


Nanoplastics in Aquatic Environments: Impacts on Aquatic Species and Interactions with Environmental Factors and Pollutants


Abstract
:1. Introduction
2. Plastics Breakdown and Small Plastic Particles: MPs and NPs as Ubiquitous Particles
3. Cellular and Molecular Impacts of NPs on Aquatic Species
3.1. The Small Size of NPs as a Key Factor in Their Ecotoxicity
3.2. Main Biological Effects or Modes of Actions Associated with NPs Toxicity
3.3. NPs as Inducers of Oxidative Stress
3.4. NPs as Disruptors of Energy Metabolism
3.5. NPs as Modulators of the Nervous and Immune Systems
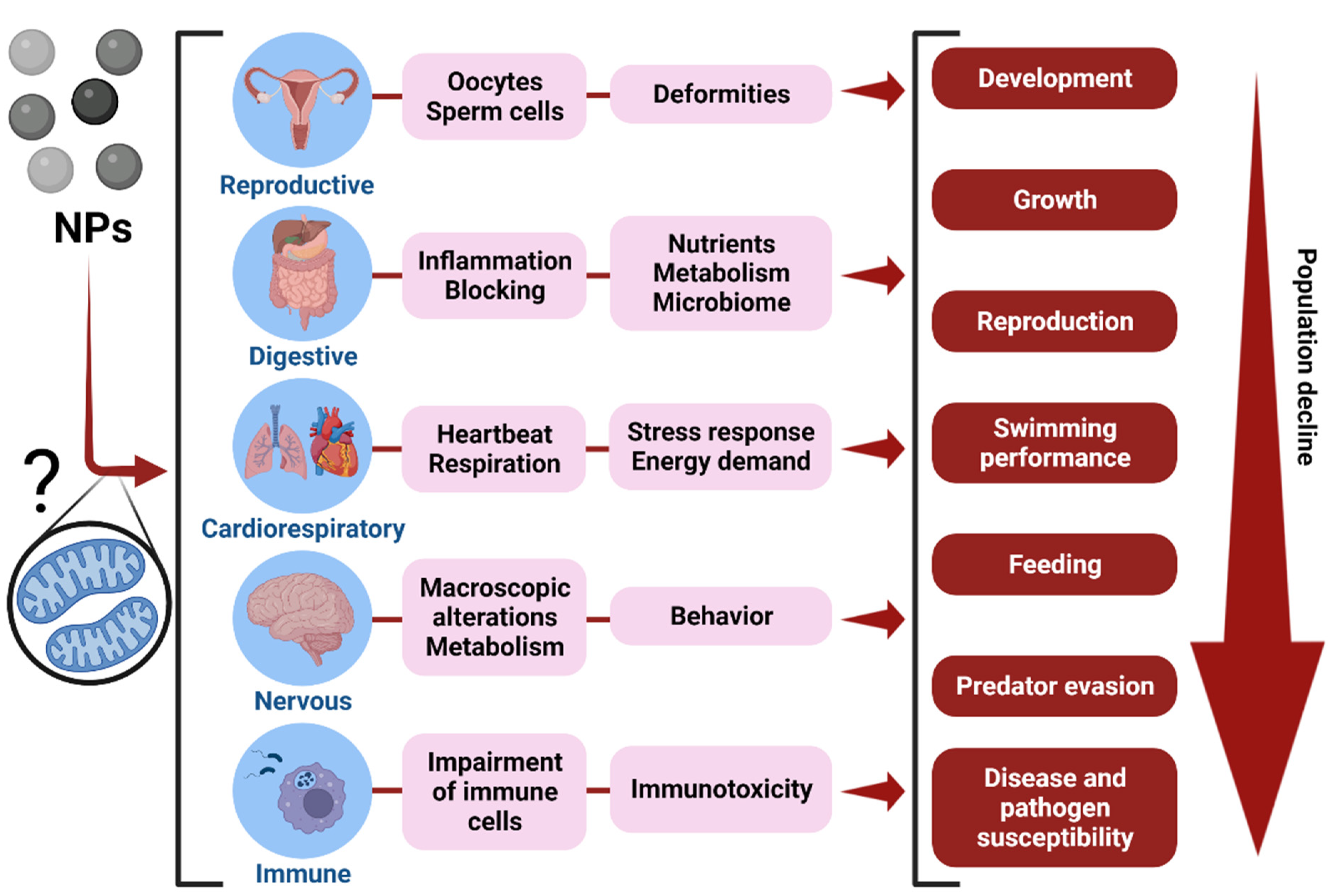
3.6. The Effects of NPs Exposure on Animal Health and Fitness
4. NPs as Carriers of Environmental Pollutants
4.1. Interactions of NPs with Environmental Pollutants and Resulting Toxicity to Aquatic Species
4.1.1. Metals
4.1.2. Polycyclic Aromatic Hydrocarbons (PAHs) and Polychlorinated Biphenyls (PCBs)
4.1.3. Pesticides, Biocides, and Antibiotics
4.1.4. Other Compounds
4.1.5. Complex Environmental Chemical Mixtures
4.2. Interactions of NPs with Microorganisms and Antibiotic-Resistance Genes
4.3. Effects of Aging and Weathering of Plastic Particles on the Carrier Effect
5. Effects of Other Environmental Stressors on NPs’ Toxicity and Interactions with other Environmental Pollutants
5.1. Temperature
5.2. Salinity and pH
5.3. Natural Organic Matter (NOM)
5.4. Food Availability
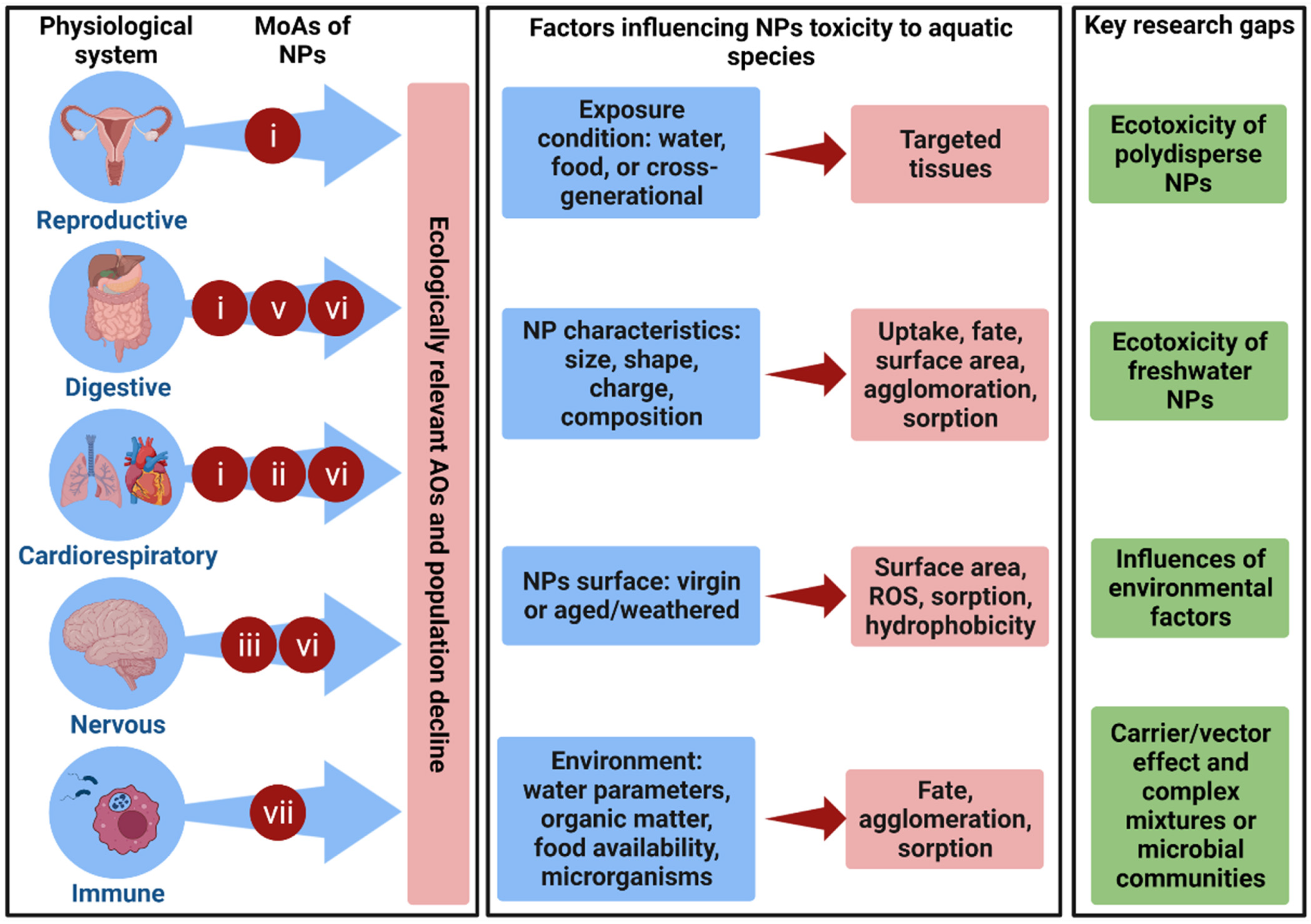
6. Perspectives for Future Research in the Field
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, M.; Gower, R.; Green, J.; Whitebread, E.; Lenkiewicz, Z.; Schröder, P. No Time to Waste: Tackling the Plastic Pollution Crisis before It’s Too Late; Tearfund: Teddington, UK, 2019. [Google Scholar]
- Marine Debris: Understanding, Preventing and Mitigating the Significant Adverse Impacts on Marine and Coastal Biodiversity; Technical Series No. 83; Secretariat of the Convention on Biological Diversity: Montreal, QC, Canada, 2016.
- Kalina, M.; Tilley, E. “This is our next problem”: Cleaning up from the COVID-19 response. Waste Manag. 2020, 108, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.R. Why are we still polluting the marine environment with personal protective equipment? Mar. Pollut. Bull. 2021, 169, 112528. [Google Scholar] [CrossRef] [PubMed]
- Lebreton, L.; Slat, B.; Ferrari, F.; Sainte-Rose, B.; Aitken, J.; Marthouse, R.; Hajbane, S.; Cunsolo, S.; Schwarz, A.; Levivier, A.; et al. Evidence that the Great Pacific Garbage Patch is rapidly accumulating plastic. Sci. Rep. 2018, 8, 4666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijer, L.J.J.; van Emmerik, T.; van der Ent, R.; Schmidt, C.; Lebreton, L. More than 1000 rivers account for 80% of global riverine plastic emissions into the ocean. Sci. Adv. 2021, 7, eaaz5803. [Google Scholar] [CrossRef]
- Lavers, J.L.; Bond, A.L. Exceptional and rapid accumulation of anthropogenic debris on one of the world’s most remote and pristine islands. Proc. Natl. Acad. Sci. USA 2017, 114, 6052–6055. [Google Scholar] [CrossRef] [Green Version]
- Chamas, A.; Moon, H.; Zheng, J.; Qiu, Y.; Tabassum, T.; Jang, J.H.; Abu-Omar, M.; Scott, S.L.; Suh, S. Degradation Rates of Plastics in the Environment. ACS Sustain. Chem. Eng. 2020, 8, 3494–3511. [Google Scholar] [CrossRef] [Green Version]
- Gigault, J.; ter Halle, A.; Baudrimont, M.; Pascal, P.Y.; Gauffre, F.; Phi, T.L.; El Hadri, H.; Grassl, B.; Reynaud, S. Current opinion: What is a nanoplastic? Environ. Pollut. 2018, 235, 1030–1034. [Google Scholar] [CrossRef]
- Jambeck, J.R.; Geyer, R.; Wilcox, C.; Siegler, T.R.; Perryman, M.; Andrady, A.; Narayan, R.; Law, K.L. Plastic waste inputs from land into the ocean. Science 2015, 347, 768–771. [Google Scholar] [CrossRef]
- Erni-Cassola, G.; Zadjelovic, V.; Gibson, M.I.; Christie-Oleza, J.A. Distribution of plastic polymer types in the marine environment; A meta-analysis. J. Hazard. Mater. 2019, 369, 691–698. [Google Scholar] [CrossRef]
- Rostami, S.; Talaie, M.R.; Talaiekhozani, A.; Sillanpää, M. Evaluation of the available strategies to control the emission of microplastics into the aquatic environment. Environ. Sci. Pollut. Res. 2021, 28, 18908–18917. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, A.J.; Brooks, L.S.R.; Reid, W.D.K.; Piertney, S.B.; Narayanaswamy, B.E.; Linley, T.D. Microplastics and synthetic particles ingested by deep-sea amphipods in six of the deepest marine ecosystems on Earth. R. Soc. Open Sci. 2019, 6, 180667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waller, C.L.; Griffiths, H.J.; Waluda, C.M.; Thorpe, S.E.; Loaiza, I.; Moreno, B.; Pacherres, C.O.; Hughes, K.A. Microplastics in the Antarctic marine system: An emerging area of research. Sci. Total Environ. 2017, 598, 220–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cózar, A.; Martí, E.; Duarte, C.M.; García-de-Lomas, J.; van Sebille, E.; Ballatore, T.J.; Eguíluz, V.M.; González-Gordillo, J.I.; Pedrotti, M.L.; Echevarría, F.; et al. The Arctic Ocean as a dead end for floating plastics in the North Atlantic branch of the Thermohaline Circulation. Sci. Adv. 2017, 3, e1600582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, M.C.; Ruf, C.S. Toward the Detection and Imaging of Ocean Microplastics With a Spaceborne Radar. IEEE Trans. Geosci. Remote Sens. 2021, 60, 1–9. [Google Scholar] [CrossRef]
- Burns, E.E.; Boxall, A.B.A. Microplastics in the aquatic environment: Evidence for or against adverse impacts and major knowledge gaps. Environ. Toxicol. Chem. 2018, 37, 2776–2796. [Google Scholar] [CrossRef] [Green Version]
- Avio, C.G.; Gorbi, S.; Regoli, F. Plastics and microplastics in the oceans: From emerging pollutants to emerged threat. Mar. Environ. Res. 2017, 128, 2–11. [Google Scholar] [CrossRef]
- Braun, T.; Ehrlich, L.; Henrich, W.; Koeppel, S.; Lomako, I.; Schwabl, P.; Liebmann, B. Detection of Microplastic in Human Placenta and Meconium in a Clinical Setting. Pharmaceutics 2021, 13, 921. [Google Scholar] [CrossRef]
- Ragusa, A.; Svelato, A.; Santacroce, C.; Catalano, P.; Notarstefano, V.; Carnevali, O.; Papa, F.; Rongioletti, M.C.A.; Baiocco, F.; Draghi, S.; et al. Plasticenta: First evidence of microplastics in human placenta. Environ. Int. 2021, 146, 106274. [Google Scholar] [CrossRef]
- Ter Halle, A.; Jeanneau, L.; Martignac, M.; Jardé, E.; Pedrono, B.; Brach, L.; Gigault, J. Nanoplastic in the North Atlantic Subtropical Gyre. Environ. Sci. Technol. 2017, 51, 13689–13697. [Google Scholar] [CrossRef]
- Allen, D.; Allen, S.; Abbasi, S.; Baker, A.; Bergmann, M.; Brahney, J.; Butler, T.; Duce, R.A.; Eckhardt, S.; Evangeliou, N.; et al. Microplastics and nanoplastics in the marine-atmosphere environment. Nat. Rev. Earth Environ. 2022, 3, 393–405. [Google Scholar] [CrossRef]
- Koelmans, A.A.; Besseling, E.; Shim, W.J. Nanoplastics in the aquatic environment. Critical review. In Marine Anthropogenic Litter; Bergmann, M., Gutow, L., Klages, M., Eds.; Springer: Cham, Switzerland, 2015; pp. 325–340. [Google Scholar] [CrossRef] [Green Version]
- Paul-Pont, I.; Tallec, K.; Gonzalez-Fernandez, C.; Lambert, C.; Vincent, D.; Mazurais, D.; Zambonino-Infante, J.L.; Brotons, G.; Lagarde, F.; Fabioux, C.; et al. Constraints and priorities for conducting experimental exposures of marine organisms to microplastics. Front. Mar. Sci. 2018, 5, 252. [Google Scholar] [CrossRef]
- Pitt, J.A.; Trevisan, R.; Massarsky, A.; Kozal, J.S.; Levin, E.D.; Di Giulio, R.T. Maternal transfer of nanoplastics to offspring in zebrafish (Danio rerio): A case study with nanopolystyrene. Sci. Total Environ. 2018, 643, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Cole, M.; Lindeque, P.; Fileman, E.; Halsband, C.; Goodhead, R.; Moger, J.; Galloway, T.S. Microplastic Ingestion by Zooplankton. Environ. Sci. Technol. 2013, 47, 6646–6655. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-W.; Shim, W.J.; Kwon, O.Y.; Kang, J.-H. Size-Dependent Effects of Micro Polystyrene Particles in the Marine Copepod Tigriopus japonicus. Environ. Sci. Technol. 2013, 47, 11278–11283. [Google Scholar] [CrossRef] [PubMed]
- Van Cauwenberghe, L.; Claessens, M.; Vandegehuchte, M.B.; Janssen, C.R. Microplastics are taken up by mussels (Mytilus edulis) and lugworms (Arenicola marina) living in natural habitats. Environ. Pollut. 2015, 199, 10–17. [Google Scholar] [CrossRef]
- Jeong, J.; Choi, J. Adverse outcome pathways potentially related to hazard identification of microplastics based on toxicity mechanisms. Chemosphere 2019, 231, 249–255. [Google Scholar] [CrossRef]
- Matthews, S.; Mai, L.; Jeong, C.B.; Lee, J.S.; Zeng, E.Y.; Xu, E.G. Key mechanisms of micro- and nanoplastic (MNP) toxicity across taxonomic groups. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 247, 109056. [Google Scholar] [CrossRef]
- González-Acedo, A.; García-Recio, E.; Illescas-Montes, R.; Ramos-Torrecillas, J.; Melguizo-Rodríguez, L.; Costela-Ruiz, V.J. Evidence from in vitro and in vivo studies on the potential health repercussions of micro- and nanoplastics. Chemosphere 2021, 280, 130826. [Google Scholar] [CrossRef]
- Baglioni, P.; Chelazzi, D.; Giorgi, R. Innovative Nanomaterials: Principles, Availability and Scopes. In Nanotechnologies in the Conservation of Cultural Heritage; Springer: Dordrecht, The Netherlands, 2015; pp. 1–14. [Google Scholar] [CrossRef]
- Liu, L.; Fokkink, R.; Koelmans, A.A. Sorption of polycyclic aromatic hydrocarbons to polystyrene nanoplastic. Environ. Toxicol. Chem. 2016, 35, 1650–1655. [Google Scholar] [CrossRef]
- Wright, S.L.; Thompson, R.C.; Galloway, T.S. The physical impacts of microplastics on marine organisms: A review. Environ. Pollut. 2013, 178, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.; Venâncio, C.; Lopes, I.; Oliveira, M. Nanoplastics and marine organisms: What has been studied? Environ. Toxicol. Pharmacol. 2019, 67, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.A.; Kozal, J.S.; Jayasundara, N.; Massarsky, A.; Trevisan, R.; Geitner, N.; Wiesner, M.; Levin, E.D.; Di Giulio, R.T. Uptake, tissue distribution, and toxicity of polystyrene nanoparticles in developing zebrafish (Danio rerio). Aquat. Toxicol. 2018, 194, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Mattsson, K.; Johnson, E.V.; Malmendal, A.; Linse, S.; Hansson, L.A.; Cedervall, T. Brain damage and behavioural disorders in fish induced by plastic nanoparticles delivered through the food chain. Sci. Rep. 2017, 7, 11452. [Google Scholar] [CrossRef] [PubMed]
- Rubio, L.; Marcos, R.; Hernández, A. Potential adverse health effects of ingested micro- and nanoplastics on humans. Lessons learned from in vivo and in vitro mammalian models. J. Toxicol. Environ. Health Part B 2020, 23, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Bouwmeester, H.; Hollman, P.C.H.; Peters, R.J.B. Potential Health Impact of Environmentally Released Micro- and Nanoplastics in the Human Food Production Chain: Experiences from Nanotoxicology. Environ. Sci. Technol. 2015, 49, 8932–8947. [Google Scholar] [CrossRef]
- Cedervall, T.; Hansson, L.-A.; Lard, M.; Frohm, B.; Linse, S. Food Chain Transport of Nanoparticles Affects Behaviour and Fat Metabolism in Fish. PLoS ONE 2012, 7, e32254. [Google Scholar] [CrossRef] [Green Version]
- Jacob, H.; Besson, M.; Swarzenski, P.W.; Lecchini, D.; Metian, M. Effects of Virgin Micro- and Nanoplastics on Fish: Trends, Meta-Analysis, and Perspectives. Environ. Sci. Technol. 2020, 54, 4733–4745. [Google Scholar] [CrossRef]
- Jeong, C.-B.; Kang, H.-M.; Lee, M.-C.; Kim, D.-H.; Han, J.; Hwang, D.-S.; Souissi, S.; Lee, S.-J.; Shin, K.-H.; Park, H.G.; et al. Adverse effects of microplastics and oxidative stress-induced MAPK/Nrf2 pathway-mediated defense mechanisms in the marine copepod Paracyclopina nana. Sci. Rep. 2017, 7, 41323. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Li, Y.; Sepúlveda, M.S.; Jiang, Q.; Jiao, Y.; Chen, Q.; Huang, Y.; Tian, J.; Zhao, Y. Development of an adverse outcome pathway for nanoplastic toxicity in Daphnia pulex using proteomics. Sci. Total Environ. 2021, 766, 144249. [Google Scholar] [CrossRef]
- Hu, M.; Palić, D. Micro- and nano-plastics activation of oxidative and inflammatory adverse outcome pathways. Redox Biol. 2020, 37, 101620. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Chen, B.; Li, Q.; Liu, N.; Xia, B.; Zhu, L.; Qu, K. Toxicities of polystyrene nano- and microplastics toward marine bacterium Halomonas alkaliphila. Sci. Total Environ. 2018, 642, 1378–1385. [Google Scholar] [CrossRef]
- González-Fernández, C.; Tallec, K.; Le Goïc, N.; Lambert, C.; Soudant, P.; Huvet, A.; Suquet, M.; Berchel, M.; Paul-Pont, I. Cellular responses of Pacific oyster (Crassostrea gigas) gametes exposed in vitro to polystyrene nanoparticles. Chemosphere 2018, 208, 764–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, S.T.; Adiseshaiah, P.P.; Crist, R.M. Autophagy and lysosomal dysfunction as emerging mechanisms of nanomaterial toxicity. Part. Fibre Toxicol. 2012, 9, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, H.J.; Semmler-Behnke, M.; Brown, D.M.; Kreyling, W.; Tran, L.; Stone, V. Evaluating the uptake and intracellular fate of polystyrene nanoparticles by primary and hepatocyte cell lines in vitro. Toxicol. Appl. Pharmacol. 2010, 242, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Brandts, I.; Garcia-Ordoñez, M.; Tort, L.; Teles, M.; Roher, N. Polystyrene nanoplastics accumulate in ZFL cell lysosomes and in zebrafish larvae after acute exposure, inducing a synergistic immune response in vitro without affecting larval survival in vivo. Environ. Sci. Nano 2020, 7, 2410–2422. [Google Scholar] [CrossRef]
- Gaspar, T.R.; Chi, R.J.; Parrow, M.W.; Ringwood, A.H. Cellular Bioreactivity of Micro- and Nano-Plastic Particles in Oysters. Front. Mar. Sci. 2018, 5, 345. [Google Scholar] [CrossRef]
- Hemmerich, P.H.; Mikecz, A.H. von Defining the Subcellular Interface of Nanoparticles by Live-Cell Imaging. PLoS ONE 2013, 8, e62018. [Google Scholar] [CrossRef] [Green Version]
- Brun, N.R.; van Hage, P.; Hunting, E.R.; Haramis, A.P.G.; Vink, S.C.; Vijver, M.G.; Schaaf, M.J.M.; Tudorache, C. Polystyrene nanoplastics disrupt glucose metabolism and cortisol levels with a possible link to behavioural changes in larval zebrafish. Commun. Biol. 2019, 2, 382. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, Z.; Yang, Y.; Jiang, Q.; Wu, D.; Huang, Y.; Jiao, Y.; Chen, Q.; Huang, Y.; Zhao, Y. Effects of nanoplastics on energy metabolism in the oriental river prawn (Macrobrachium nipponense). Environ. Pollut. 2021, 268, 115890. [Google Scholar] [CrossRef]
- Seoane, M.; González-Fernández, C.; Soudant, P.; Huvet, A.; Esperanza, M.; Cid, Á.; Paul-Pont, I. Polystyrene microbeads modulate the energy metabolism of the marine diatom Chaetoceros neogracile. Environ. Pollut. 2019, 251, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Tallec, K.; Paul-Pont, I.; Petton, B.; Alunno-Bruscia, M.; Bourdon, C.; Bernardini, I.; Boulais, M.; Lambert, C.; Quéré, C.; Bideau, A.; et al. Amino-nanopolystyrene exposures of oyster (Crassostrea gigas) embryos induced no apparent intergenerational effects. Nanotoxicology 2021, 15, 477–493. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, R.; Uzochukwu, D.; Di Giulio, R.T. PAH Sorption to Nanoplastics and the Trojan Horse Effect as Drivers of Mitochondrial Toxicity and PAH Localization in Zebrafish. Front. Environ. Sci. 2020, 8, 78. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, R.; Voy, C.; Chen, S.; Di Giulio, R.T. Nanoplastics decrease the toxicity of a complex PAH mixture but impair mitochondrial energy production in developing Zebrafish. Environ. Sci. Technol. 2019, 53, 8405–8415. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Gundlach, M.; Yang, S.; Jiang, J.; Velki, M.; Yin, D.; Hollert, H. Quantitative investigation of the mechanisms of microplastics and nanoplastics toward zebrafish larvae locomotor activity. Sci. Total Environ. 2017, 584–585, 1022–1031. [Google Scholar] [CrossRef]
- Silva, M.S.S.; Oliveira, M.; Valente, P.; Figueira, E.; Martins, M.; Pires, A. Behavior and biochemical responses of the polychaeta Hediste diversicolor to polystyrene nanoplastics. Sci. Total Environ. 2020, 707, 134434. [Google Scholar] [CrossRef]
- Varó, I.; Perini, A.; Torreblanca, A.; Garcia, Y.; Bergami, E.; Vannuccini, M.L.; Corsi, I. Time-dependent effects of polystyrene nanoparticles in brine shrimp Artemia franciscana at physiological, biochemical and molecular levels. Sci. Total Environ. 2019, 675, 570–580. [Google Scholar] [CrossRef]
- Brandts, I.; Teles, M.; Gonçalves, A.P.; Barreto, A.; Franco-Martinez, L.; Tvarijonaviciute, A.; Martins, M.A.; Soares, A.M.V.M.; Tort, L.; Oliveira, M. Effects of nanoplastics on Mytilus galloprovincialis after individual and combined exposure with carbamazepine. Sci. Total Environ. 2018, 643, 775–784. [Google Scholar] [CrossRef]
- Ding, J.; Zhang, S.; Razanajatovo, R.M.; Zou, H.; Zhu, W. Accumulation, tissue distribution, and biochemical effects of polystyrene microplastics in the freshwater fish red tilapia (Oreochromis niloticus). Environ. Pollut. 2018, 238, 1–9. [Google Scholar] [CrossRef]
- Brun, N.R.; Koch, B.E.V.; Varela, M.; Peijnenburg, W.J.G.M.; Spaink, H.P.; Vijver, M.G. Nanoparticles induce dermal and intestinal innate immune system responses in zebrafish embryos. Environ. Sci. Nano 2018, 5, 904–916. [Google Scholar] [CrossRef]
- Jin, Y.; Xia, J.; Pan, Z.; Yang, J.; Wang, W.; Fu, Z. Polystyrene microplastics induce microbiota dysbiosis and inflammation in the gut of adult zebrafish. Environ. Pollut. 2018, 235, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Brandts, I.; Balasch, J.C.; Gonçalves, A.P.; Martins, M.A.; Pereira, M.L.; Tvarijonaviciute, A.; Teles, M.; Oliveira, M. Immuno-modulatory effects of nanoplastics and humic acids in the European seabass (Dicentrarchus labrax). J. Hazard. Mater. 2021, 414, 125562. [Google Scholar] [CrossRef] [PubMed]
- Dreier, D.A.; Mello, D.F.; Meyer, J.N.; Martyniuk, C.J. Linking Mitochondrial Dysfunction to Organismal and Population Health in the Context of Environmental Pollutants: Progress and Considerations for Mitochondrial Adverse Outcome Pathways. Environ. Toxicol. Chem. 2019, 38, 1625–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokolova, I.M.; Frederich, M.; Bagwe, R.; Lannig, G.; Sukhotin, A.A. Energy homeostasis as an integrative tool for assessing limits of environmental stress tolerance in aquatic invertebrates. Mar. Environ. Res. 2012, 79, 1–15. [Google Scholar] [CrossRef]
- Wang, L.; Wu, W.M.; Bolan, N.S.; Tsang, D.C.W.; Li, Y.; Qin, M.; Hou, D. Environmental fate, toxicity and risk management strategies of nanoplastics in the environment: Current status and future perspectives. J. Hazard. Mater. 2021, 401, 123415. [Google Scholar] [CrossRef]
- Zhang, Q.; He, Y.; Cheng, R.; Li, Q.; Qian, Z.; Lin, X. Recent advances in toxicological research and potential health impact of microplastics and nanoplastics in vivo. Environ. Sci. Pollut. Res. 2022, 29, 40418–40448. [Google Scholar] [CrossRef]
- Fu, L.; Li, J.; Wang, G.; Luan, Y.; Dai, W. Adsorption behavior of organic pollutants on microplastics. Ecotoxicol. Environ. Saf. 2021, 217, 112207. [Google Scholar] [CrossRef]
- Bandini, F.; Hchaichi, I.; Zitouni, N.; Missawi, O.; Cocconcelli, P.S.; Puglisi, E.; Banni, M. Bacterial community profiling of floating plastics from South Mediterranean sites: First evidence of effects on mussels as possible vehicles of transmission. J. Hazard. Mater. 2021, 411, 125079. [Google Scholar] [CrossRef]
- Li, R.; Zhu, L.; Yang, K.; Li, H.; Zhu, Y.-G.; Cui, L. Impact of Urbanization on Antibiotic Resistome in Different Microplastics: Evidence from a Large-Scale Whole River Analysis. Environ. Sci. Technol. 2021, 55, 8760–8770. [Google Scholar] [CrossRef]
- Taniguchi, S.; Colabuono, F.I.; Dias, P.S.; Oliveira, R.; Fisner, M.; Turra, A.; Izar, G.M.; Abessa, D.M.S.; Saha, M.; Hosoda, J.; et al. Spatial variability in persistent organic pollutants and polycyclic aromatic hydrocarbons found in beach-stranded pellets along the coast of the state of São Paulo, southeastern Brazil. Mar. Pollut. Bull. 2016, 106, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Vedolin, M.C.; Teophilo, C.Y.S.; Turra, A.; Figueira, R.C.L. Spatial variability in the concentrations of metals in beached microplastics. Mar. Pollut. Bull. 2018, 129, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Gorman, D.; Moreira, F.T.; Turra, A.; Fontenelle, F.R.; Combi, T.; Bícego, M.C.; de Castro Martins, C. Organic contamination of beached plastic pellets in the South Atlantic: Risk assessments can benefit by considering spatial gradients. Chemosphere 2019, 223, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Koelmans, A.A.; Besseling, E.; Foekema, E.M. Leaching of plastic additives to marine organisms. Environ. Pollut. 2014, 187, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Koelmans, A.A.; Bakir, A.; Burton, G.A.; Janssen, C.R. Microplastic as a Vector for Chemicals in the Aquatic Environment: Critical Review and Model-Supported Reinterpretation of Empirical Studies. Environ. Sci. Technol. 2016, 50, 3315–3326. [Google Scholar] [CrossRef]
- Huang, B.; Wei, Z.-B.; Yang, L.-Y.; Pan, K.; Miao, A.-J. Combined Toxicity of Silver Nanoparticles with Hematite or Plastic Nanoparticles toward Two Freshwater Algae. Environ. Sci. Technol. 2019, 53, 3871–3879. [Google Scholar] [CrossRef]
- Jeong, C.B.; Kang, H.M.; Byeon, E.; Kim, M.S.; Ha, S.Y.; Kim, M.; Jung, J.H.; Lee, J.S. Phenotypic and transcriptomic responses of the rotifer Brachionus koreanus by single and combined exposures to nano-sized microplastics and water-accommodated fractions of crude oil. J. Hazard. Mater. 2021, 416, 125703. [Google Scholar] [CrossRef]
- Jeong, C.-B.; Kang, H.-M.; Lee, Y.H.; Kim, M.-S.; Lee, J.-S.; Seo, J.S.; Wang, M.; Lee, J.-S. Nanoplastic Ingestion Enhances Toxicity of Persistent Organic Pollutants (POPs) in the Monogonont Rotifer Brachionus koreanus via Multixenobiotic Resistance (MXR) Disruption. Environ. Sci. Technol. 2018, 52, 11411–11418. [Google Scholar] [CrossRef]
- Bhagat, J.; Zang, L.; Nakayama, H.; Nishimura, N.; Shimada, Y. Effects of nanoplastic on toxicity of azole fungicides (ketoconazole and fluconazole) in zebrafish embryos. Sci. Total Environ. 2021, 800, 149463. [Google Scholar] [CrossRef]
- Zhao, H.J.; Xu, J.K.; Yan, Z.H.; Ren, H.Q.; Zhang, Y. Microplastics enhance the developmental toxicity of synthetic phenolic antioxidants by disturbing the thyroid function and metabolism in developing zebrafish. Environ. Int. 2020, 140, 105750. [Google Scholar] [CrossRef]
- Velzeboer, I.; Kwadijk, C.J.A.F.; Koelmans, A.A. Strong Sorption of PCBs to Nanoplastics, Microplastics, Carbon Nanotubes, and Fullerenes. Environ. Sci. Technol. 2014, 48, 4869–4876. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, Z.; Ren, X.; Duan, X. Parental transfer of nanopolystyrene-enhanced tris(1,3-dichloro-2-propyl) phosphate induces transgenerational thyroid disruption in zebrafish. Aquat. Toxicol. 2021, 236, 105871. [Google Scholar] [CrossRef] [PubMed]
- Bellingeri, A.; Bergami, E.; Grassi, G.; Faleri, C.; Redondo-Hasselerharm, P.; Koelmans, A.A.; Corsi, I. Combined effects of nanoplastics and copper on the freshwater alga Raphidocelis subcapitata. Aquat. Toxicol. 2019, 210, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Jiang, R.; Xiong, Y.; Wu, J.; Xu, J.; Zheng, J.; Zhu, F.; Ouyang, G. Quantification of the combined toxic effect of polychlorinated biphenyls and nano-sized polystyrene on Daphnia magna. J. Hazard. Mater. 2019, 364, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.J.; Shi, Y.; Li, X.Y.; Sun, X.D.; Xiao, F.; Sun, J.W.; Wang, Y.; Liu, X.Y.; Wang, S.G.; Yuan, X.Z. Behavior of tetracycline and polystyrene nanoparticles in estuaries and their joint toxicity on marine microalgae Skeletonema costatum. Environ. Pollut. 2020, 263, 114453. [Google Scholar] [CrossRef]
- Zhang, Q.; Qu, Q.; Lu, T.; Ke, M.; Zhu, Y.; Zhang, M.; Zhang, Z.; Du, B.; Pan, X.; Sun, L.; et al. The combined toxicity effect of nanoplastics and glyphosate on Microcystis aeruginosa growth. Environ. Pollut. 2018, 243, 1106–1112. [Google Scholar] [CrossRef]
- Yilimulati, M.; Wang, L.; Ma, X.; Yang, C.; Habibul, N. Adsorption of ciprofloxacin to functionalized nano-sized polystyrene plastic: Kinetics, thermochemistry and toxicity. Sci. Total Environ. 2021, 750, 142370. [Google Scholar] [CrossRef]
- González-Soto, N.; Hatfield, J.; Katsumiti, A.; Duroudier, N.; Lacave, J.M.; Bilbao, E.; Orbea, A.; Navarro, E.; Cajaraville, M.P. Impacts of dietary exposure to different sized polystyrene microplastics alone and with sorbed benzo[a]pyrene on biomarkers and whole organism responses in mussels Mytilus galloprovincialis. Sci. Total Environ. 2019, 684, 548–566. [Google Scholar] [CrossRef]
- Abdolahpur Monikh, F.; Vijver, M.G.; Guo, Z.; Zhang, P.; Darbha, G.K.; Peijnenburg, W.J.G.M. Metal sorption onto nanoscale plastic debris and trojan horse effects in Daphnia magna: Role of dissolved organic matter. Water Res. 2020, 186, 116410. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Chen, M.; He, S.; Fang, C.; Chen, M.; Li, D.; Wu, D.; Chernick, M.; Hinton, D.E.; Bo, J.; et al. Microplastics decrease the toxicity of triphenyl phosphate (TPhP) in the marine medaka (Oryzias melastigma) larvae. Sci. Total Environ. 2021, 763, 143040. [Google Scholar] [CrossRef]
- Davranche, M.; Veclin, C.; Pierson-Wickmann, A.C.; El Hadri, H.; Grassl, B.; Rowenczyk, L.; Dia, A.; Ter Halle, A.; Blancho, F.; Reynaud, S.; et al. Are nanoplastics able to bind significant amount of metals? The lead example. Environ. Pollut. 2019, 249, 940–948. [Google Scholar] [CrossRef] [Green Version]
- Domenech, J.; Cortés, C.; Vela, L.; Marcos, R.; Hernández, A. Polystyrene Nanoplastics as Carriers of Metals. Interactions of Polystyrene Nanoparticles with Silver Nanoparticles and Silver Nitrate, and Their Effects on Human Intestinal Caco-2 Cells. Biomolecules 2021, 11, 859. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Wang, Y.; Shen, D.; Kang, Q.; Ma, J.; Chen, L. Revisiting the cellular toxicity of benzo[a]pyrene from the view of nanoclusters: Size- and nanoplastic adsorption-dependent bioavailability. Nanoscale 2021, 13, 1016–1028. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Yang, B.; Waigi, M.G.; Peng, F.; Li, Z.; Hu, X. The effects of functional groups on the sorption of naphthalene on microplastics. Chemosphere 2020, 261, 127592. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, H.; He, H.; Cheng, X.; Ma, T.; Hu, J.; Yang, S.; Li, S.; Zhang, L. Adsorption behavior and mechanism of 9-Nitroanthracene on typical microplastics in aqueous solutions. Chemosphere 2020, 245, 125628. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Meng, H.; Kong, X.; Cheng, X.; Ma, T.; He, H.; Du, W.; Yang, S.; Li, S.; Zhang, L. Combined effects of polyethylene and organic contaminant on zebrafish (Danio rerio): Accumulation of 9-Nitroanthracene, biomarkers and intestinal microbiota. Environ. Pollut. 2021, 277, 116767. [Google Scholar] [CrossRef]
- Gu, H.; Wei, S.; Hu, M.; Wei, H.; Wang, X.; Shang, Y.; Li, L.; Shi, H.; Wang, Y. Microplastics aggravate the adverse effects of BDE-47 on physiological and defense performance in mussels. J. Hazard. Mater. 2020, 398, 122909. [Google Scholar] [CrossRef]
- Scopetani, C.; Cincinelli, A.; Martellini, T.; Lombardini, E.; Ciofini, A.; Fortunati, A.; Pasquali, V.; Ciattini, S.; Ugolini, A. Ingested microplastic as a two-way transporter for PBDEs in Talitrus saltator. Environ. Res. 2018, 167, 411–417. [Google Scholar] [CrossRef]
- Devriese, L.I.; De Witte, B.; Vethaak, A.D.; Hostens, K.; Leslie, H.A. Bioaccumulation of PCBs from microplastics in Norway lobster (Nephrops norvegicus): An experimental study. Chemosphere 2017, 186, 10–16. [Google Scholar] [CrossRef]
- Gerdes, Z.; Ogonowski, M.; Nybom, I.; Ek, C.; Adolfsson-Erici, M.; Barth, A.; Gorokhova, E. Microplastic-mediated transport of PCBs? A depuration study with Daphnia magna. PLoS ONE 2019, 14, e0205378. [Google Scholar] [CrossRef] [Green Version]
- Rummel, C.D.; Adolfsson-Erici, M.; Jahnke, A.; MacLeod, M. No measurable “cleaning” of polychlorinated biphenyls from Rainbow Trout in a 9 week depuration study with dietary exposure to 40% polyethylene microspheres. Environ. Sci. Process. Impacts 2016, 18, 788–795. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.; Mondon, J.; Merenda, A.; Dumée, L.F.; Callahan, D.L. Surface adsorption of metallic species onto microplastics with long-term exposure to the natural marine environment. Sci. Total Environ. 2021, 780, 146613. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, W.; Yang, X.; Wang, J.; Lin, H.; Yang, Y. Microplastics are a hotspot for antibiotic resistance genes: Progress and perspective. Sci. Total Environ. 2021, 773, 145643. [Google Scholar] [CrossRef] [PubMed]
- Rummel, C.D.; Lechtenfeld, O.J.; Kallies, R.; Benke, A.; Herzsprung, P.; Rynek, R.; Wagner, S.; Potthoff, A.; Jahnke, A.; Schmitt-Jansen, M. Conditioning Film and Early Biofilm Succession on Plastic Surfaces. Environ. Sci. Technol. 2021, 55, 11006–11018. [Google Scholar] [CrossRef] [PubMed]
- Jacquin, J.; Cheng, J.; Odobel, C.; Pandin, C.; Conan, P.; Pujo-Pay, M.; Barbe, V.; Meistertzheim, A.-L.; Ghiglione, J.-F. Microbial Ecotoxicology of Marine Plastic Debris: A Review on Colonization and Biodegradation by the “Plastisphere. ” Front. Microbiol. 2019, 10, 865. [Google Scholar] [CrossRef] [Green Version]
- Arias-Andres, M.; Kettner, M.T.; Miki, T.; Grossart, H.P. Microplastics: New substrates for heterotrophic activity contribute to altering organic matter cycles in aquatic ecosystems. Sci. Total Environ. 2018, 635, 1152–1159. [Google Scholar] [CrossRef] [PubMed]
- Fabra, M.; Williams, L.; Watts, J.E.M.; Hale, M.S.; Couceiro, F.; Preston, J. The plastic Trojan horse: Biofilms increase microplastic uptake in marine filter feeders impacting microbial transfer and organism health. Sci. Total Environ. 2021, 797, 149217. [Google Scholar] [CrossRef] [PubMed]
- Murano, C.; Donnarumma, V.; Corsi, I.; Casotti, R.; Palumbo, A. Impact of Microbial Colonization of Polystyrene Microbeads on the Toxicological Responses in the Sea Urchin Paracentrotus lividus. Environ. Sci. Technol. 2021, 55, 7990–8000. [Google Scholar] [CrossRef]
- Hodgson, D.J.; Bréchon, A.L.; Thompson, R.C. Ingestion and fragmentation of plastic carrier bags by the amphipod Orchestia gammarellus: Effects of plastic type and fouling load. Mar. Pollut. Bull. 2018, 127, 154–159. [Google Scholar] [CrossRef]
- Jin, M.; Yu, X.; Yao, Z.; Tao, P.; Li, G.; Yu, X.; Zhao, J.L.; Peng, J. How biofilms affect the uptake and fate of hydrophobic organic compounds (HOCs) in microplastic: Insights from an In situ study of Xiangshan Bay, China. Water Res. 2020, 184, 116118. [Google Scholar] [CrossRef]
- de Oliveira, T.T.S.; Andreu, I.; Machado, M.C.; Vimbela, G.; Tripathi, A.; Bose, A. Interaction of Cyanobacteria with Nanometer and Micron Sized Polystyrene Particles in Marine and Fresh Water. Langmuir 2020, 36, 3963–3969. [Google Scholar] [CrossRef]
- Yan, M.; Li, W.; Chen, X.; He, Y.; Zhang, X.; Gong, H. A preliminary study of the association between colonization of microorganism on microplastics and intestinal microbiota in shrimp under natural conditions. J. Hazard. Mater. 2021, 408, 124882. [Google Scholar] [CrossRef] [PubMed]
- Avio, C.G.; Gorbi, S.; Milan, M.; Benedetti, M.; Fattorini, D.; D’Errico, G.; Pauletto, M.; Bargelloni, L.; Regoli, F. Pollutants bioavailability and toxicological risk from microplastics to marine mussels. Environ. Pollut. 2015, 198, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Guo, Y.; Hu, C.; Lam, P.K.S.; Lam, J.C.W.; Zhou, B. Dysbiosis of gut microbiota by chronic coexposure to titanium dioxide nanoparticles and bisphenol A: Implications for host health in zebrafish. Environ. Pollut. 2018, 234, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Adamovsky, O.; Buerger, A.N.; Vespalcova, H.; Sohag, S.R.; Hanlon, A.T.; Ginn, P.E.; Craft, S.L.; Smatana, S.; Budinska, E.; Persico, M.; et al. Evaluation of Microbiome-Host Relationships in the Zebrafish Gastrointestinal System Reveals Adaptive Immunity Is a Target of Bis(2-ethylhexyl) Phthalate (DEHP) Exposure. Environ. Sci. Technol. 2020, 54, 5719–5728. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Gan, R.; Liu, J.; Xie, Y.; Xu, D.; Xiang, Y.; Su, J.; Teng, Z.; Hou, J. Adsorption and desorption behaviors of antibiotics by tire wear particles and polyethylene microplastics with or without aging processes. Sci. Total Environ. 2021, 771, 145451. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhu, Z.; Yang, Y.; Sun, Y.; Yu, F.; Ma, J. Sorption behavior and mechanism of hydrophilic organic chemicals to virgin and aged microplastics in freshwater and seawater. Environ. Pollut. 2019, 246, 26–33. [Google Scholar] [CrossRef]
- Huang, X.; Zemlyanov, D.Y.; Diaz-Amaya, S.; Salehi, M.; Stanciu, L.; Whelton, A.J. Competitive heavy metal adsorption onto new and aged polyethylene under various drinking water conditions. J. Hazard. Mater. 2020, 385, 121585. [Google Scholar] [CrossRef]
- Hüffer, T.; Weniger, A.K.; Hofmann, T. Sorption of organic compounds by aged polystyrene microplastic particles. Environ. Pollut. 2018, 236, 218–225. [Google Scholar] [CrossRef]
- Oliveira, M.M.; Proenca, A.M.; Moreira-Silva, E.; de Castro, A.M.; dos Santos, F.M.; Marconatto, L.; Medina-Silva, R. Biofilms of Pseudomonas and Lysinibacillus Marine Strains on High-Density Polyethylene. Microb. Ecol. 2021, 81, 833–846. [Google Scholar] [CrossRef]
- Li, X.; Ji, S.; He, E.; Peijnenburg, W.J.G.M.; Cao, X.; Zhao, L.; Xu, X.; Zhang, P.; Qiu, H. UV/ozone induced physicochemical transformations of polystyrene nanoparticles and their aggregation tendency and kinetics with natural organic matter in aqueous systems. J. Hazard. Mater. 2022, 433, 128790. [Google Scholar] [CrossRef]
- Reichel, J.; Grabmann, J.; Knoop, O.; Drewes, J.E.; Letzel, T. Organic Contaminants and Interactions with Micro- and Nano-Plastics in the Aqueous Environment: Review of Analytical Methods. Molecules 2021, 26, 1164. [Google Scholar] [CrossRef] [PubMed]
- Sadler, D.E.; Brunner, F.S.; Plaistow, S.J. Temperature and clone-dependent effects of microplastics on immunity and life history in Daphnia magna. Environ. Pollut. 2019, 255, 113178. [Google Scholar] [CrossRef] [PubMed]
- Jaikumar, G.; Baas, J.; Brun, N.R.; Vijver, M.G.; Bosker, T. Acute sensitivity of three Cladoceran species to different types of microplastics in combination with thermal stress. Environ. Pollut. 2018, 239, 733–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilhermino, L.; Martins, A.; Cunha, S.; Fernandes, J.O. Long-term adverse effects of microplastics on Daphnia magna reproduction and population growth rate at increased water temperature and light intensity: Combined effects of stressors and interactions. Sci. Total Environ. 2021, 784, 147082. [Google Scholar] [CrossRef] [PubMed]
- Serra, T.; Barcelona, A.; Pous, N.; Salvadó, V.; Colomer, J. Synergistic effects of water temperature, microplastics and ammonium as second and third order stressors on Daphnia magna. Environ. Pollut. 2020, 267, 115439. [Google Scholar] [CrossRef] [PubMed]
- Fonte, E.; Ferreira, P.; Guilhermino, L. Temperature rise and microplastics interact with the toxicity of the antibiotic cefalexin to juveniles of the common goby (Pomatoschistus microps): Post-exposure predatory behaviour, acetylcholinesterase activity and lipid peroxidation. Aquat. Toxicol. 2016, 180, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Zhang, N.; Jin, S.R.; Chen, Z.Z.; Gao, J.Z.; Liu, Y.; Liu, H.P.; Xu, Z. Microplastics have a more profound impact than elevated temperatures on the predatory performance, digestion and energy metabolism of an Amazonian cichlid. Aquat. Toxicol. 2018, 195, 67–76. [Google Scholar] [CrossRef]
- Wu, J.; Jiang, R.; Lin, W.; Ouyang, G. Effect of salinity and humic acid on the aggregation and toxicity of polystyrene nanoplastics with different functional groups and charges. Environ. Pollut. 2019, 245, 836–843. [Google Scholar] [CrossRef]
- Shupe, H.J.; Boenisch, K.M.; Harper, B.J.; Brander, S.M.; Harper, S.L. Effect of Nanoplastic Type and Surface Chemistry on Particle Agglomeration over a Salinity Gradient. Environ. Toxicol. Chem. 2021, 40, 1820–1826. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhao, J.; Li, L.; Wang, Y.; Dai, X.; Yu, F.; Ma, J. Interfacial interaction between micro/nanoplastics and typical PPCPs and nanoplastics removal via electrosorption from an aqueous solution. Water Res. 2020, 184, 116100. [Google Scholar] [CrossRef]
- Llorca, M.; Schirinzi, G.; Martínez, M.; Barceló, D.; Farré, M. Adsorption of perfluoroalkyl substances on microplastics under environmental conditions. Environ. Pollut. 2018, 235, 680–691. [Google Scholar] [CrossRef] [PubMed]
- Fadare, O.O.; Wan, B.; Liu, K.; Yang, Y.; Zhao, L.; Guo, L.-H. Eco-Corona vs. Protein Corona: Effects of Humic Substances on Corona Formation and Nanoplastic Particle Toxicity in Daphnia magna. Environ. Sci. Technol. 2020, 54, 8001–8009. [Google Scholar] [CrossRef] [PubMed]
- Aljaibachi, R.; Laird, W.B.; Stevens, F.; Callaghan, A. Impacts of polystyrene microplastics on Daphnia magna: A laboratory and a mesocosm study. Sci. Total Environ. 2020, 705, 135800. [Google Scholar] [CrossRef] [PubMed]
- Aljaibachi, R.; Callaghan, A. Impact of polystyrene microplastics on Daphnia magna mortality and reproduction in relation to food availability. PeerJ 2018, 2018, e4601. [Google Scholar] [CrossRef] [Green Version]
- Ogonowski, M.; Schür, C.; Jarsén, Å.; Gorokhova, E. The effects of natural and anthropogenic microparticles on individual fitness in daphnia magna. PLoS ONE 2016, 11, 155063. [Google Scholar] [CrossRef]
- Rodríguez-Torres, R.; Almeda, R.; Kristiansen, M.; Rist, S.; Winding, M.S.; Nielsen, T.G. Ingestion and impact of microplastics on arctic Calanus copepods. Aquat. Toxicol. 2020, 228, 105631. [Google Scholar] [CrossRef]
- Liu, P.; Zhan, X.; Wu, X.; Li, J.; Wang, H.; Gao, S. Effect of weathering on environmental behavior of microplastics: Properties, sorption and potential risks. Chemosphere 2020, 242, 125193. [Google Scholar] [CrossRef]
- Holmes, L.A.; Turner, A.; Thompson, R.C. Interactions between trace metals and plastic production pellets under estuarine conditions. Mar. Chem. 2014, 167, 25–32. [Google Scholar] [CrossRef]
- Alimi, O.S.; Budarz, J.F.; Hernandez, L.M.; Tufenkji, N. Microplastics and Nanoplastics in Aquatic Environments: Aggregation, Deposition, and Enhanced Contaminant Transport. Environ. Sci. Technol. 2018, 52, 1704–1724. [Google Scholar] [CrossRef]
- Guo, J.J.; Huang, X.P.; Xiang, L.; Wang, Y.Z.; Li, Y.W.; Li, H.; Cai, Q.Y.; Mo, C.H.; Wong, M.H. Source, migration and toxicology of microplastics in soil. Environ. Int. 2020, 137, 105263. [Google Scholar] [CrossRef]
- Wang, F.; Shih, K.M.; Li, X.Y. The partition behavior of perfluorooctanesulfonate (PFOS) and perfluorooctanesulfonamide (FOSA) on microplastics. Chemosphere 2015, 119, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Kleber, M.; Sollins, P.; Sutton, R. A conceptual model of organo-mineral interactions in soils: Self-assembly of organic molecular fragments into zonal structures on mineral surfaces. Biogeochemistry 2007, 85, 9–24. [Google Scholar] [CrossRef]
- Lambert, S.; Sinclair, C.J.; Bradley, E.L.; Boxall, A.B.A. Effects of environmental conditions on latex degradation in aquatic systems. Sci. Total Environ. 2013, 447, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Besseling, E.; Quik, J.T.K.; Sun, M.; Koelmans, A.A. Fate of nano- and microplastic in freshwater systems: A modeling study. Environ. Pollut. 2017, 220, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Galloway, T.S.; Cole, M.; Lewis, C. Interactions of microplastic debris throughout the marine ecosystem. Nat. Ecol. Evol. 2017, 1, 116. [Google Scholar] [CrossRef]
- Shiu, R.F.; Vazquez, C.I.; Tsai, Y.Y.; Torres, G.V.; Chen, C.S.; Santschi, P.H.; Quigg, A.; Chin, W.C. Nano-plastics induce aquatic particulate organic matter (microgels) formation. Sci. Total Environ. 2020, 706, 135681. [Google Scholar] [CrossRef]
- Stankus, D.P.; Lohse, S.E.; Hutchison, J.E.; Nason, J.A. Interactions between Natural Organic Matter and Gold Nanoparticles Stabilized with Different Organic Capping Agents. Environ. Sci. Technol. 2010, 45, 3238–3244. [Google Scholar] [CrossRef]
- Baalousha, M.; Manciulea, A.; Cumberland, S.; Kendall, K.; Lead, J.R. Aggregation and surface properties of iron oxide nanoparticles: Influence of ph and natural organic matter. Environ. Toxicol. Chem. 2008, 27, 1875–1882. [Google Scholar] [CrossRef]
- Harrison, J.P.; Sapp, M.; Schratzberger, M.; Mark Osborn, A. Interactions Between Microorganisms and Marine Microplastics: A Call for Research. Mar. Technol. Soc. J. 2011, 45, 12–20. [Google Scholar] [CrossRef]
- Cole, M.; Lindeque, P.K.; Fileman, E.; Clark, J.; Lewis, C.; Halsband, C.; Galloway, T.S. Microplastics Alter the Properties and Sinking Rates of Zooplankton Faecal Pellets. Environ. Sci. Technol. 2016, 50, 3239–3246. [Google Scholar] [CrossRef] [Green Version]
- Wiedner, C.; Vareschi, E. Evaluation of a fluorescent microparticle technique for measuring filtering rates of Daphnia. Hydrobiologia 1995, 302.2, 89–96. [Google Scholar] [CrossRef]
- Piccardo, M.; Renzi, M.; Terlizzi, A. Nanoplastics in the oceans: Theory, experimental evidence and real world. Mar. Pollut. Bull. 2020, 157, 111317. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.C. Microplastics in the Marine Environment: Sources, Consequences and Solutions. In Marine Anthropogenic Litter; Bergmann, M., Gutow, L., Klages, M., Eds.; Springer: Cham, Switzerland, 2015; pp. 85–200. [Google Scholar] [CrossRef] [Green Version]
- Ding, R.; Tong, L.; Zhang, W. Microplastics in Freshwater Environments: Sources, Fates and Toxicity. Water. Air. Soil Pollut. 2021, 232, 181. [Google Scholar] [CrossRef]
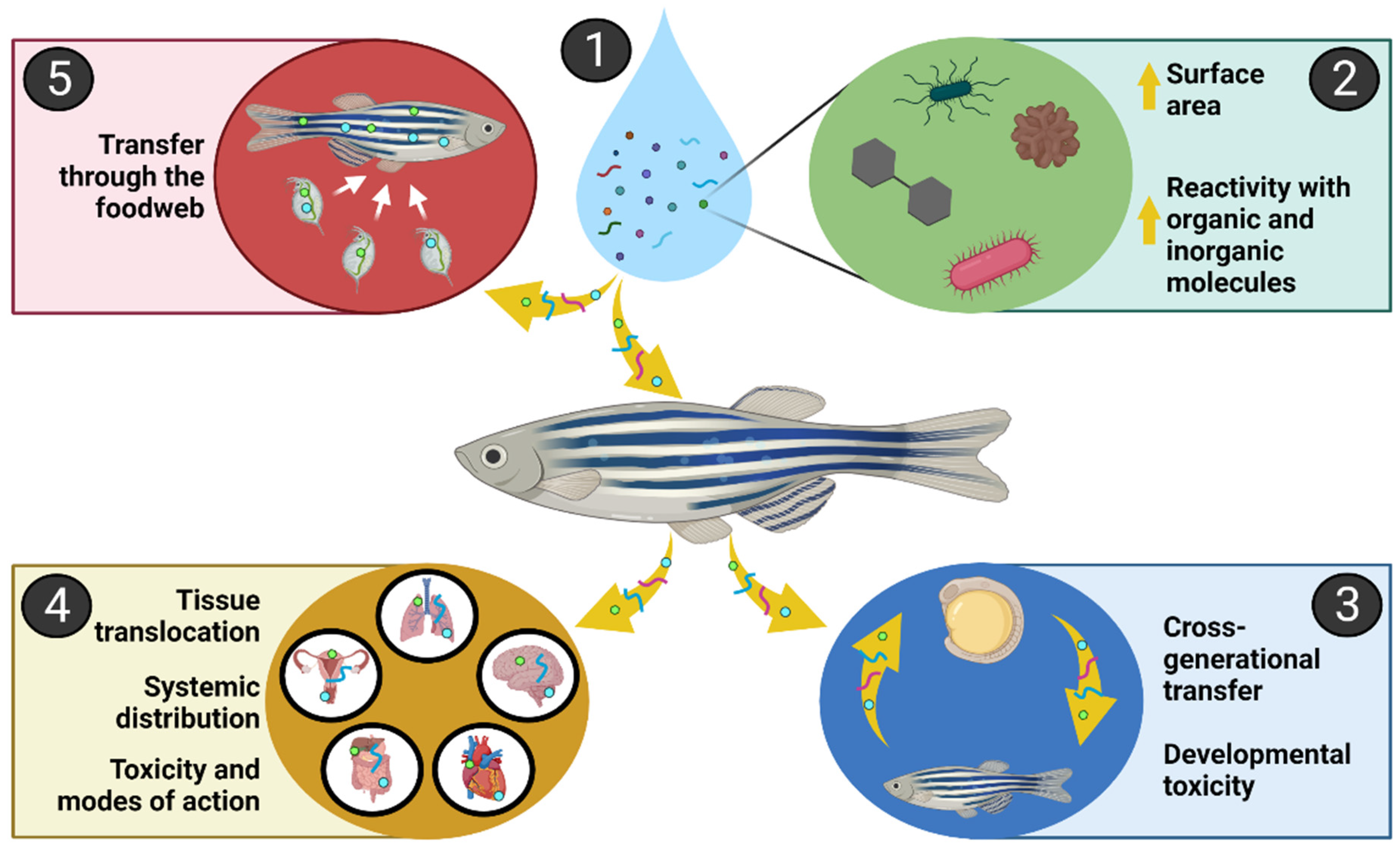
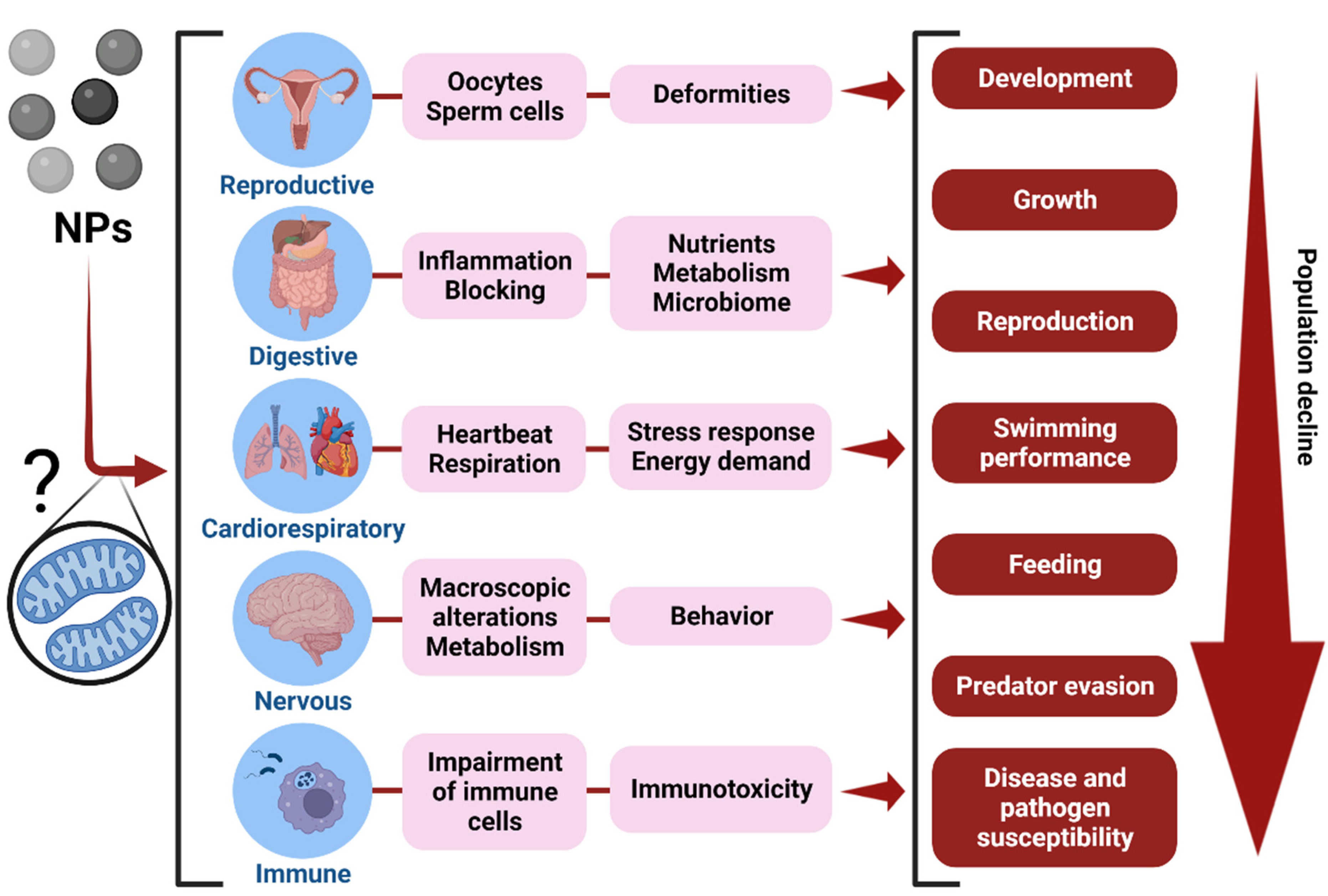
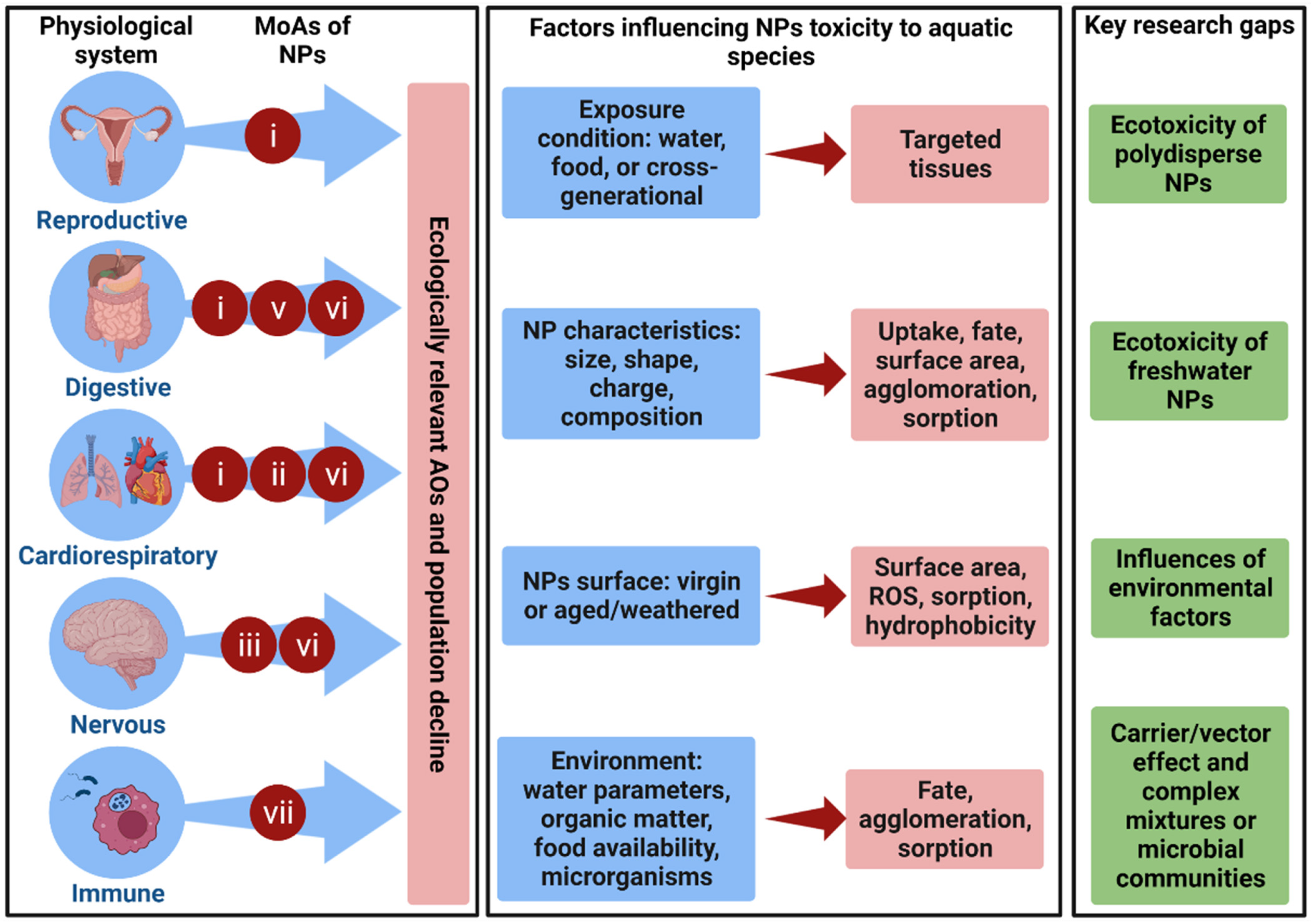

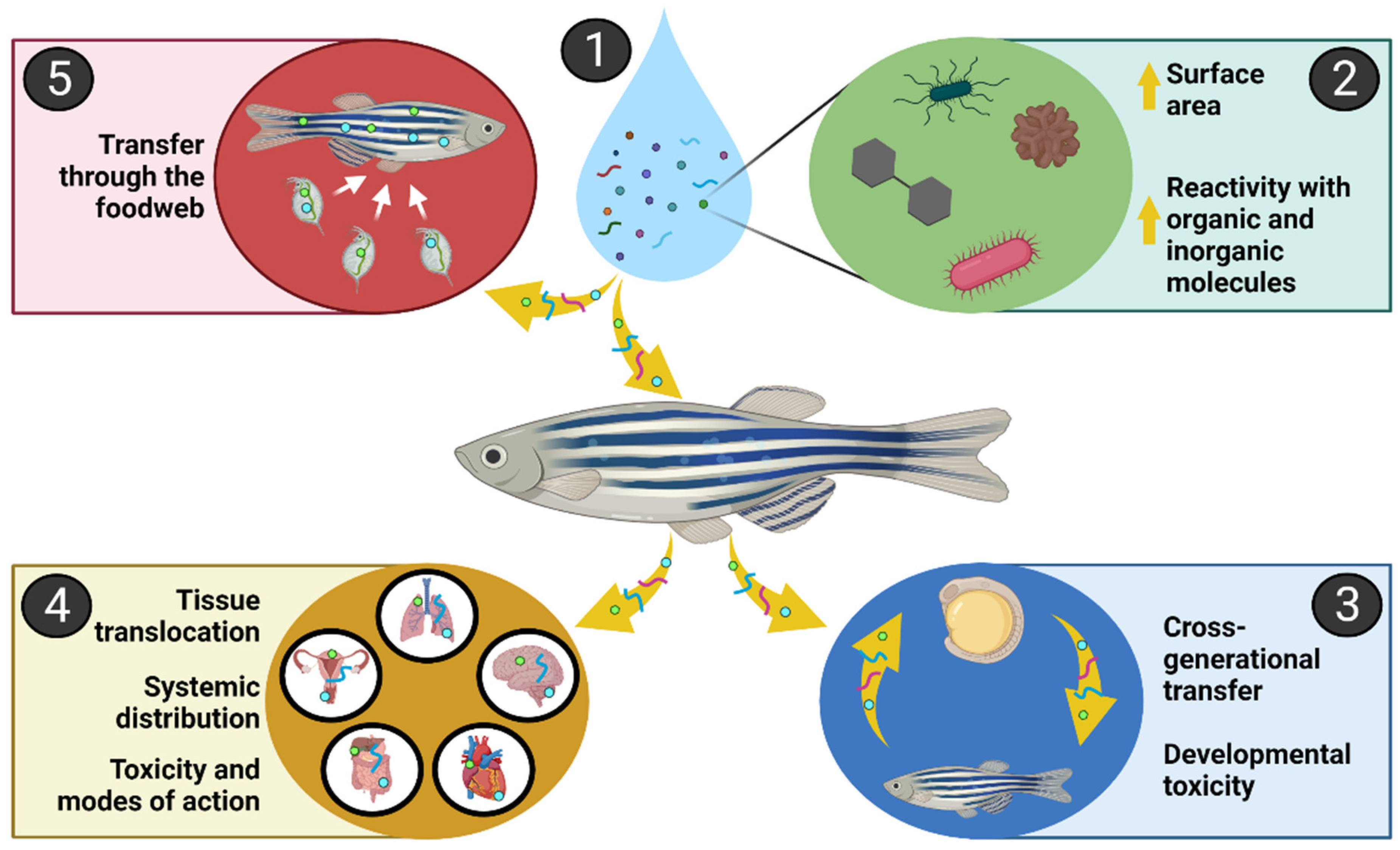{kind=link}
{kind=link}
{kind=link}
| NP Type * and Size # | Additional Environmental Pollutant | Effects Detected | References |
|---|---|---|---|
| PS-NP (10 nm) | Ag+ from silver nanoparticles | Ag+ reacts with neutral or charged PS-NPs. Depending on the species, sorption of Ag+ promotes or lowers bioaccumulation and toxicity to microalgae. | [79] |
| PS-NP (44 nm) | Complex PAH mixture (sediment extract) | Co-exposure decreases PAH bioaccumulation and developmental toxicity in developing fish through sorption to NPs and lower water PHAs levels. NPs impair mitochondrial energy production. | [58] |
| PS-NP (44 nm) | Complex PAH mixture (sediment extract) | PAH-spiked NPs transfer PAHs to the brain and the yolk sac, and cause greater mitochondrial toxicity in developing fish. | [57] |
| PS-NP (50 nm) | Complex PAH mixture (crude oil WAF) | Co-exposures increase PS toxicity, impairing rotifer reproduction, mitochondrial function, energy metabolism, and population growth. | [80] |
| PS-NP (50 nm) | Triclosan | NPs block efflux proteins, increasing triclosan’s toxicity and bioaccumulation. | [81] |
| PS-NP (50 nm) | Ketoconazole or fluconazole | Co-exposure enhances the developmental and cardiovascular toxicity to developing fish. | [82] |
| PS-NP (65 nm) | Butylated hydroxyanisole | Co-exposure increases bioaccumulation of this synthetic antioxidant in developing fish and impairs fish growth and energy metabolism. | [83] |
| PS-NP (70 nm) | Various PCB congeners | PCB interactions with PS aromatic structure occur via hydrophobic and π–π interactions. | [84] |
| PS-NP (70 nm) | TDCIPP | TDCIPP binds to NPs and accumulates in fish gonads and digestive organs. NPs and TDCIPP are transferred to offspring and cause endocrine dysregulation. | [85] |
| COOH-PS-NP (88 nm) | Cu2+ | Cu2+ does not bind to COOH-PS-NPs or alter toxicity to microalgae. | [86] |
| PS-NP (100 nm) | PCB-18 | Sorption of PCB-18 increases NPs toxicity to daphnids. Low doses of NPs reduce the toxicity of PCB-18 by decreasing free PCB-18 levels in the water. | [87] |
| PS-NP, HSO3-PS-NP, and NH2-PS-NP (100 nm) | Tetracycline | Tetracycline binds to plain or HSO3-PS-NPs through weak interactions or to NH2-PS-NPs by partition function. Tetracycline (neutral or negatively charged) has restricted sorption to negatively charged NPs. Higher salinity increases NPs zeta potential and sorption of tetracycline. NPs containing tetracycline were more toxic to microalgae. | [88] |
| NH2-PS-NP (23 nm) | Glyphosate | Glyphosate binds to NH2-NPs by the interaction of oxidizing and reducing functional groups. NH2-NPs favor the stability and uptake of glyphosate by microalgae. | [89] |
| COOH-PS-NP (200 and 500 nm) | Ciprofloxacin | Smaller NPs have a greater adsorption affinity towards ciprofloxacin. Sorption to NPs increases the toxicity to nematodes. | [90] |
| PS-NP (500 nm) | BaP | Exposure to BaP-spiked PS-NPs promotes BaP bioaccumulation in mussels, affecting the immune, digestive, and antioxidant systems. | [91] |
| PS-NP and PE-NP (300 and 600 nm) | Ag+ | Greater surface areas for sorption of Ag+ in 300 nm NPs. Higher PE-NPs agglomeration reduced surface area and Ag+ sorption compared to PS-NPs. NOM decreases the sorption of Ag+ to NPs. Sorption of Ag+ increases NPs toxicity to daphnids. | [92] |
| PS-NP (1000 nm) | Triphenyl phosphate | NPs can reduce the toxicity of triphenyl phosphate to fish embryos by decreasing free triphenyl phosphate levels in water | [93] |
| Various (140–450 nm) | Pb2+ | Higher pH values favor Pb2+ sorption to NPs through electrostatic interaction or covalent binding via NPs functional group deprotonation. | [94] |
| Environmental Factors | Plastic Type * and Size # | Main Effect | References |
|---|---|---|---|
| Temperature | COOH-PS-NPs (500 nm) | Temperature increases the impact of genotypic immunological responses of daphnids to plastics. | [126] |
| Temperature | MPs (1–5 µm, polymer not described) and secondary PE-MPs (1–10 µm) | Rising temperature increases acute sensitivity to MPs in daphnids, but not in ceriodaphnids. | [127] |
| Temperature | PE-MPs (1–5 μm) | Daphnid MP-driven population fitness is unaffected by high temperatures. | [128] |
| Temperature | PS-MPs (5–100 µm) | Co-exposure to elevated temperatures and ammonia increases the impacts of MPs on daphnid survival and feeding rate. | [129] |
| Temperature | PE-MPs (1–5 µm) | Higher water temperatures increase MP-exposed fish mortality and decrease predatory performance. | [130] |
| Temperature | PE-MPs (70–88 μm) | Temperature rise enhances MP bioaccumulation but does not affect fish survival or predatory performance. | [131] |
| Salinity | PS-NPs COOH-PS-NPs, and NH2-PS-NPs (100 nm) | High salinity causes NP aggregation and sedimentation. | [132] |
| Salinity | PMMA-NPs, COOH-PPMA-NPs, and blue and red PS-NPs (55 to 62 nm) | Effects of salinity on NP aggregation depend on NP composition and surface chemistry. | [133] |
| Salinity | PS-NPs (40 nm) | The sorption capacity of NPs to ciprofloxacin and bisphenol A increases as salinity increases. However, it is reduced at extreme salinities. | [134] |
| Salinity | PE-MPs (10−180 μm) and PS-NPs (70 nm) | Higher salinities increase the sorption of PCBs to PE-MPs and PS-NPs. | [84] |
| pH | HDPE-MPs (3–16 µm), PS-MPs (10 µm), and PS-COOH-MPs (10 µm) | Higher pH values enhance the sorption of perfluoroalkyl compounds to MPs. | [135] |
| Natural organic matter | PS-NPs (110 nm) | Natural organic matter, humic acid, and fulvic acid reduce the acute toxicity of PS-NPs to daphnids. Natural organic matter and humic acid mitigate the expression of genes related to detoxification, oxidative stress, and endocrine activity. | [136] |
| Natural organic matter | PS-NPs (50–300 nm) | Decreases aggregation of NPs and toxicity to daphnids. | [132] |
| Food availability | PS-MPs (2 μm) | Daphnids select natural food over MPs with abundant food supply. | [137] |
| Food availability | PS-MPs (15 μm) | Reduces ingestion of MPs by daphnids at higher algal levels. | [138] |
| Food availability | PE-MPs (2.6 µm), MP (1–5 μm, polymer not described) | Food availability has no impact on MP ingestion in daphnids | [139] |
| Food availability | PE-MP (20 µm) | Food availability does not affect food ingestion in copepods | [140] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trevisan, R.; Ranasinghe, P.; Jayasundara, N.; Di Giulio, R.T. Nanoplastics in Aquatic Environments: Impacts on Aquatic Species and Interactions with Environmental Factors and Pollutants. Toxics 2022, 10, 326. https://doi.org/10.3390/toxics10060326
Trevisan R, Ranasinghe P, Jayasundara N, Di Giulio RT. Nanoplastics in Aquatic Environments: Impacts on Aquatic Species and Interactions with Environmental Factors and Pollutants. Toxics. 2022; 10(6):326. https://doi.org/10.3390/toxics10060326
Chicago/Turabian StyleTrevisan, Rafael, Prabha Ranasinghe, Nishad Jayasundara, and Richard T. Di Giulio. 2022. "Nanoplastics in Aquatic Environments: Impacts on Aquatic Species and Interactions with Environmental Factors and Pollutants" Toxics 10, no. 6: 326. https://doi.org/10.3390/toxics10060326
APA StyleTrevisan, R., Ranasinghe, P., Jayasundara, N., & Di Giulio, R. T. (2022). Nanoplastics in Aquatic Environments: Impacts on Aquatic Species and Interactions with Environmental Factors and Pollutants. Toxics, 10(6), 326. https://doi.org/10.3390/toxics10060326