

Zinc Biofortification through Basal Zinc Supply Reduces Grain Cadmium in Mung Beans: Metal Partitioning and Health Risks Assessment




Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Collection, Processing, Analysis, and Experimental Design
2.2. Plant Growth
2.3. Plant Harvest and Analysis
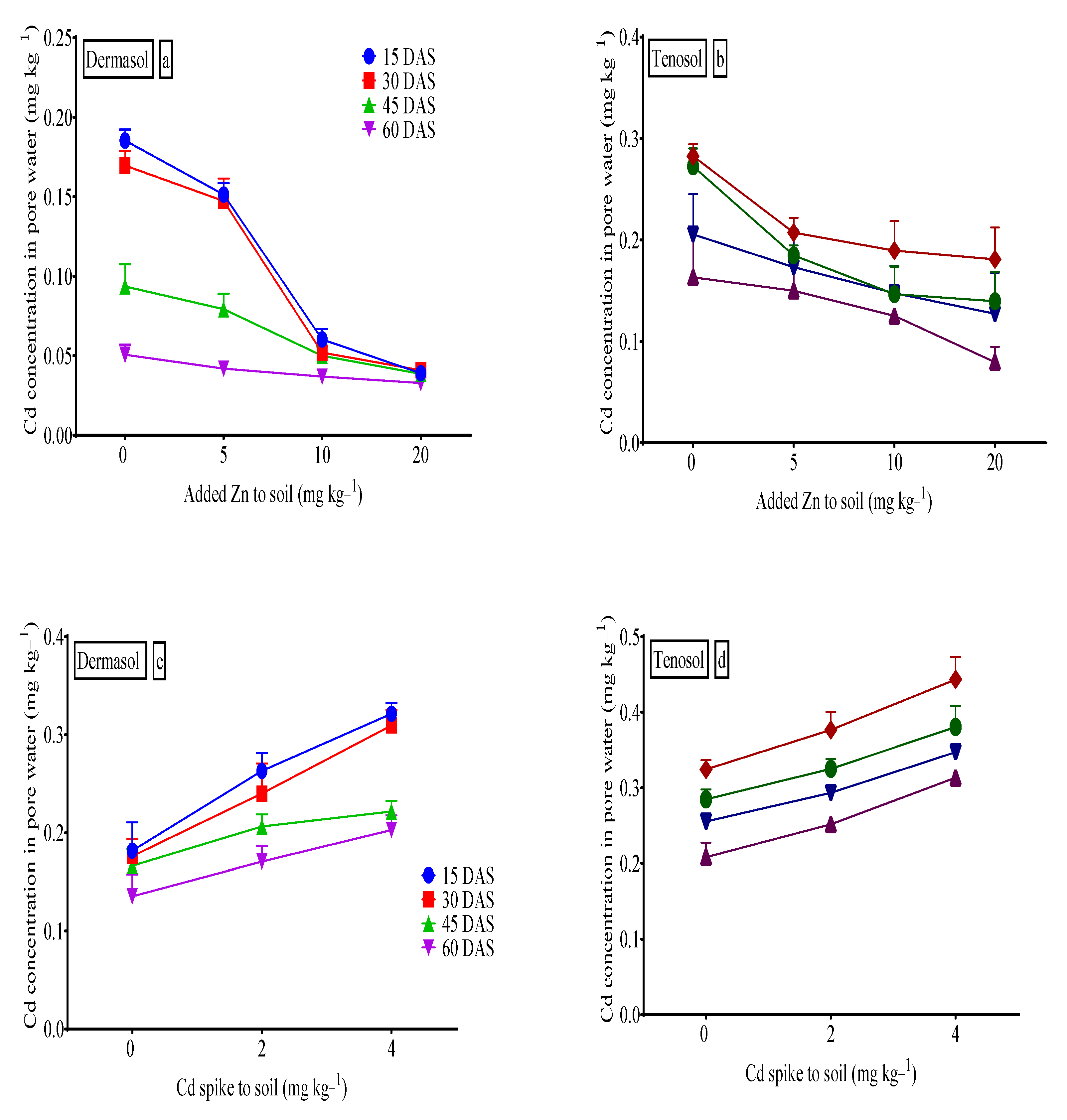
2.4. Soil Pore Water Analysis
2.5. Human Health Risks Assessment
2.5.1. Measurement of Daily Intake of Heavy Metals
2.5.2. Non-Carcinogenic Health Risk
2.5.3. Carcinogenic Risk
2.6. Statistical Analysis
3. Results
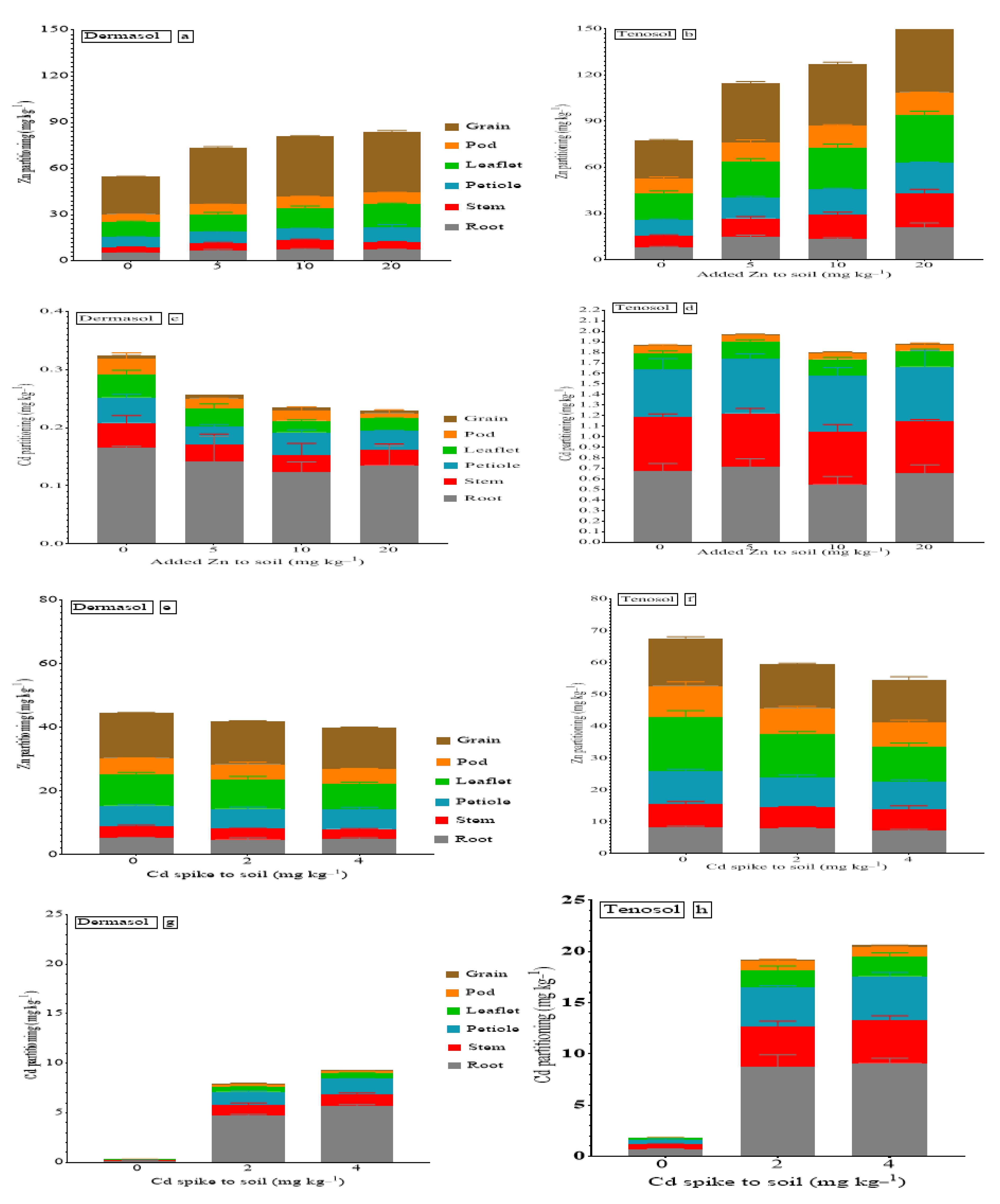
3.1. Partitioning of Cadmium and Zinc in Mung Beans
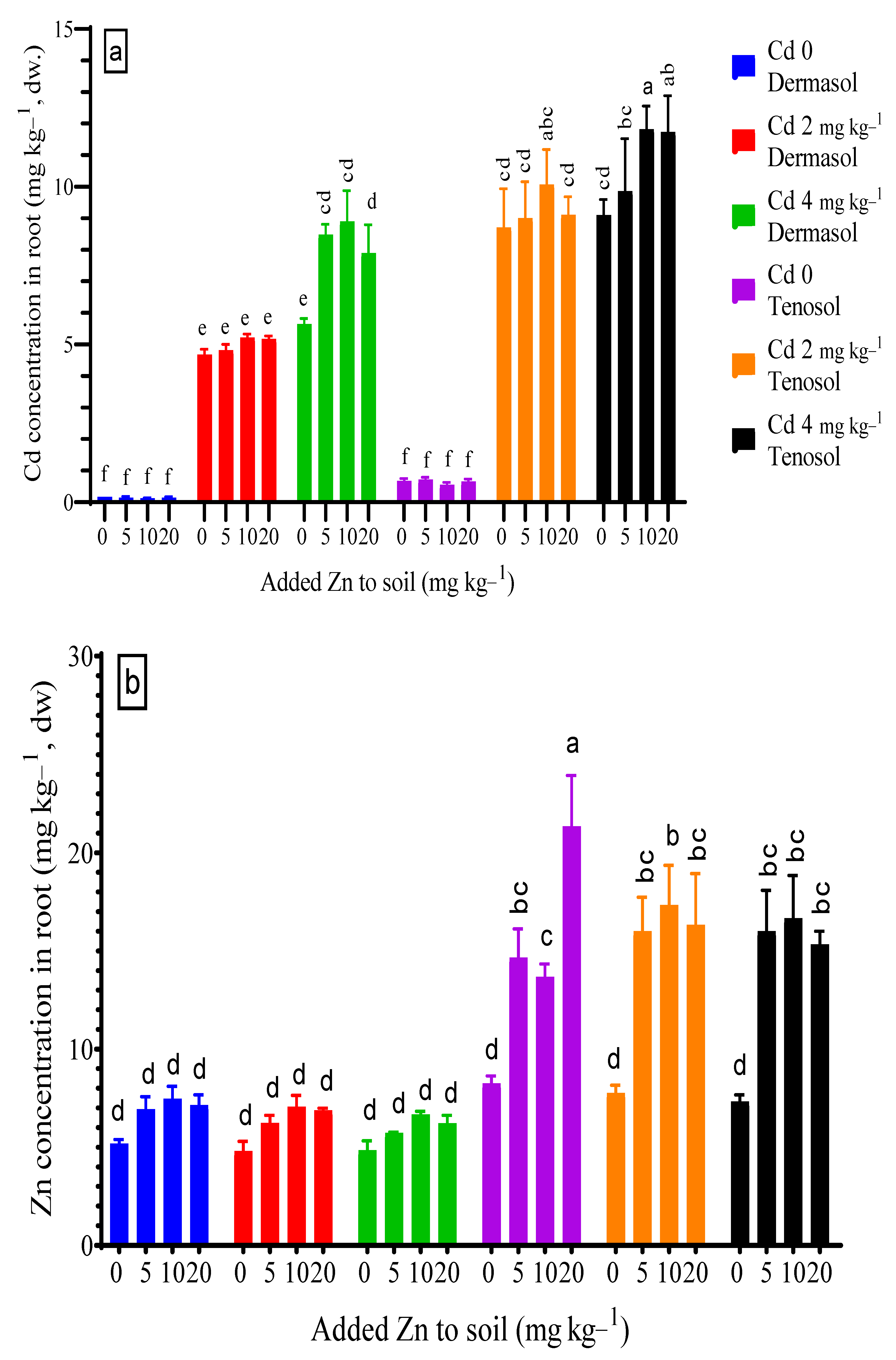
3.2. Concentration of Cadmium and Zinc in Roots
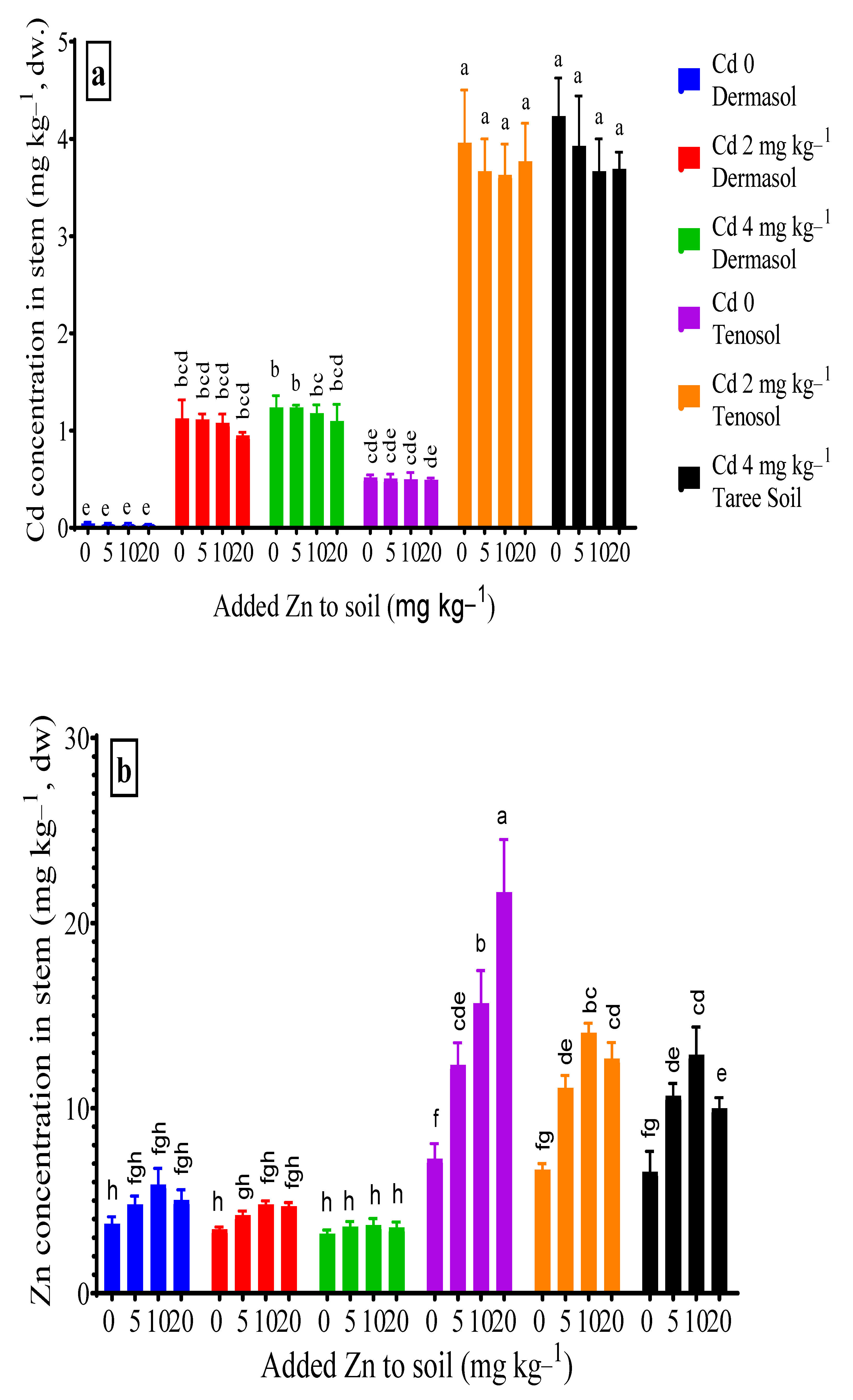
3.3. Concentrations of Cadmium and Zinc in Stem
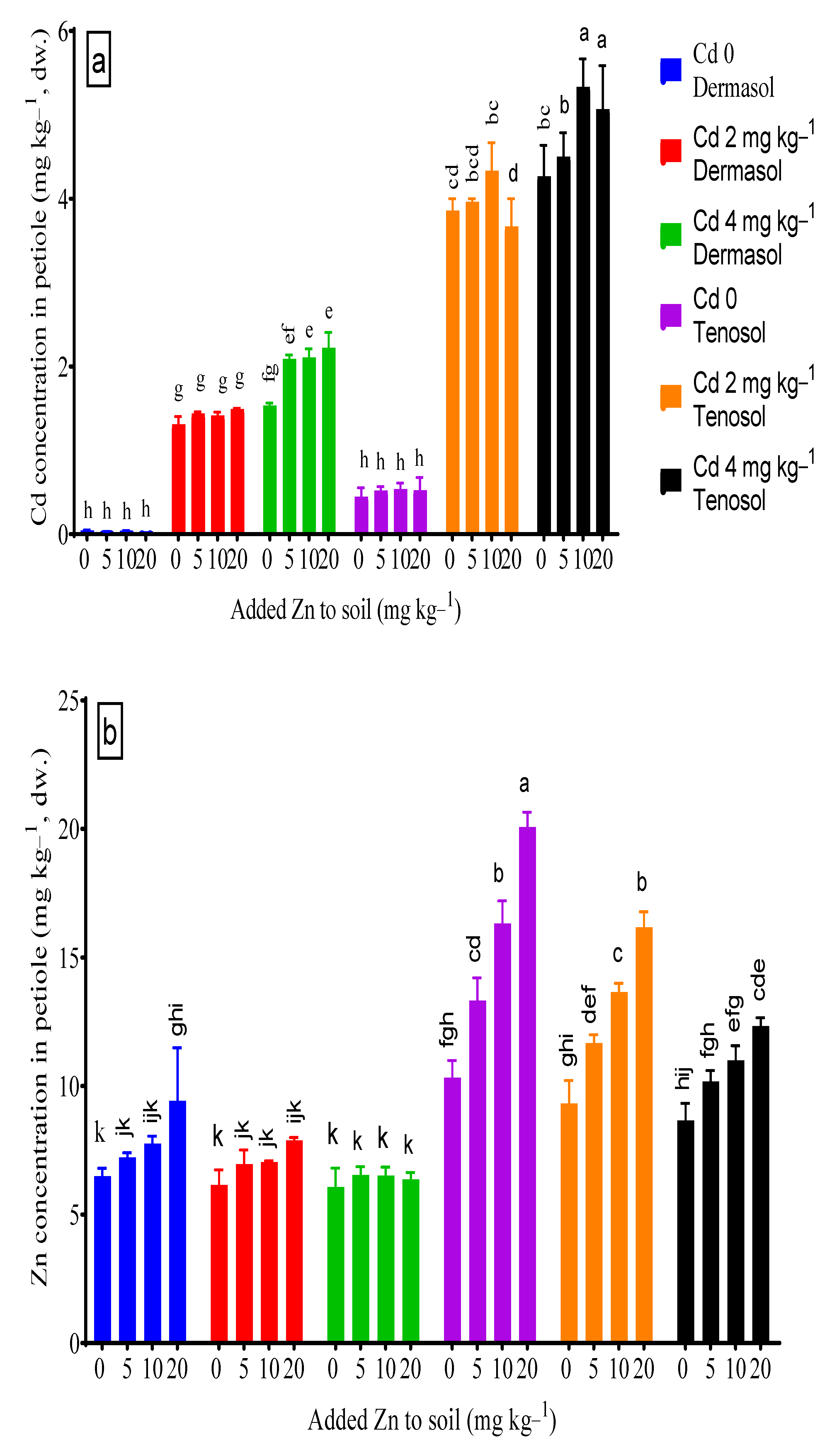
3.4. Concentrations of Cadmium and Zinc in Petiole
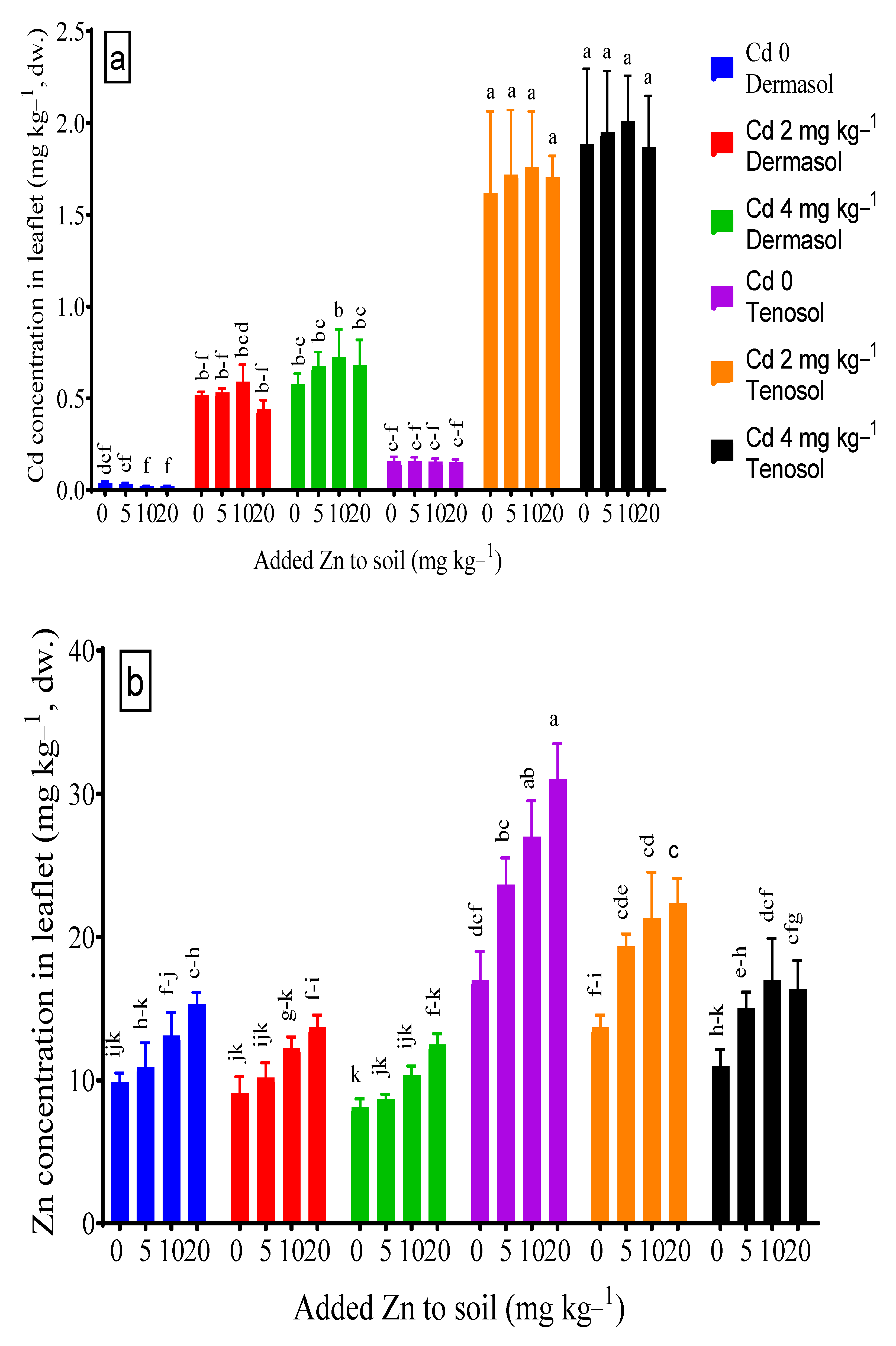
3.5. Concentrations of Cadmium and Zinc in Leaflets
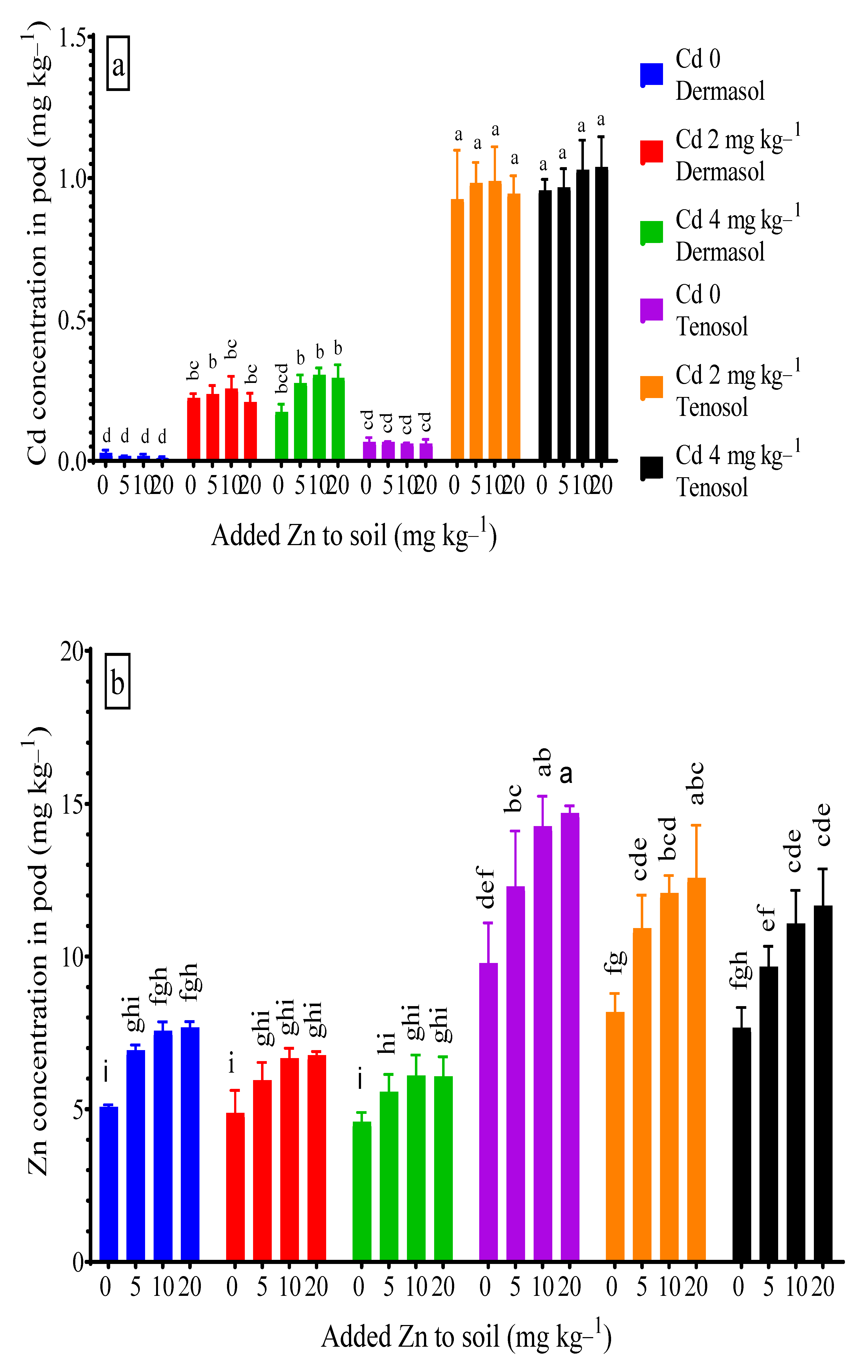
3.6. Concentration of Cadmium and Zinc in Pods
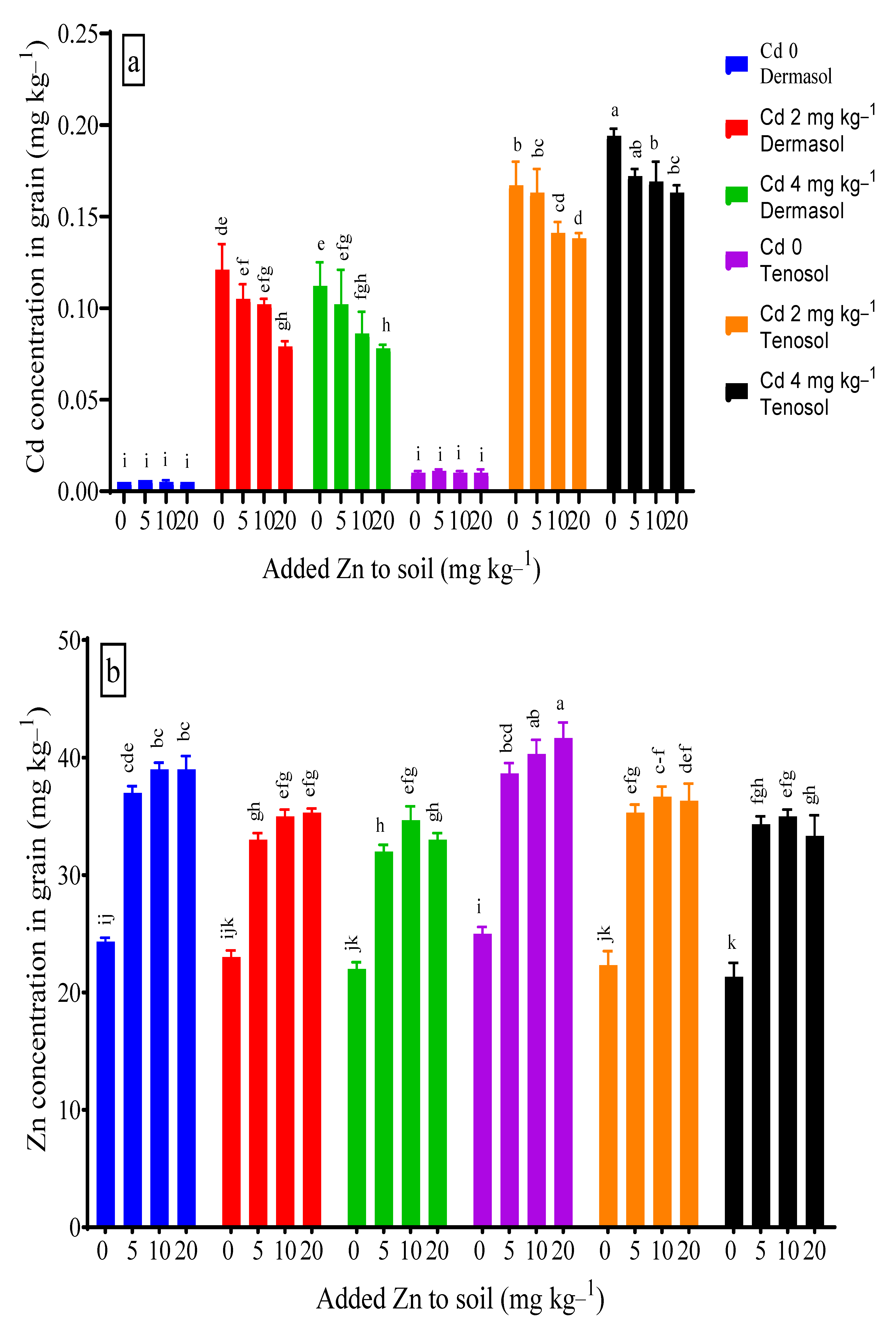
3.7. Concentrations of Cadmium and Zinc in Grains
3.8. Correlation Matrix
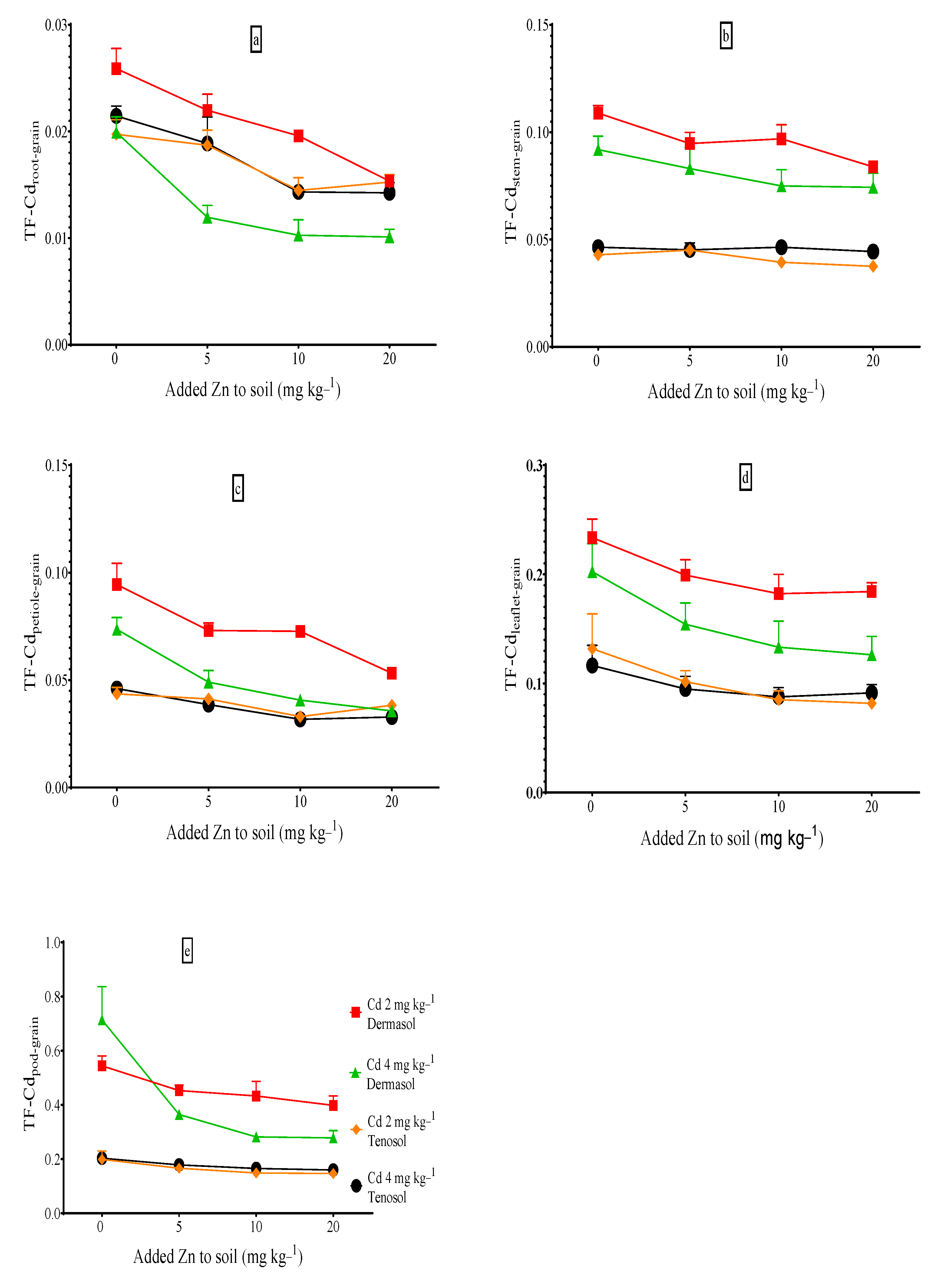
3.9. Translocation Factors (TFs) of Cadmium to Grains
3.10. Cadmium (Cd) Concentration in Pore Water
3.11. Health Risk Assessment of Cadmium Exposure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karahan, F.; Ozyigit, I.I.; Saracoglu, I.A.; Yalcin, I.E.; Ozyigit, A.H.; Ilcim, A. Heavy metal levels and mineral nutrient status in different parts of various medicinal plants collected from eastern Mediterranean region of Turkey. Biol. Trace Elem. Res. 2020, 197, 316–329. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Prasad, S.; Yadav, K.K.; Shrivastava, M.; Gupta, N.; Nagar, S.; Bach, Q.V.; Kamyab, H.; Khan, S.A.; Yadav, S. Hazardous heavy metals contamination of vegetables and food chain: Role of sustainable remediation approaches–A review. Environ. Res. 2019, 179, 108792. [Google Scholar] [CrossRef] [PubMed]
- Sevik, H.; Ozel, H.B.; Cetin, M.; Özel, H.U.; Erdem, T. Determination of changes in heavy metal accumulation depending on plant species, plant organism, and traffic density in some landscape plants. Air Qual. Atmos. Health 2019, 12, 189–195. [Google Scholar] [CrossRef]
- Schweizer, S.A.; Seitz, B.; Van Der Heijden, M.G.; Schulin, R.; Tandy, S. Impact of organic and conventional farming systems on wheat grain uptake and soil bioavailability of zinc and cadmium. Sci. Total Environ. 2018, 639, 608–616. [Google Scholar] [CrossRef]
- Xu, Z.; Dong, M.; Peng, X.; Ku, W.; Zhao, Y.; Yang, G. New insight into the molecular basis of cadmium stress responses of wild paper mulberry plant by transcriptome analysis. Ecotoxicol. Environ. Saf. 2019, 171, 301–312. [Google Scholar] [CrossRef]
- Tian, S.; Xie, R.; Wang, H.; Hu, Y.; Hou, D.; Liao, X.; Brown, P.H.; Yang, H.; Lin, X.; Labavitch, J.M. Uptake, sequestration and tolerance of cadmium at cellular levels in the hyperaccumulator plant species Sedum alfredii. J. Exp. Bot. 2017, 68, 2387–2398. [Google Scholar] [CrossRef]
- Arthur, E.; Crews, H.; Morgan, C. Optimizing plant genetic strategies for minimizing environmental contamination in the food chain: Report on the MAFF funded joint JIC/CSL Workshop held at the John Innes Centre, October 21–23, 1998. Int. J. Phytoremediat. 2000, 2, 1–21. [Google Scholar] [CrossRef]
- Zhang, G.; Fukami, M.; Sekimoto, H. Influence of cadmium on mineral concentrations and yield components in wheat genotypes differing in Cd tolerance at seedling stage. Field Crops Res. 2002, 77, 93–98. [Google Scholar] [CrossRef]
- Osma, E.; Özyiğit, İ.İ.; Leblebici, Z.; Demir, G.; Serin, M. Determination of Heavy Metal Concentrations in Tomato (Lycopersicon esculentum Miller) Grown in Different Station Types. 2012. Available online: https://acikerisim.nevsehir.edu.tr/handle/20.500.11787/4811 (accessed on 8 October 2022).
- Ozyigi, İ.; Yilmaz, S.; Dogan, I.; Sakcali, M.; Tombuloglu, G.; Demir, G.G. Detection of physiological and genotoxic damages reflecting toxicity in kalanchoe clones. Glob. Nest J. 2016, 18, 223–232. [Google Scholar]
- WHO. World Health Organization. Cadmium Review. Available online: https://www.who.int/ifcs/documents/forums/forum5/nmrcadmium.pdf (accessed on 26 July 2020).
- Altay, V.; Ozyigit, I.I. An ecological study of endemic plant Polygonum istanbulicum Keskin and its environs. Pak. J. Bot. 2013, 45, 455–459. [Google Scholar]
- Ozyigit, I.I.; Yalcin, B.; Turan, S.; Saracoglu, I.A.; Karadeniz, S.; Yalcin, I.E.; Demir, G. Investigation of heavy metal level and mineral nutrient status in widely used medicinal plants’ leaves in Turkey: Insights into health implications. Biol. Trace Elem. Res. 2018, 182, 387–406. [Google Scholar] [CrossRef]
- Antoniadis, V.; Levizou, E.; Shaheen, S.M.; Ok, Y.S.; Sebastian, A.; Baum, C.; Prasad, M.N.; Wenzel, W.W.; Rinklebe, J. Trace elements in the soil-plant interface: Phytoavailability, translocation, and phytoremediation—A review. Earth-Sci. Rev. 2017, 171, 621–645. [Google Scholar] [CrossRef]
- Nannoni, F.; Protano, G. Chemical and biological methods to evaluate the availability of heavy metals in soils of the Siena urban area (Italy). Sci. Total Environ. 2016, 568, 1–10. [Google Scholar] [CrossRef]
- Peijnenburg, W.J.; Zablotskaja, M.; Vijver, M.G. Monitoring metals in terrestrial environments within a bioavailability framework and a focus on soil extraction. Ecotoxicol. Environ. Saf. 2007, 67, 163–179. [Google Scholar] [CrossRef]
- Kalembkiewicz, J.; Sitarz-Palczak, E. Validation and Optimization of Conditions for the Sequential Extraction of Manganese from Ash. Pol. J. Environ. Stud. 2005, 14, 41–47. [Google Scholar] [CrossRef]
- Willard, L.L. Chemical Equilibria in Soils; John Wiley & Sons: Chichester, UK, 1979. [Google Scholar]
- Haisel, D.; Cyrusová, T.; Vaněk, T.; Podlipná, R. The effect of nanoparticles on the photosynthetic pigments in cadmium—Zinc interactions. Environ. Sci. Pollut. Res. 2019, 26, 4147–4151. [Google Scholar] [CrossRef]
- Cojocaru, P.; Gusiatin, Z.M.; Cretescu, I. Phytoextraction of Cd and Zn as single or mixed pollutants from soil by rape (Brassica napus). Environ. Sci. Pollut. Res. 2016, 23, 10693–10701. [Google Scholar] [CrossRef]
- Oliver, D.P.; Wilhelm, N.; Tiller, K.; McFarlane, J.; Cozens, G. Effect of soil and foliar applications of zinc on cadmium concentration in wheat grain. Aust. J. Exp. Agric. 1997, 37, 677–681. [Google Scholar] [CrossRef]
- Hart, J.J.; Welch, R.M.; Norvell, W.A.; Kochian, L.V. Transport interactions between cadmium and zinc in roots of bread and durum wheat seedlings. Physiol. Plant. 2002, 116, 73–78. [Google Scholar] [CrossRef]
- Du, J.; Zeng, J.; Ming, X.; He, Q.; Tao, Q.; Jiang, M.; Gao, S.; Li, X.; Lei, T.; Pan, Y. The presence of zinc reduced cadmium uptake and translocation in Cosmos bipinnatus seedlings under cadmium/zinc combined stress. Plant Physiol. Biochem. 2020, 151, 223–232. [Google Scholar] [CrossRef]
- Ghasemi, R.; Share, H.; Sharifi, R.; Boyd, R.S.; Rajakaruna, N. Inducing Ni sensitivity in the Ni hyperaccumulator plant Alyssum inflatum Nyárády (Brassicaceae) by transforming with CAX1, a vacuolar membrane calcium transporter. Ecol. Res. 2018, 33, 737–747. [Google Scholar] [CrossRef]
- Zou, R.; Wang, L.; Li, Y.C.; Tong, Z.; Huo, W.; Chi, K.; Fan, H. Cadmium absorption and translocation of amaranth (Amaranthus mangostanus L.) affected by iron deficiency. Environ. Pollut. 2020, 256, 113410. [Google Scholar] [CrossRef] [PubMed]
- Smilde, K.; Van Luit, B.; Van Driel, W. The extraction by soil and absorption by plants of applied zinc and cadmium. Plant Soil 1992, 143, 233–238. [Google Scholar] [CrossRef]
- Moraghan, J.T. Accumulation of cadmium and selected elements in flax seed grown on a calcareous soil. Plant Soil 1993, 150, 61–68. [Google Scholar] [CrossRef]
- Nan, Z.; Li, J.; Zhang, J.; Cheng, G. Cadmium and zinc interactions and their transfer in soil-crop system under actual field conditions. Sci. Total Environ. 2002, 285, 187–195. [Google Scholar] [CrossRef]
- Aravind, P.; Prasad, M.N.V. Cadmium-Zinc interactions in a hydroponic system using Ceratophyllum demersum L.: Adaptive ecophysiology, biochemistry and molecular toxicology. Braz. J. Plant Physiol. 2005, 17, 3–20. [Google Scholar] [CrossRef]
- McKenna, I.M.; Chaney, R.L.; Williams, F.M. The effects of cadmium and zinc interactions on the accumulation and tissue distribution of zinc and cadmium in lettuce and spinach. Environ. Pollut. 1993, 79, 113–120. [Google Scholar] [CrossRef]
- Wu, F.; Zhang, G. Alleviation of cadmium-toxicity by application of zinc and ascorbic acid in barley. J. Plant Nutr. 2002, 25, 2745–2761. [Google Scholar] [CrossRef]
- Sankaran, R.P.; Ebbs, S.D. Transport of Cd and Zn to seeds of Indian mustard (Brassica juncea) during specific stages of plant growth and development. Physiol. Plant. 2008, 132, 69–78. [Google Scholar] [CrossRef]
- Ueno, D.; Yamaji, N.; Kono, I.; Huang, C.F.; Ando, T.; Yano, M.; Ma, J.F. Gene limiting cadmium accumulation in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 16500–16505. [Google Scholar] [CrossRef]
- Kopittke, P.M.; de Jonge, M.D.; Wang, P.; McKenna, B.A.; Lombi, E.; Paterson, D.J.; Howard, D.L.; James, S.A.; Spiers, K.M.; Ryan, C.G. Laterally resolved speciation of arsenic in roots of wheat and rice using fluorescence-XANES imaging. New Phytol. 2014, 201, 1251–1262. [Google Scholar] [CrossRef]
- Asopa, P.P.; Bhatt, R.; Sihag, S.; Kothari, S.; Kachhwaha, S. Effect of cadmium on physiological parameters of cereal and millet plants—A comparative study. Int. J. Phytoremediation 2017, 19, 225–230. [Google Scholar] [CrossRef]
- Su, C.; Jiang, Y.; Li, F.; Yang, Y.; Lu, Q.; Zhang, T.; Hu, D.; Xu, Q. Investigation of subcellular distribution, physiological, and biochemical changes in Spirodela polyrhiza as a function of cadmium exposure. Environ. Exp. Bot. 2017, 142, 24–33. [Google Scholar] [CrossRef]
- Saini, S.; Kaur, N.; Pati, P.K. Phytohormones: Key players in the modulation of heavy metal stress tolerance in plants. Ecotoxicol. Environ. Saf. 2021, 223, 112578. [Google Scholar] [CrossRef] [PubMed]
- Martinoia, E.; Meyer, S.; De Angeli, A.; Nagy, R. Vacuolar transporters in their physiological context. Annu. Rev. Plant Biol. 2012, 63, 183–213. [Google Scholar] [CrossRef]
- Mathys, W. The role of malate, oxalate, and mustard oil glucosides in the evolution of zinc-resistance in herbage plants. Physiol. Plant. 1977, 40, 130–136. [Google Scholar] [CrossRef]
- Bouyoucos, G.J. Hydrometer method improved for making particle size analyses of soils 1. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Sumner, M.E.; Miller, W.P. Cation exchange capacity and exchange coefficients. Methods Soil Anal. Part 3 Chem. Methods 1996, 5, 1201–1229. [Google Scholar]
- Lu, X.; Cui, J.; Tian, X.; Ogunniyi, J.E.; Gale, W.J.; Zhao, A. Effects of zinc fertilization on zinc dynamics in potentially zinc-deficient calcareous soil. Agron. J. 2012, 104, 963–969. [Google Scholar] [CrossRef]
- Duan, G.; Shao, G.; Tang, Z.; Chen, H.; Wang, B.; Tang, Z.; Yang, Y.; Liu, Y.; Zhao, F.J. Genotypic and environmental variations in grain cadmium and arsenic concentrations among a panel of high yielding rice cultivars. Rice 2017, 10, 1–13. [Google Scholar] [CrossRef]
- Liu, X.; Tian, G.; Jiang, D.; Zhang, C.; Kong, L. Cadmium (Cd) distribution and contamination in Chinese paddy soils on national scale. Environ. Sci. Pollut. Res. 2016, 23, 17941–17952. [Google Scholar] [CrossRef]
- Honma, T.; Ohba, H.; Kaneko-Kadokura, A.; Makino, T.; Nakamura, K.; Katou, H. Optimal soil Eh, pH, and water management for simultaneously minimizing arsenic and cadmium concentrations in rice grains. Environ. Sci. Technol. 2016, 50, 4178–4185. [Google Scholar] [CrossRef]
- AMA. Australian Mungbean Association (AMA). Variety Factsheet. 2015. Available online: http://www.mungbean.org.au/varieties.html (accessed on 8 October 2022).
- Rahman, M.M.; Dong, Z.; Naidu, R. Concentrations of arsenic and other elements in groundwater of Bangladesh and West Bengal, India: Potential cancer risk. Chemosphere 2015, 139, 54–64. [Google Scholar] [CrossRef]
- Nisha, U.S.; Khan, M.S.I.; Prodhan, M.D.H.; Meftaul, I.M.; Begum, N.; Parven, A.; Shahriar, S.; Juraimi, A.S.; Hakim, M.A. Quantification of pesticide residues in fresh vegetables available in local markets for human consumption and the associated health risks. Agronomy 2021, 11, 1804. [Google Scholar] [CrossRef]
- Parven, A.; Khan, M.S.I.; Prodhan, M.D.H.; Venkateswarlu, K.; Megharaj, M.; Meftaul, I.M. Human health risk assessment through quantitative screening of insecticide residues in two green beans to ensure food safety. J. Food Compos. Anal. 2021, 103, 104121. [Google Scholar] [CrossRef]
- Rashid, M.H.; Rahman, M.M.; Correll, R.; Naidu, R. Arsenic and other elemental concentrations in mushrooms from Bangladesh: Health risks. Int. J. Environ. Res. Public Health 2018, 15, 919. [Google Scholar] [CrossRef]
- Varma, P. The Global Economy of Pulses; Vikas, R., Dorian, K.N., Eds.; SAGE Publications Sage India: New Delhi, India, 2019. [Google Scholar]
- Afrin, S.; Alam, M.K.; Ahmed, M.W.; Parven, A.; Jubayer, M.F.; Megharaj, M.; Meftaul, I.M.; Khan, M.S.I. Determination and probabilistic health risk assessment of heavy metals in widely consumed market basket fruits from Dhaka city Bangladesh. Int. J. Environ. Anal. Chem. 2021, 1–16. [Google Scholar] [CrossRef]
- Hossain, M.S.; Ahmed, F.; Abdullah, A.T.M.; Akbor, M.A.; Ahsan, M.A. Public health risk assessment of heavy metal uptake by vegetables grown at a waste-water-irrigated site in Dhaka, Bangladesh. J. Health Pollut. 2015, 5, 78–85. [Google Scholar] [CrossRef]
- Sultana, R.; Chamon, A.; Mondol, M. Heavy metal concentration in commonly sold stem vegetables in Dhaka city market and probable health risk. Dhaka Univ. J. Biol. Sci. 2021, 30, 221–232. [Google Scholar] [CrossRef]
- International Agency for Reseach on Cancer (IARC). Cadmium and Cadmium Compounds. In Monograph on the Evaluation of Carcinogenic Risks to Humans; Review of Human Carcinogens: Arsenic, Metals, Fibre and Dusts; International Agency for Reseach on Cancer: Lyon, France, 2012; Volume 58, pp. 119–237. [Google Scholar]
- Kasozi, K.I.; Hamira, Y.; Zirintunda, G.; Alsharif, K.F.; Altalbawy, F.M.; Ekou, J.; Tamale, A.; Matama, K.; Ssempijja, F.; Muyinda, R. Descriptive analysis of heavy metals content of beef from eastern Uganda and their safety for public consumption. Front. Nutr. 2021, 8, 592340. [Google Scholar] [CrossRef]
- Pirsaheb, M.; Hadei, M.; Sharafi, K. Human health risk assessment by Monte Carlo simulation method for heavy metals of commonly consumed cereals in Iran-Uncertainty and sensitivity analysis. J. Food Compos. Anal. 2021, 96, 103697. [Google Scholar] [CrossRef]
- Kusin, F.M.; Azani, N.N.M.; Hasan, S.N.M.S.; Sulong, N.A. Distribution of heavy metals and metalloid in surface sediments of heavily-mined area for bauxite ore in Pengerang, Malaysia and associated risk assessment. Catena 2018, 165, 454–464. [Google Scholar] [CrossRef]
- Kubier, A.; Pichler, T. Cadmium in groundwater−A synopsis based on a large hydrogeochemical data set. Sci. Total Environ. 2019, 689, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.; Rahman, M.M.; Halim, M.A.; Naidu, R. Growth, metal partitioning and antioxidant enzyme activities of mung beans as influenced by zinc oxide nanoparticles under cadmium stress. Crop Pasture Sci. 2022, 73, 862–876. [Google Scholar] [CrossRef]
- Sarwar, N.; Malhi, S.S.; Zia, M.H.; Naeem, A.; Bibi, S.; Farid, G. Role of mineral nutrition in minimizing cadmium accumulation by plants. J. Sci. Food Agric. 2010, 90, 925–937. [Google Scholar] [CrossRef]
- Ismael, M.A.; Elyamine, A.M.; Moussa, M.G.; Cai, M.; Zhao, X.; Hu, C. Cadmium in plants: Uptake, toxicity, and its interactions with selenium fertilizers. Metallomics 2019, 11, 255–277. [Google Scholar] [CrossRef]
- Yang, Y.; Ge, Y.; Zeng, H.; Zhou, X.; Peng, L.; Zeng, Q. Phytoextraction of cadmium-contaminated soil and potential of regenerated tobacco biomass for recovery of cadmium. Sci. Rep. 2017, 7, 7210. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; ur Rehman, M.Z.; Rinklebe, J.; Tsang, D.C.; Bashir, A.; Maqbool, A.; Tack, F.; Ok, Y.S. Cadmium phytoremediation potential of Brassica crop species: A review. Sci. Total Environ. 2018, 631, 1175–1191. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Abbas, T.; Zia-ur-Rehman, M.; Hannan, F.; Keller, C.; Al-Wabel, M.I.; Ok, Y.S. Cadmium minimization in wheat: A critical review. Ecotoxicol. Environ. Saf. 2016, 130, 43–53. [Google Scholar] [CrossRef]
- Cakmak, I.; Kutman, U. Agronomic biofortification of cereals with zinc: A review. Eur. J. Soil Sci. 2018, 69, 172–180. [Google Scholar] [CrossRef]
- Huang, G.; Ding, C.; Zhou, Z.; Zhang, T.; Wang, X. A tillering application of zinc fertilizer based on basal stabilization reduces Cd accumulation in rice (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2019, 167, 338–344. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, B.; Liu, H.; Liang, X.; Ma, W.; Shi, Z.; Yang, S. Zinc effects on cadmium toxicity in two wheat varieties (Triticum aestivum L.) differing in grain cadmium accumulation. Ecotox. Environ. Safe 2019, 183, 109562. [Google Scholar] [CrossRef]
- Farooq, M.; Ullah, A.; Usman, M.; Siddique, K.H. Application of zinc and biochar help to mitigate cadmium stress in bread wheat raised from seeds with high intrinsic zinc. Chemosphere 2020, 260, 127652. [Google Scholar] [CrossRef]
- Praharaj, S.; Skalicky, M.; Maitra, S.; Bhadra, P.; Shankar, T.; Brestic, M.; Hejnak, V.; Vachova, P.; Hossain, A. Zinc biofortification in food crops could alleviate the zinc malnutrition in human health. Molecules 2021, 26, 3509. [Google Scholar] [CrossRef]
- Dhaliwal, S.S.; Sharma, V.; Shukla, A.K.; Verma, V.; Kaur, M.; Shivay, Y.S.; Nisar, S.; Gaber, A.; Brestic, M.; Barek, V. Biofortification—A Frontier Novel Approach to Enrich Micronutrients in Field Crops to Encounter the Nutritional Security. Molecules 2022, 27, 1340. [Google Scholar] [CrossRef]
- Haider, M.U.; Hussain, M.; Farooq, M.; Nawaz, A. Zinc nutrition for improving the productivity and grain biofortification of mungbean. J. Soil Sci. Plant Nutr. 2020, 20, 1321–1335. [Google Scholar] [CrossRef]
- Hussain, M.; Shahid, M.Z.; Mehboob, N.; Minhas, W.A.; Akram, M. Zinc application improves growth, yield and grain zinc concentration of mung bean (Vigna radiata L.). Semin. Ciências Agrárias 2021, 42, 487–500. [Google Scholar] [CrossRef]
- Rengel, Z.; Batten, G.; Crowley, D.d. Agronomic approaches for improving the micronutrient density in edible portions of field crops. Field Crops Res. 1999, 60, 27–40. [Google Scholar] [CrossRef]
- Rehman, A.; Farooq, M.; Naveed, M.; Nawaz, A.; Shahzad, B. Seed priming of Zn with endophytic bacteria improves the productivity and grain biofortification of bread wheat. Eur. J. Agron. 2018, 94, 98–107. [Google Scholar] [CrossRef]
- Rehman, A.; Farooq, M.; Naveed, M.; Ozturk, L.; Nawaz, A. Pseudomonas-aided zinc application improves the productivity and biofortification of bread wheat. Crop Pasture Sci. 2018, 69, 659–672. [Google Scholar] [CrossRef]
- Rehman, A.; Farooq, M.; Nawaz, A.; Al-Sadi, A.M.; Al-Hashmi, K.S.; Nadeem, F.; Ullah, A. Characterizing bread wheat genotypes of Pakistani origin for grain zinc biofortification potential. J. Sci. Food Agric. 2018, 98, 4824–4836. [Google Scholar] [CrossRef] [PubMed]
- Nazar, R.; Iqbal, N.; Masood, A.; Khan, M.I.R.; Syeed, S.; Khan, N.A. Cadmium toxicity in plants and role of mineral nutrients in its alleviation. Am. J. Plant Sci. 2012, 3, 24162. [Google Scholar] [CrossRef]
- Xu, Z.M.; Li, Q.S.; Yang, P.; Ye, H.J.; Chen, Z.S.; Guo, S.H.; Wang, L.L.; He, B.Y.; Zeng, E.Y. Impact of osmoregulation on the differences in Cd accumulation between two contrasting edible amaranth cultivars grown on Cd-polluted saline soils. Environ. Pollut. 2017, 224, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Malar, S.; Shivendra Vikram, S.; Favas, P.; Perumal, V. Lead heavy metal toxicity induced changes on growth and antioxidative enzymes level in water hyacinths [Eichhornia crassipes (Mart.)]. Bot. Stud. 2016, 55, 54. [Google Scholar] [CrossRef]
- García-Gómez, C.; García, S.; Obrador, A.F.; González, D.; Babín, M.; Fernández, M.D. Effects of aged ZnO NPs and soil type on Zn availability, accumulation and toxicity to pea and beet in a greenhouse experiment. Ecotox. Environ. Safe 2018, 160, 222–230. [Google Scholar] [CrossRef]
- Kailasam, S.; Peiter, E. A path toward concurrent biofortification and cadmium mitigation in plant-based foods. New Phytol. 2021, 232, 17–24. [Google Scholar] [CrossRef]
- Zare, A.; Khoshgoftarmanesh, A.; Malakouti, M.; Bahrami, H.; Chaney, R. Root uptake and shoot accumulation of cadmium by lettuce at various Cd: Zn ratios in nutrient solution. Ecotoxicol. Environ. Saf. 2018, 148, 441–446. [Google Scholar] [CrossRef]
- Cakmak, I.; Welch, R.; Erenoglu, B.; Römheld, V.; Norvell, W.; Kochian, L. Influence of varied zinc supply on re-translocation of cadmium (109Cd) and rubidium (86Rb) applied on mature leaf of durum wheat seedlings. Plant Soil 2000, 219, 279–284. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, J.; Christie, P.; Zhang, F. Influence of external zinc and phosphorus supply on Cd uptake by rice (Oryza sativa L.) seedlings with root surface iron plaque. Plant Soil 2007, 300, 105–115. [Google Scholar] [CrossRef]
- Hart, J.J.; Welch, R.M.; Norvell, W.A.; Clarke, J.M.; Kochian, L.V. Zinc effects on cadmium accumulation and partitioning in near-isogenic lines of durum wheat that differ in grain cadmium concentration. New Phytol. 2005, 167, 391–401. [Google Scholar] [CrossRef]
- Welch, R.; Hart, J.; Norvell, W.; Sullivan, L.; Kochian, L. Effects of nutrient solution zinc activity on net uptake, translocation, and root export of cadmium and zinc by separated sections of intact durum wheat (Triticum turgidum L. var durum) seedling roots. Plant Soil 1999, 208, 243–250. [Google Scholar] [CrossRef]
- Seth, C.S.; Remans, T.; Keunen, E.; Jozefczak, M.; Gielen, H.; Opdenakker, K.; Weyens, N.; Vangronsveld, J.; Cuypers, A. Phytoextraction of toxic metals: A central role for glutathione. Plant Cell Environ. 2012, 35, 334–346. [Google Scholar] [CrossRef]
- Rhaman, M.S.; Imran, S.; Rauf, F.; Khatun, M.; Baskin, C.C.; Murata, Y.; Hasanuzzaman, M. Seed priming with phytohormones: An effective approach for the mitigation of abiotic stress. Plants 2020, 10, 37. [Google Scholar] [CrossRef]
- Lepp, N. Effect of Heavy Metal Pollution on Plants; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1981; Volumes 1 and 2. [Google Scholar]
- Alloway, B.J. Cadmium. In Heavy Metals in Soils; Alloway, B.J., Ed.; Blackie: London, UK, 1990; pp. 100–124. [Google Scholar]
- Siddique, A.B.; Rahman, M.M.; Islam, M.R.; Mondal, D.; Naidu, R. Response of Iron and Cadmium on Yield and Yield Components of Rice and Translocation in Grain: Health Risk Estimation. Front. Environ. Sci. 2021, 9, 716770. [Google Scholar] [CrossRef]
- Wang, Y.M.; Liu, Q.; Li, M.; Yuan, X.Y.; Uchimiya, M.; Wang, S.W.; Zhang, Z.Y.; Ji, T.; Wang, Y.; Zhao, Y.Y. Rhizospheric pore-water content predicts the biochar-attenuated accumulation, translocation, and toxicity of cadmium to lettuce. Ecotox. Environ. Safe 2021, 208, 111675. [Google Scholar] [CrossRef]
- Gao, X.; Flaten, D.N.; Tenuta, M.; Grimmett, M.G.; Gawalko, E.J.; Grant, C.A. Soil solution dynamics and plant uptake of cadmium and zinc by durum wheat following phosphate fertilization. Plant Soil 2011, 338, 423–434. [Google Scholar] [CrossRef]
- Beesley, L.; Dickinson, N. Carbon and trace element mobility in an urban soil amended with green waste compost. J. Soils Sediments 2010, 10, 215–222. [Google Scholar] [CrossRef]
- Meers, E.; Du Laing, G.; Unamuno, V.; Ruttens, A.; Vangronsveld, J.; Tack, F.M.; Verloo, M.G. Comparison of cadmium extractability from soils by commonly used single extraction protocols. Geoderma 2007, 141, 247–259. [Google Scholar] [CrossRef]
- ATSDR. Toxicological Profile for Cadmium, Agency for Toxic Substances and Disease Registry (ATSDR) Toxicological Profiles; Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2012. [Google Scholar]
- EFSA. EFSA Panel on Contaminants in the Food Chain. Statement on Tolerable Weekly Intake for Cadmium. EFSA J. 2011, 9, 1975. [Google Scholar] [CrossRef]
- JECFA. Joint FAO/WHO Expert Committee on Food Additives, Seventy-Third Meeting. Geneva, 8–17 June 2010. Summary and Conclusions. JECFA/73/SC; Food and Agriculture Organization of the United Nations, WHO: Geneva, Switzerland, 2010. [Google Scholar]
- Zhou, H.; Yang, W.T.; Zhou, X.; Liu, L.; Gu, J.F.; Wang, W.L.; Zou, J.L.; Tian, T.; Peng, P.Q.; Liao, B.H. Accumulation of heavy metals in vegetable species planted in contaminated soils and the health risk assessment. Int. J. Environ. Res. Public Health 2016, 13, 289. [Google Scholar] [CrossRef]
- Wei, J.; Cen, K. Contamination and health risk assessment of heavy metals in cereals, legumes, and their products: A case study based on the dietary structure of the residents of Beijing, China. J. Clean. Prod. 2020, 260, 121001. [Google Scholar] [CrossRef]
- Wei, J.; Cen, K. Assessment of human health risk based on characteristics of potential toxic elements (PTEs) contents in foods sold in Beijing, China. Sci. Total Environ. 2020, 703, 134747. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Types | pH (H2O) | EC µs/cm | %C | %N | %S | Texture | % Sand | % Silt | % Clay | Cd (mg kg−1) | Zn (mg kg−1) | DTPA Zn (mg kg−1) | As (mg kg−1) | Pb (mg kg−1) | Cr (mg kg−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dermosol | 7.7 | 161.2 | 2.7 | 0.21 | 0.055 | Silt loam | 33.8 | 55 | 11.2 | 0.01 | 5.9 | 0.5 | 1.6 | 5.2 | 9.1 |
| Tenosol | 6.4 | 190 | 1.4 | 0.22 | 0.009 | Sandy loam | 63.8 | 23.8 | 12.5 | 0.05 | 37 | 5 | 6.2 | 8.8 | 6.2 |
| Parameters | Root Zn | Root Cd | Stem Zn | Stem Cd | Petiole Zn | Petiole Cd | Pod Zn | Pod Cd | Leaflet Zn | Leaflet Cd | Grain Zn | Grain Cd |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Root Zn | 1.00 | |||||||||||
| Root Cd | 0.21 | 1.00 | ||||||||||
| Stem Zn | 0.887 ** | 0.03 | 1.00 | |||||||||
| Stem Cd | 0.414 ** | 0.815 ** | 0.269 * | 1.00 | ||||||||
| Petiole Zn | 0.808 ** | −0.028 | 0.898 ** | 0.229 | 1.00 | |||||||
| Petiole Cd | 0.403 ** | 0.913 ** | 0.226 | 0.925 ** | 0.156 | 1.00 | ||||||
| Pod Zn | 0.801 ** | 0.035 | 0.842 ** | 0.281 * | 0.862 ** | 0.235 * | 1.00 | |||||
| Pod Cd | 0.441 ** | 0.829 ** | 0.266 * | 0.927 ** | 0.219 | 0.949 ** | 0.281 * | 1.00 | ||||
| Leaflet Zn | 0.746 ** | −0.158 | 0.860 ** | 0.090 | 0.862 ** | 0.025 | 0.833 ** | 0.072 | 1.00 | |||
| Leaflet Cd | 0.399 ** | 0.833 ** | 0.244 * | 0.896 ** | 0.179 | 0.938 ** | 0.255 * | 0.908 ** | 0.033 | 1.00 | ||
| Grain Zn | 0.552 ** | −0.341 ** | 0.597 ** | −0.193 | 0.618 ** | −0.188 | 0.641 ** | −0.125 | 0.655 ** | −0.153 | 1.00 | |
| Grain Cd | 0.165 | 0.882 ** | −0.002 | 0.889 ** | −0.058 | 0.896 ** | −0.012 | 0.861 ** | −0.188 | 0.846 ** | −0.384 ** | 1.00 |
| Soil Types | Zn (mg kg−1) | Cd (mg kg−1) | Grain Cd (mg kg−1) | EDI × 10−3 | THQ | ILCR × 10−4 | |||
|---|---|---|---|---|---|---|---|---|---|
| Children | Adults | Children | Adults | Children | Adults | ||||
| Tenosol | 0 | 0 | 0.0100 | 0.0084 | 0.0027 | 0.0084 | 0.0027 | 0.5292 | 0.1694 |
| 5 | 0 | 0.0107 | 0.0090 | 0.0029 | 0.0090 | 0.0029 | 0.5647 | 0.1807 | |
| 10 | 0 | 0.0103 | 0.0087 | 0.0028 | 0.0087 | 0.0028 | 0.5467 | 0.1750 | |
| 20 | 0 | 0.0103 | 0.0087 | 0.0028 | 0.0087 | 0.0028 | 0.5467 | 0.1750 | |
| 0 | 2 | 0.1667 | 0.1400 | 0.0448 | 0.1400 | 0.0448 | 8.8202 | 2.8234 | |
| 5 | 2 | 0.1633 | 0.1372 | 0.0439 | 0.1372 | 0.0439 | 8.6434 | 2.7668 | |
| 10 | 2 | 0.1413 | 0.1187 | 0.0380 | 0.1187 | 0.0380 | 7.4792 | 2.3941 | |
| 20 | 2 | 0.1377 | 0.1156 | 0.0370 | 0.1156 | 0.0370 | 7.2855 | 2.3321 | |
| 0 | 4 | 0.1937 | 0.1627 | 0.0521 | 0.1627 | 0.0521 | 10.2490 | 3.2807 | |
| 5 | 4 | 0.1717 | 0.1442 | 0.0462 | 0.1442 | 0.0462 | 9.0848 | 2.9081 | |
| 10 | 4 | 0.1690 | 0.1420 | 0.0454 | 0.1420 | 0.0454 | 8.9435 | 2.8628 | |
| 20 | 4 | 0.1633 | 0.1372 | 0.0439 | 0.1372 | 0.0439 | 8.6434 | 2.7668 | |
| Dermosol | 0 | 0 | 0.0050 | 0.0042 | 0.0013 | 0.0042 | 0.0013 | 0.2646 | 0.0847 |
| 5 | 0 | 0.0057 | 0.0048 | 0.0015 | 0.0048 | 0.0015 | 0.3001 | 0.0960 | |
| 10 | 0 | 0.0053 | 0.0045 | 0.0014 | 0.0045 | 0.0014 | 0.2821 | 0.0903 | |
| 20 | 0 | 0.0050 | 0.0042 | 0.0013 | 0.0042 | 0.0013 | 0.2646 | 0.0847 | |
| 0 | 2 | 0.1207 | 0.1014 | 0.0324 | 0.1014 | 0.0324 | 6.3859 | 2.0441 | |
| 5 | 2 | 0.1050 | 0.0882 | 0.0282 | 0.0882 | 0.0282 | 5.5566 | 1.7787 | |
| 10 | 2 | 0.1020 | 0.0857 | 0.0274 | 0.0857 | 0.0274 | 5.3978 | 1.7279 | |
| 20 | 2 | 0.0793 | 0.0666 | 0.0213 | 0.0666 | 0.0213 | 4.1981 | 1.3438 | |
| 0 | 4 | 0.1120 | 0.0941 | 0.0301 | 0.0941 | 0.0301 | 5.9270 | 1.8973 | |
| 5 | 4 | 0.1020 | 0.0857 | 0.0274 | 0.0857 | 0.0274 | 5.3978 | 1.7279 | |
| 10 | 4 | 0.0863 | 0.0725 | 0.0232 | 0.0725 | 0.0232 | 4.5686 | 1.4624 | |
| 20 | 4 | 0.0777 | 0.0652 | 0.0209 | 0.0652 | 0.0209 | 4.1103 | 1.3157 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rashid, M.H.; Rahman, M.M.; Naidu, R. Zinc Biofortification through Basal Zinc Supply Reduces Grain Cadmium in Mung Beans: Metal Partitioning and Health Risks Assessment. Toxics 2022, 10, 689. https://doi.org/10.3390/toxics10110689
Rashid MH, Rahman MM, Naidu R. Zinc Biofortification through Basal Zinc Supply Reduces Grain Cadmium in Mung Beans: Metal Partitioning and Health Risks Assessment. Toxics. 2022; 10(11):689. https://doi.org/10.3390/toxics10110689
Chicago/Turabian StyleRashid, Md Harunur, Mohammad Mahmudur Rahman, and Ravi Naidu. 2022. "Zinc Biofortification through Basal Zinc Supply Reduces Grain Cadmium in Mung Beans: Metal Partitioning and Health Risks Assessment" Toxics 10, no. 11: 689. https://doi.org/10.3390/toxics10110689
APA StyleRashid, M. H., Rahman, M. M., & Naidu, R. (2022). Zinc Biofortification through Basal Zinc Supply Reduces Grain Cadmium in Mung Beans: Metal Partitioning and Health Risks Assessment. Toxics, 10(11), 689. https://doi.org/10.3390/toxics10110689