
Arbuscular Mycorrhizal Fungal Inoculation Increases Organic Selenium Accumulation in Soybean (Glycine max (Linn.) Merr.) Growing in Selenite-Spiked Soils

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Pot Experiment
2.2. Measurement of Plant Dry Weights (DWs) and AMF Mycorrhizal Colonization in Roots
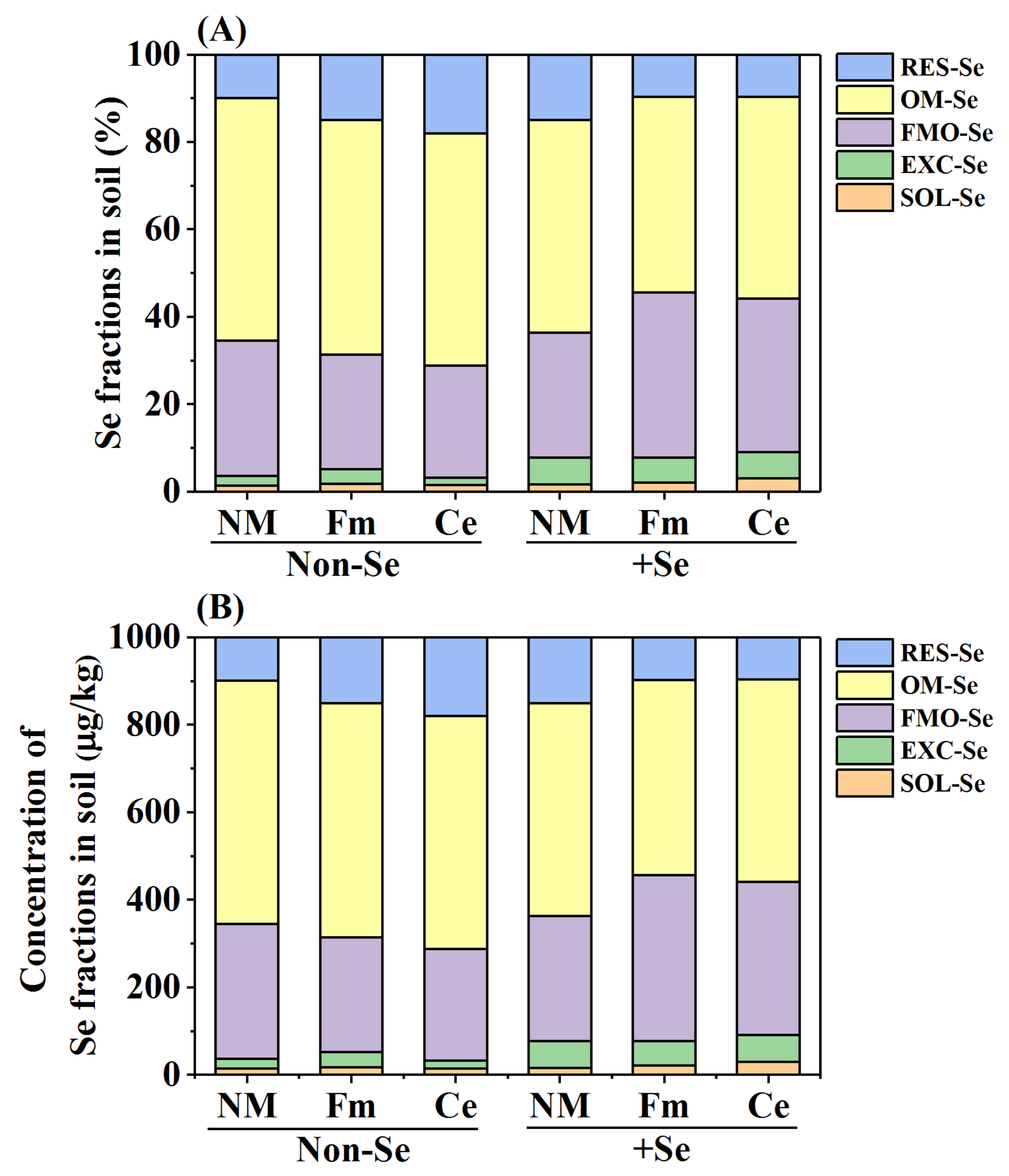
2.3. Determination of Soil Se Fractions
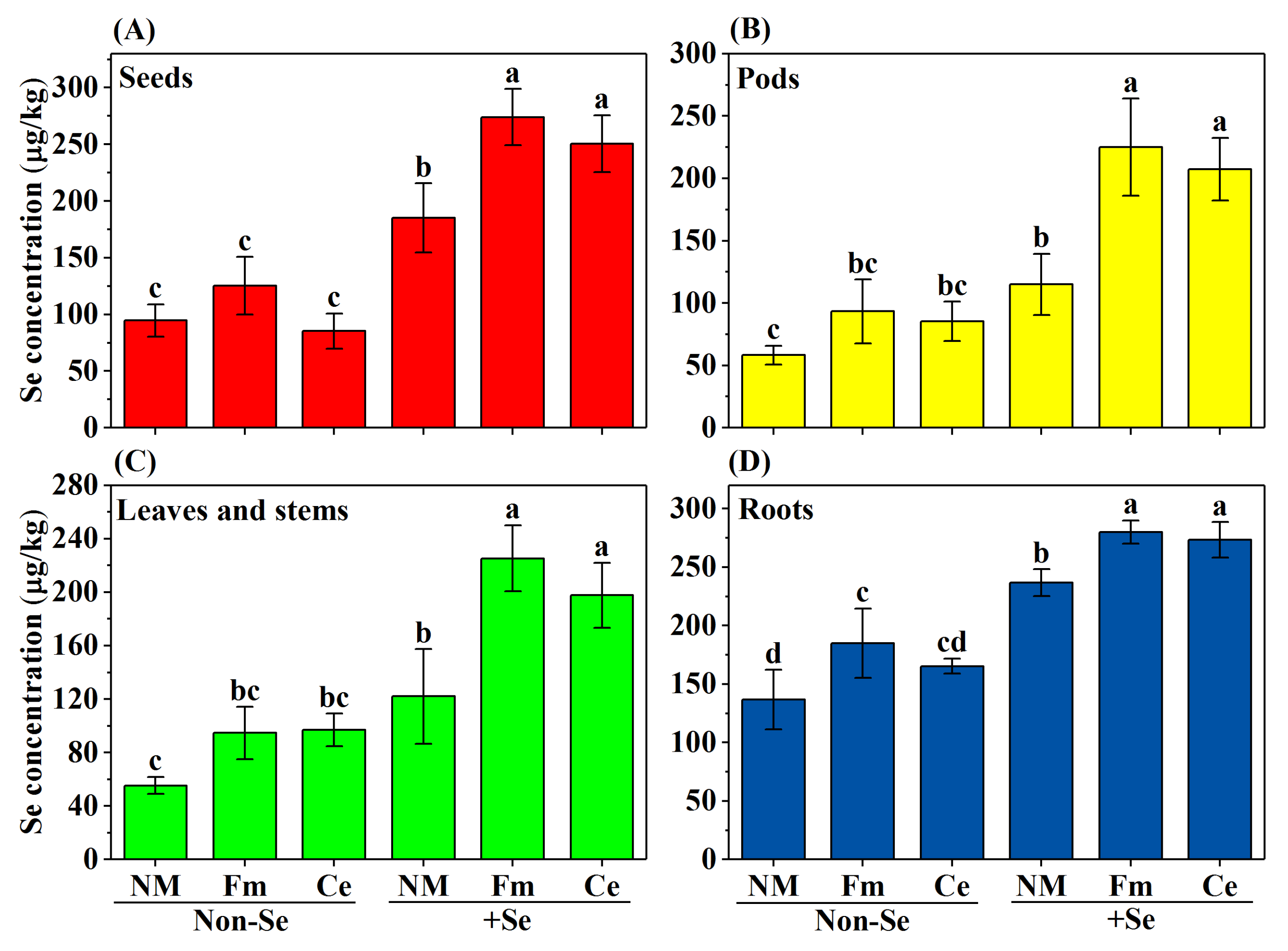
2.4. Determination of Se Concentration in Soybean
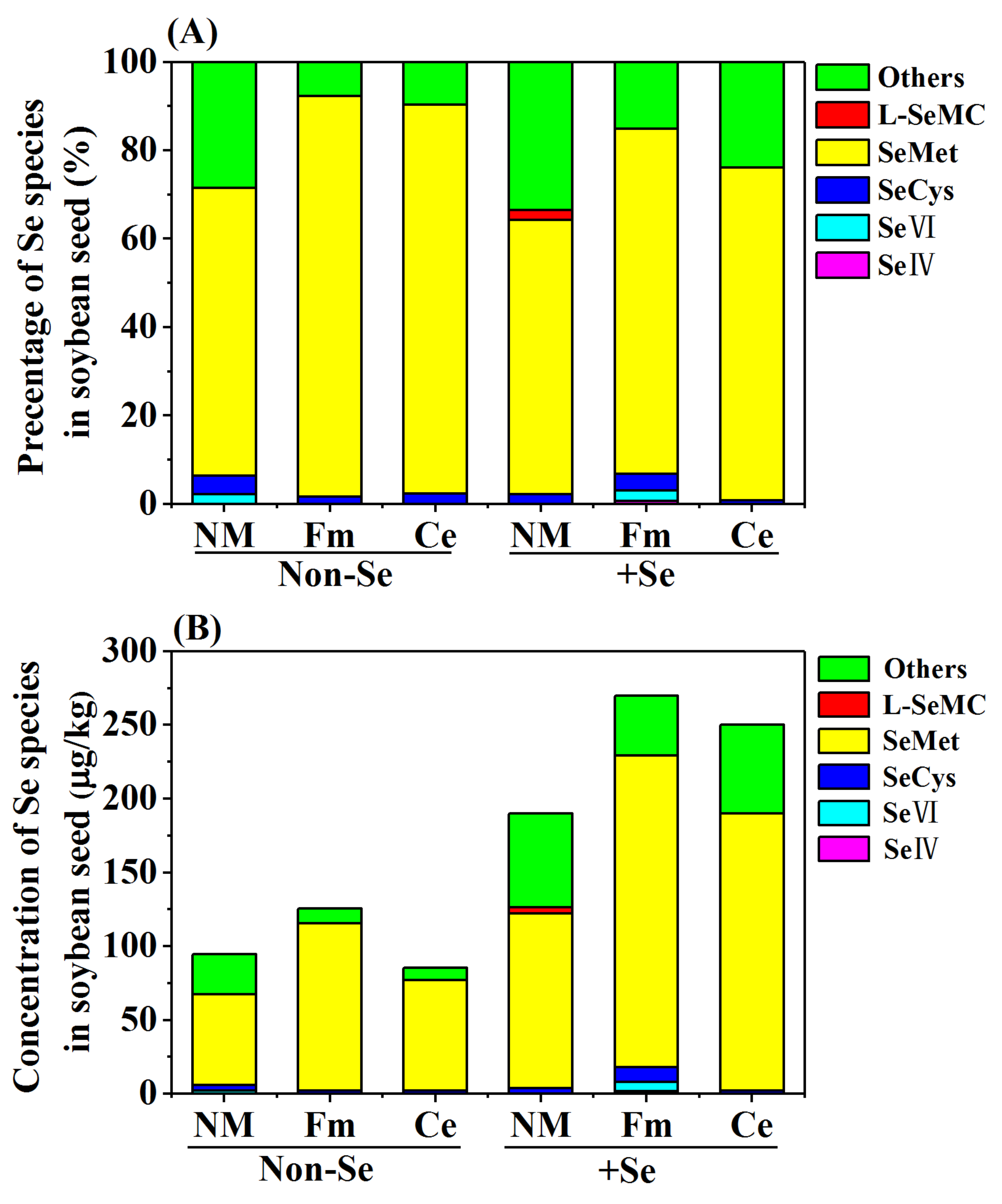
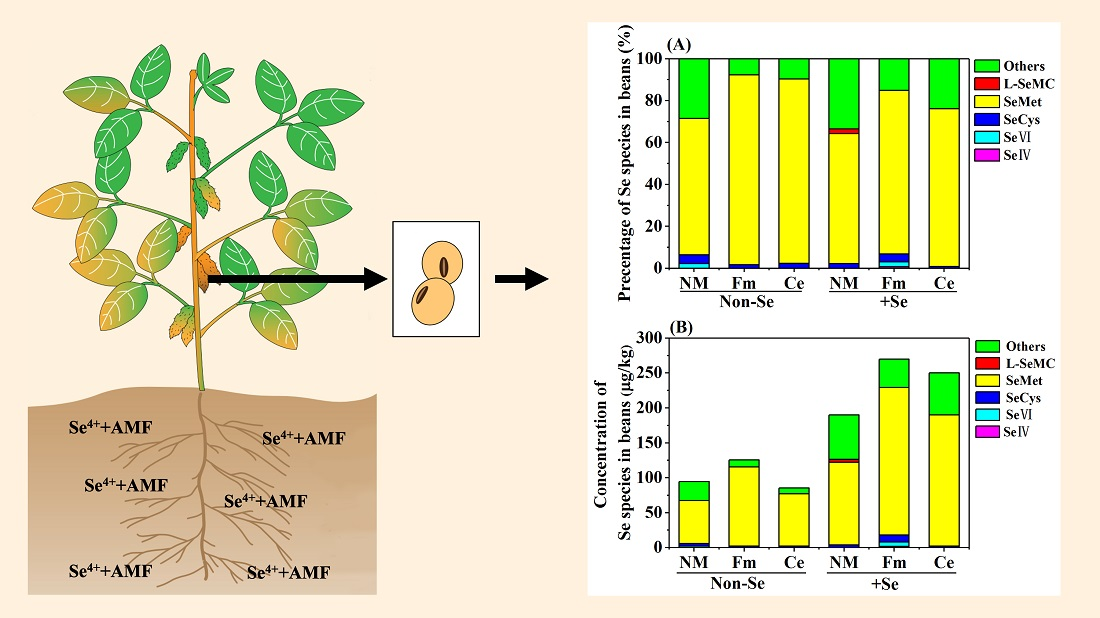
2.5. Analysis of Se Speciation in Soybean Seed
2.6. Calculation of Bioaccumulation Factors (BAFs) and Translocation Coefficients (TCs)
2.7. Se Biofortification Potentiality Analysis
2.8. Statistical Analysis
3. Results
3.1. Root Colonization and Plant Development
3.2. Fractions of Se in Soil
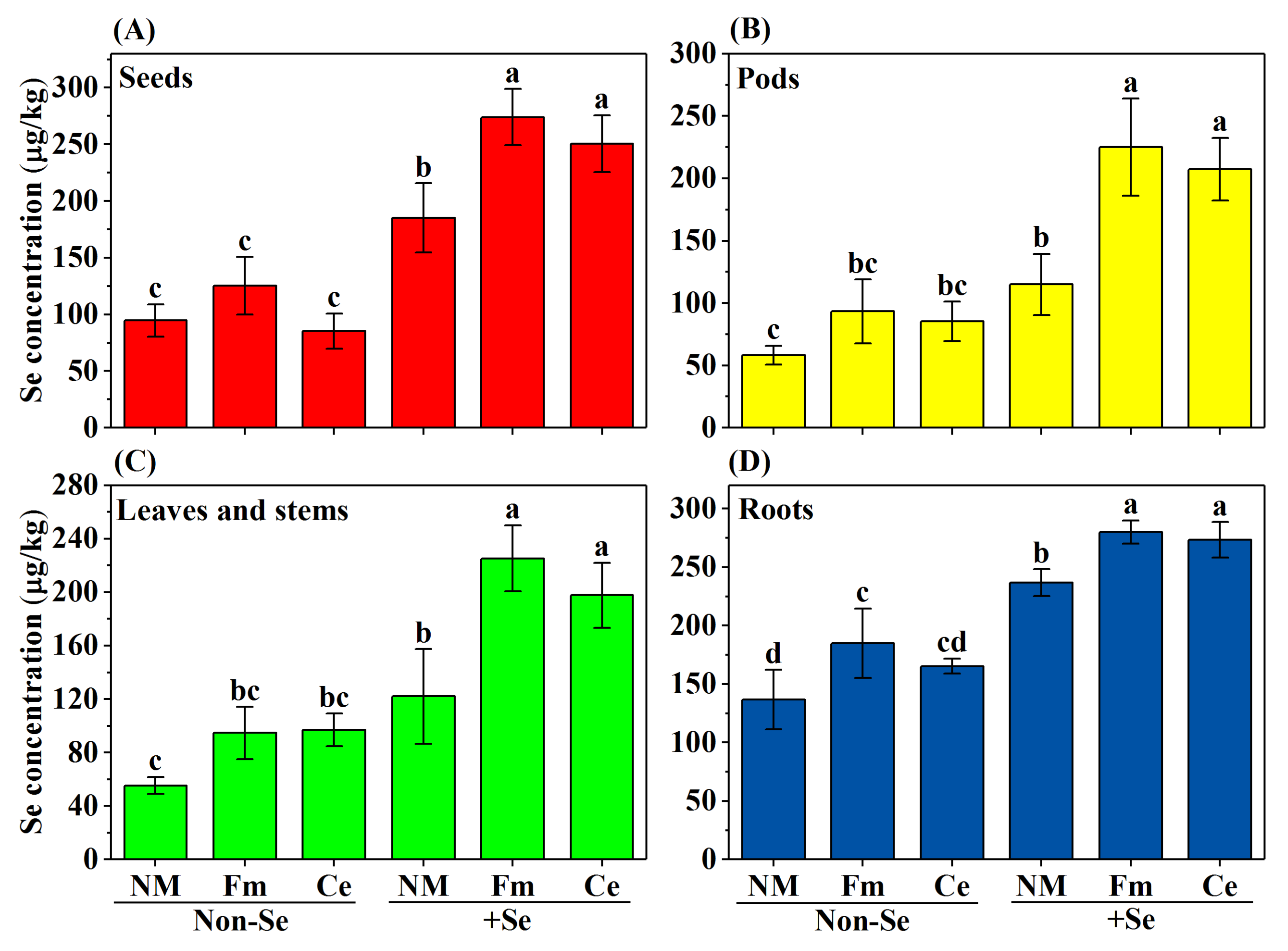
3.3. Distribution of Se Concentration in Soybean Organs
3.4. Speciation of Se in Soybean Seed
3.5. Bioaccumulation Factor and Translocation Coefficients of Se in Soybean
3.6. Se Biofortification Potentiality Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Al-Fartusie, F.S.; Mohssan, S.N. Essential trace elements and their vital roles in human body. Indian J. Adv. Chem. Sci. 2017, 5, 127–136. [Google Scholar] [CrossRef]
- Doroshow, J.H.; Juhasz, A. Modulation of selenium-dependent glutathione peroxidase activity enhances doxorubicin-induced apoptosis, tumour cell killing and hydroxyl radical production in human NCI/ADR-RES cancer cells despite high-level P-glycoprotein expression. Free Radic. Res. 2019, 53, 882–891. [Google Scholar] [CrossRef] [PubMed]
- Kieliszek, M.; Błażejak, S. Current knowledge on the importance of selenium in food for living organisms: A review. Molecules 2016, 21, 609. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.A.; Zhu, W.Y.; Wang, W.Y.; Li, R.B.; Hou, S.F.; Wang, D.C.; Yang, L.S. Selenium in soil and endemic diseases in China. Sci. Total Environ. 2002, 284, 227–235. [Google Scholar] [CrossRef]
- Li, N.; Gao, Z.D.; Luo, D.G.; Tang, X.; Chen, D.F.; Hu, Y.H. Selenium level in the environment and the population of Zhoukoudian area, Beijing, China. Sci. Total Environ. 2007, 381, 105–111. [Google Scholar] [CrossRef]
- Han, J.; Liang, H.; Yi, J.H.; Tan, W.H.; He, S.L.; Wu, X.F.; Shi, X.W.; Ma, J.; Guo, X. Selenium deficiency induced damages and altered expressions of metalloproteinases and their inhibitors (MMP1/3, TIMP1/3) in the kidneys of growing rats. J. Trace Elem. Med. Biol. 2016, 34, 1–9. [Google Scholar] [CrossRef]
- Fang, H.; Guo, X.; Farooq, U.; Xia, C.; Dong, R. Development and validation of a quality of life instrument for Kashin-Beck disease: An endemic osteoarthritis in China. Osteoarthr. Cartil. 2012, 20, 630–637. [Google Scholar] [CrossRef]
- Navarro-Alarcon, M.; Cabrera-Vique, C. Selenium in food and the human body: A review. Sci. Total Environ. 2008, 400, 115–141. [Google Scholar] [CrossRef]
- Matos-Reyes, M.N.; Cervera, M.L.; Campos, R.C.; De la Guardia, M. Total content of As, Sb, Se, Te and Bi in Spanish vegetables, cereals and pulses and estimation of the contribution of these foods to the Mediterranean daily intake of trace elements. Food Chem. 2010, 122, 188–194. [Google Scholar] [CrossRef]
- Sors, T.G.; Ellis, D.R.; Salt, D.E. Selenium uptake, translocation, assimilation and metabolic fate in plants. Photosynth. Res. 2005, 86, 373–389. [Google Scholar] [CrossRef]
- Wan, J.; Zhang, M.; Adhikari, B. Advances in selenium-enriched foods: From the farm to the fork. Trends Food Sci. Technol. 2018, 76, 1–5. [Google Scholar] [CrossRef]
- Ros, G.H.; Van Rotterdam, A.M.D.; Bussink, D.W.; Bindraban, P.S. Selenium fertilization strategies for bio-fortification of food: An agro-ecosystem approach. Plant Soil 2016, 404, 99–112. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Shen, F.K.; Gu, M.H.; Liu, Y.X.; Pan, L.S.; Shohag, M.J.I.; Li, T.T.; Wei, Y.Y. Evaluation of selenium bioavailability to Brassica juncea in representative Chinese soils based on diffusive gradients in thin-films (DGT) and chemical extraction methods. Int. J. Phytoremediat. 2020, 22, 952–962. [Google Scholar] [CrossRef]
- Alfthan, G.; Eurola, M.; Ekholm, P.; Venäläinen, E.R.; Root, T.; Korkalainen, K.; Hartikainen, H.; Salminen, P.; Hietaniemi, V.; Aspila, P.; et al. Effects of nationwide addition of selenium to fertilizers on foods, and animal and human health in Finland: From deficiency to optimal selenium status of the population. J. Trace Elem. Med. Biol. 2015, 31, 142–147. [Google Scholar] [CrossRef]
- Meetu, G.; Shikha, G. An overview of Selenium uptake, metabolism, and toxicity in plants. Front. Plant Sci. 2017, 7, 2074. [Google Scholar] [CrossRef]
- Yasin, M.; Mehdawi, A.F.E.; Jahn, C.E.; Anwar, A.; Turner, M.F.S.; Faisal, M.; Pilon-Smits, E.A.H. Seleniferous soils as a source for production of selenium-enriched foods and potential of bacteria to enhance plant selenium uptake. Plant Soil 2015, 386, 385–394. [Google Scholar] [CrossRef]
- Smith, F.A.; Smith, S.E. What is the significance of the arbuscular mycorrhizal colonisation of many economically important crop plants? Plant Soil 2011, 348, 63–79. [Google Scholar] [CrossRef]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef]
- Miransari, M. Contribution of arbuscular mycorrhizal symbiosis to plant growth under different types of soil stress. Plant Biol. 2010, 12, 563–569. [Google Scholar] [CrossRef]
- Wang, F.Y. Occurrence of arbuscular mycorrhizal fungi in mining-impacted sites and their contribution to ecological restoration: Mechanisms and applications. Crit. Rev. Environ. Sci. Technol. 2017, 47, 1901–1957. [Google Scholar] [CrossRef]
- Luo, W.Q.; Li, J.; Ma, X.N.; Niu, H.; Hou, S.W.; Wu, F.Y. Effect of arbuscular mycorrhizal fungi on uptake of selenate, selenite, and selenomethionine by roots of winter wheat. Plant Soil 2019, 438, 71–83. [Google Scholar] [CrossRef]
- Li, J.; Liu, R.F.; Wu, B.Y.; Zhang, C.Y.; Wang, J.F.; Lyu, L.H.; Tong, X.G.; Wu, F.Y. Influence of arbuscular mycorrhizal fungi on selenium uptake by winter wheat depends on the level of selenate spiked in soil. Chemosphere 2021, 2021, 132813. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, Z.Y.; Gu, M.H.; Li, H.; Shohag, M.J.I.; Shen, F.K.; Wang, X.L.; Wei, Y.Y. Combined use of arbuscular mycorrhizal fungus and selenium fertilizer shapes microbial community structure and enhances organic selenium accumulation in rice grain. Sci. Total Environ. 2020, 748, 141166. [Google Scholar] [CrossRef] [PubMed]
- Golubkina, N.; Amagova, Z.; Matsadze, V.; Zamana, S.; Tallarita, A.; Caruso, G. Effects of arbuscular mycorrhizal fungi on yield, biochemical characteristics, and elemental composition of garlic and onion under Selenium supply. Plants 2020, 9, 84. [Google Scholar] [CrossRef]
- Li, J.; Awasthi, M.K.; Xing, W.J.; Liu, R.F.; Bao, H.Y.; Wang, X.; Wang, J.F.; Wu, F.Y. Arbuscular mycorrhizal fungi increase the bioavailability and wheat (Triticum aestivum L.) uptake of selenium in soil. Ind. Crop Prod. 2020, 150, 112383. [Google Scholar] [CrossRef]
- Whanger, D.P. Selenocompounds in plants and animals and their biological significance. J. Am. Coll. Nutr. 2002, 21, 223–232. [Google Scholar] [CrossRef]
- Dai, Z.; Imtiaz, M.; Rizwan, M.; Yuan, Y.; Huang, H.L.; Tu, S.X. Dynamics of Selenium uptake, speciation, and antioxidant response in rice at different panicle initiation stages. Sci. Total Environ. 2019, 691, 827–834. [Google Scholar] [CrossRef]
- Rayman, M.P.; Infante, H.G.; Sargent, M. Food-chain selenium and human health: Spotlight on speciation. Br. J. Nutr. 2008, 100, 238–253. [Google Scholar] [CrossRef]
- Li, H.; Ye, Z.H.; Chan, W.F.; Chen, X.W.; Wu, F.Y.; Wu, S.C.; Wong, M.H. Can arbuscular mycorrhizal fungi improve grain yield, As uptake and tolerance of rice grown under aerobic conditions? Environ. Pollut. 2011, 159, 2537–2545. [Google Scholar] [CrossRef]
- Wang, M.K.; Cui, Z.W.; Xue, M.Y.; Peng, Q.; Zhou, F.; Wang, D.; Dinh, Q.T.; Liu, Y.X.; Liang, D.L. Assessing the uptake of selenium from naturally enriched soils by maize (Zea mays L.) using diffusive gradients in thin-films technique (DGT) and traditional extractions. Sci. Total Environ. 2019, 689, 1–9. [Google Scholar] [CrossRef]
- Sha, Z.M.; Chu, Q.N.; Zhao, Z.; Yue, Y.B.; Lu, L.F.; Yuan, J.; Cao, L.K. Variations in nutrient and trace element composition of rice in an organic rice-frog coculture system. Sci. Rep. 2017, 7, 15706. [Google Scholar] [CrossRef]
- Renkema, H.; Koopmans, A.; Kersbergen, L.; Kikkert, J.; Hale, B.; Berkelaar, E. The effect of transpiration on selenium uptake and mobility in durum wheat and spring canola. Plant Soil 2012, 354, 239–250. [Google Scholar] [CrossRef]
- Wu, Z.L.; Bañuelos, G.S.; Lin, Z.Q.; Liu, Y.; Yuan, L.X.; Yin, X.B.; Li, M. Biofortification and phytoremediation of selenium in China. Front. Plant Sci. 2015, 6, 136. [Google Scholar] [CrossRef]
- Ullah, H.; Liu, G.J.; Yousaf, B.; Ali, M.U.; Abbas, Q.; Munir, M.A.M.; Mian, M.M. Developmental selenium exposure and health risk in daily foodstuffs: A systematic review and meta-analysis. Ecotox. Environ. Safe 2018, 149, 291–306. [Google Scholar] [CrossRef]
- Patharajan, S.; Raaman, N. Influence of arbuscular mycorrhizal fungi on growth and selenium uptake by garlic plants. Arch Phytopathol. Plant Prot. 2012, 45, 138–151. [Google Scholar] [CrossRef]
- Nogueira, M.A.; Nehls, U.; Hampp, R.; Poralla, K.; Cardoso, E.J.B.N. Mycorrhiza and soil bacteria influence extractable iron and manganese in soil and uptake by soybean. Plant Soil. 2007, 298, 273–284. [Google Scholar] [CrossRef]
- Dehghanian, H.; Halajnia, A.; Lakzian, A.; Astaraei, A.R. The effect of earthworm and arbuscular mycorrhizal fungi on availability and chemical distribution of Zn, Fe and Mn in a calcareous soil. Appl. Soil Ecol. 2018, 130, 98–103. [Google Scholar] [CrossRef]
- Adeyemi, N.O.; Atayese, M.O.; Olubode, A.A.; Akan, M.E. Effect of commercial arbuscular mycorrhizal fungi inoculant on growth and yield of soybean under controlled and natural field conditions. J. Plant Nutr. 2020, 43, 487–499. [Google Scholar] [CrossRef]
- Alori, E.T.; Dare, M.O.; Babalola, O.O. Microbial Inoculants for Soil Quality and Plant Health; Lichtfouse, E., Ed.; Sustainable agriculture reviews; Springer International Publishing: Cham, Switzerland, 2017; pp. 281–307. [Google Scholar]
- Li, Z.; Liang, D.L.; Peng, Q.; Cui, Z.W.; Huang, J.; Lin, Z.Q. Interaction between selenium and soil organic matter and its impact on soil selenium bioavailability: A review. Geoderma 2017, 295, 69–79. [Google Scholar] [CrossRef]
- Peng, Q.; Wang, M.K.; Cui, Z.W.; Huang, J.; Chen, C.E.; Guo, L.; Liang, D.L. Assessment of bioavailability of selenium in different plant-soil systems by diffusive gradients in thin-films (DGT). Environ. Pollut. 2017, 225, 637–643. [Google Scholar] [CrossRef]
- Peng, Q.; Li, J.; Wang, D.; Wei, T.J.; Chen, C.E.L.; Liang, D.L. Effects of ageing on bioavailability of selenium in soils assessed by diffusive gradients in thin-films and sequential extraction. Plant Soil 2019, 436, 159–171. [Google Scholar] [CrossRef]
- Yu, Y.; Luo, L.; Yang, K.; Zhang, S.Z. Influence of mycorrhizal inoculation on the accumulation and speciation of selenium in maize growing in selenite and selenate spiked soils. Pedobiologia 2011, 54, 267–272. [Google Scholar] [CrossRef]
- Bamberg, S.M.; Ramos, S.J.; Aurelio, M.; Carneiro, M.A.C.; Siqueira, J.O. Effects of selenium (Se) application and arbuscular mycorrhizal (AMF) inoculation on soybean (Glycine max) and forage grass (Urochloa decumbens) development in oxisol. Aust. J. Crop Sci. 2019, 13, 1835–2707. [Google Scholar] [CrossRef]
- Li, X.; Wu, Y.S.; Li, B.Q.; Yang, Y.H.; Yang, Y.P. Selenium accumulation characteristics and biofortification potentiality in Turnip (Brassica rapa var. rapa) supplied with selenite or selenate. Front. Plant Sci. 2017, 8, 2207. [Google Scholar] [CrossRef]
- Michel-López, C.Y.; Espadas y Gil, F.; Fuentes Ortíz, G.; Santamaría, J.M.; González-Mendoza, D.; Ceceña-Duran, C.; Grimaldo Juarez, O. Bioaccumulation and effect of cadmium in the photosynthetic apparatus of Prosopis juliflora. Chem. Speciat. Bioavail. 2016, 28, 1–6. [Google Scholar] [CrossRef]
- Ximénez-Embún, P.; Alonso, I.; Madrid-Albarrán, Y.; Cámara, C. Establishment of selenium uptake and species distribution in lupine, Indian mustard, and sunflower plants. J. Agric. Food Chem. 2004, 52, 832–838. [Google Scholar] [CrossRef]
- Galinha, C.; Sánchez-Martínez, M.; Pacheco, A.M.G.; Freitas, M.D.C.; Coutinho, J.; Maçãs, B.; Almeida, A.S.; Pérez-Corona, M.T.; Madrid, Y.; Wolterbeek, H.T. Characterization of selenium-enriched wheat by agronomic biofortification. J. Food Sci. Technol. Mysore. 2015, 52, 4236–4245. [Google Scholar] [CrossRef]
- Li, H.F.; Lombi, E.; Stroud, J.L.; McGrath, S.P.; Zhao, F.J. Selenium speciation in soil and rice: Influence of water management and Se fertilization. J. Agric. Food Chem. 2010, 58, 11837–11843. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, S.Z.; Wen, B.; Huang, H.L.; Luo, L. Accumulation and speciation of selenium in plants as affected by arbuscular mycorrhizal fungus glomus mosseae. Biol. Trace Elem. Res. 2011, 143, 1789–1798. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Selenium Fertilization | Inoculation Treatment | Colonization Rate (%) | Bean (g/Pot) b | Leaf and Stem (g/Pot) | Root (g/Pot) |
|---|---|---|---|---|---|
| Non-Se | NM | - | 2.55 ± 1.02b | 3.26 ± 1.09bc | 0.26 ± 0.01b |
| Fm | 30.13 ± 0.81b | 5.99 ± 0.25a | 4.72 ± 0.36ab | 0.54 ± 0.21a | |
| Ce | 30.77 ± 1.05b | 4.87 ± 0.63a | 4.75 ± 0.96ab | 0.57 ± 0.16a | |
| +Se | NM | - | 2.08 ± 0.23b | 3.01 ± 0.13c | 0.21 ± 0.03b |
| Fm | 34.13 ± 0.26a | 5.15 ± 0.77a | 5.65 ± 1.01a | 0.51 ± 0.12a | |
| Ce | 31.69 ± 1.06b | 5.13 ± 0.56a | 5.62 ± 0.91a | 0.42 ± 0.07ab | |
| Source of variation | Se | *** | NS | NS | NS |
| AMF | *** | *** | *** | ** | |
| Se×AMF | *** | NS | NS | NS |
| Source of Variation | Se Concentration in Plant (mg/kg) | |||
|---|---|---|---|---|
| Bean | Pod | Stem and Leaf | Root | |
| Se | *** | *** | *** | *** |
| AMF | ** | *** | *** | ** |
| Se × AMF | ** | ** | NS | NS |
| Selenium Fertilization | Inoculation Treatment | BAF roots | TCroots-straws b | TCstraws to pods | TC pods to seeds |
|---|---|---|---|---|---|
| Non-Se | NM | 0.27 ± 0.05d | 0.40 ± 0.05c | 0.43 ± 0.05b | 0.69 ± 0.10c |
| Fm | 0.37 ± 0.06c | 0.51 ± 0.11c | 0.50 ± 0.14b | 0.68 ± 0.14bc | |
| Ce | 0.33 ± 0.01cd | 0.59 ± 0.07bc | 0.52 ± 0.09b | 0.52 ± 0.09bc | |
| +Se | NM | 0.47 ± 0.02b | 0.52 ± 0.15c | 0.49 ± 0.10b | 0.78 ± 0.13ab |
| Fm | 0.56 ± 0.02a | 0.80 ± 0.09a | 0.80 ± 0.14a | 0.98 ± 0.09a | |
| Ce | 0.55 ± 0.03a | 0.72 ± 0.09ab | 0.76 ± 0.09a | 0.92 ± 0.09a | |
| Source of variation | Se | *** | ** | ** | *** |
| AMF | ** | ** | * | * | |
| Se×AMF | NS | NS | NS | NS |
| Selenium Fertilization | Inoculation Treatment | Daily Intake of Se (μg) |
|---|---|---|
| Non-Se | NM | 32.66 |
| Fm | 43.24 | |
| Ce | 29.44 | |
| +Se | NM | 65.55 |
| Fm | 93.15 | |
| Ce | 86.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Li, B.; Liu, Y.; He, L.; Pang, T.; Chen, Z.; Shohag, M.J.I.; Miao, X.; Li, X.; Gu, M.; et al. Arbuscular Mycorrhizal Fungal Inoculation Increases Organic Selenium Accumulation in Soybean (Glycine max (Linn.) Merr.) Growing in Selenite-Spiked Soils. Toxics 2022, 10, 565. https://doi.org/10.3390/toxics10100565
Zhang Z, Li B, Liu Y, He L, Pang T, Chen Z, Shohag MJI, Miao X, Li X, Gu M, et al. Arbuscular Mycorrhizal Fungal Inoculation Increases Organic Selenium Accumulation in Soybean (Glycine max (Linn.) Merr.) Growing in Selenite-Spiked Soils. Toxics. 2022; 10(10):565. https://doi.org/10.3390/toxics10100565
Chicago/Turabian StyleZhang, Zengyu, Bei Li, Yongxian Liu, Lixin He, Ting Pang, Zongdao Chen, Md. Jahidul Islam Shohag, Xiuyan Miao, Xi Li, Minghua Gu, and et al. 2022. "Arbuscular Mycorrhizal Fungal Inoculation Increases Organic Selenium Accumulation in Soybean (Glycine max (Linn.) Merr.) Growing in Selenite-Spiked Soils" Toxics 10, no. 10: 565. https://doi.org/10.3390/toxics10100565