16 | 91.06 ± 8.44 | 37.41 ± 29.93 | 22.04 ± 16.55 | |||
| 5 | 10.83 ± 7.36 | 95.83 ± 6.45 | 48.50 ± 31.66 | 53.44 ± 29.18 | |||
| 10 | 26.67 ± 8.76 | 95.83 ± 10.21 | 66.01 ± 20.54 | 77.85 ± 22.52 | |||
| 20 | 58.33 ± 9.83 | 100.00 ± 0.00 | 96.82 ± 4.94 | 98.48 ± 3.71 | |||
| 40 | 76.67 ± 12.11 | 100.00 ± 0.00 | 81.48 ± 29.54 | 100.00 ± 0.00 | |||
| 96 hpf | 0 | 2.50 ± 2.74 | 0.83 ± 2.04 | ||||
| 0.625 | 0.83 ± 2.04 | 0.88 ± 2.15 | |||||
| 1.25 | 8.33 ± 4.08 | 31.90 ± 14.17 | |||||
| 2.5 | 11.67 ± 6.83 | 92.56 ± 9.53 | |||||
| 5 | 23.33 ± 14.72 | 97.22 ± 6.80 | |||||
| 10 | 54.17 ± 8.61 | 96.30 ± 9.07 | |||||
| 20 | 81.67 ± 6.83 | 97.22 ± 6.80 | |||||
| 40 | 95.00 ± 3.16 | 100.00 ± 0.00 | |||||
| Deformity | Time (hpf) | LC50 (μg/L) | EC50 (μg/L) | Teratogenic Index (96 hpf LC50/EC50) |
|---|---|---|---|---|
| Mortality | 48 | 13.10 ± 2.17 | - | - |
| Mortality | 96 | 8.87 ± 2.09 | - | - |
| Abnormal somites | 24 | - | 3.88 ± 1.23 | 2.37 ± 0.46 |
| Reduced eye pigment | 48 | - | 5.04 ± 1.82 | 1.84 ± 0.40 |
| Abnormal tail morphology | 48 | - | 6.23 ± 0.92 | 1.42 ± 0.24 |
| Abnormal tail blood flow | 48 | - | 4.25 ± 1.53 | 2.18 ± 0.44 |
| Unhatched embryos | 72 | - | 1.38 ± 0.24 | 6.66 ± 2.22 |
| Pericardial edema | 72 | - | 6.67 ± 5.55 | 2.08 ± 1.28 |
| Yolk sac edema | 72 | - | 5.24 ± 5.55 | 1.89 ± 0.76 |
| Unhatched embryos | 96 | - | 1.39 ± 0.25 | 6.60 ± 2.10 |
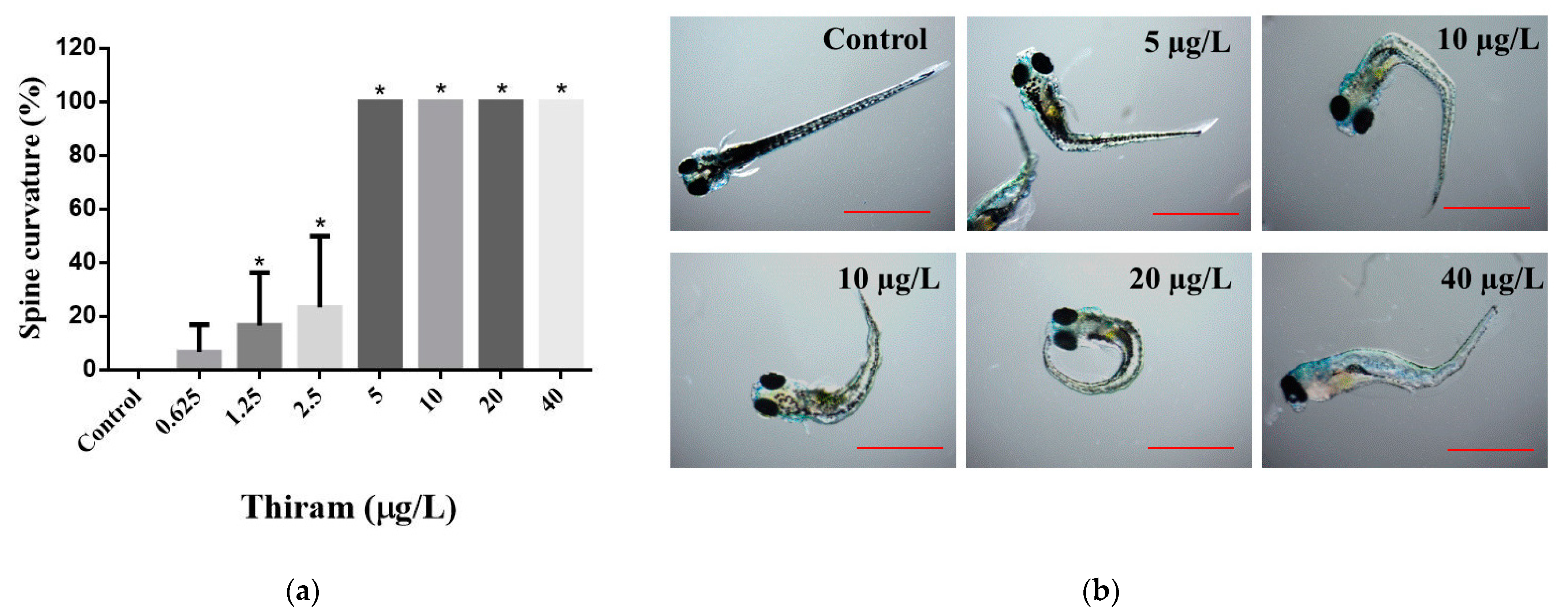
| Spine curvature | 144 | - | 2.60 ± 0.82 | 3.68 ± 1.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasamsetti, B.M.K.; Chon, K.; Kim, J.; Oh, J.-A.; Yoon, C.-Y.; Park, H.-H. Developmental Toxic Effects of Thiram on Developing Zebrafish (Danio rerio) Embryos. Toxics 2022, 10, 369. https://doi.org/10.3390/toxics10070369
Vasamsetti BMK, Chon K, Kim J, Oh J-A, Yoon C-Y, Park H-H. Developmental Toxic Effects of Thiram on Developing Zebrafish (Danio rerio) Embryos. Toxics. 2022; 10(7):369. https://doi.org/10.3390/toxics10070369
Chicago/Turabian StyleVasamsetti, Bala Murali Krishna, Kyongmi Chon, Juyeong Kim, Jin-A Oh, Chang-Young Yoon, and Hong-Hyun Park. 2022. "Developmental Toxic Effects of Thiram on Developing Zebrafish (Danio rerio) Embryos" Toxics 10, no. 7: 369. https://doi.org/10.3390/toxics10070369
APA StyleVasamsetti, B. M. K., Chon, K., Kim, J., Oh, J.-A., Yoon, C.-Y., & Park, H.-H. (2022). Developmental Toxic Effects of Thiram on Developing Zebrafish (Danio rerio) Embryos. Toxics, 10(7), 369. https://doi.org/10.3390/toxics10070369