Inhibitory Activities of Polyphenolic Extracts of Bangladeshi Vegetables against α-Amylase, α-Glucosidase, Pancreatic Lipase, Renin, and Angiotensin-Converting Enzyme

 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction of Polyphenolic Compounds
2.3. UHPLC MS/MS Analysis of Polyphenolic Compounds
2.4. Inhibition of α-Amylase Activity
2.5. Inhibition of α-Glucosidase Activity
2.6. Inhibition of Lipase Activity
2.7. ACE Inhibition Assay
2.8. Renin Inhibition Assay
2.9. Statistical Analysis
3. Results
3.1. UHPLC MS/MS Analysis
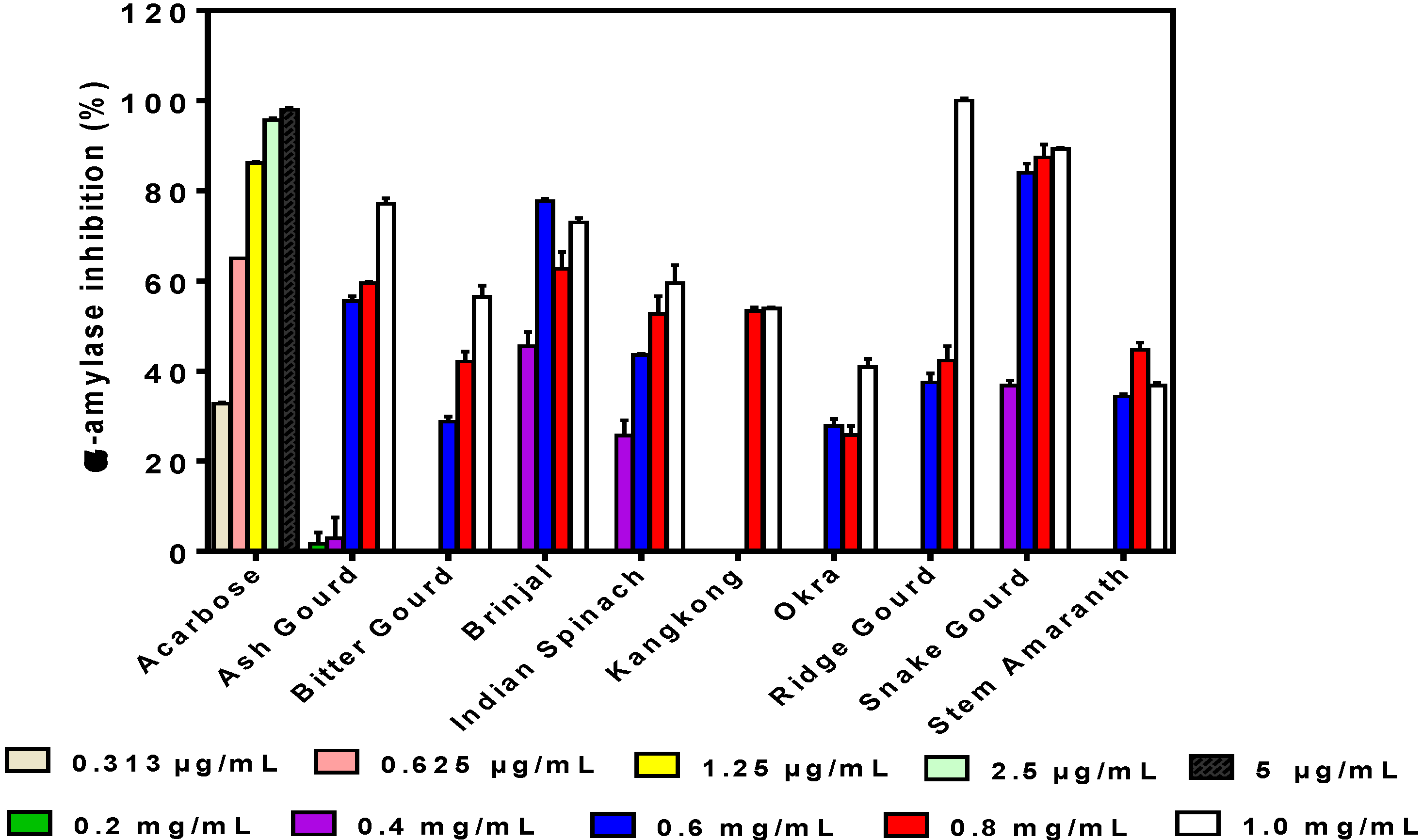
3.2. α-Amylase Inhibition
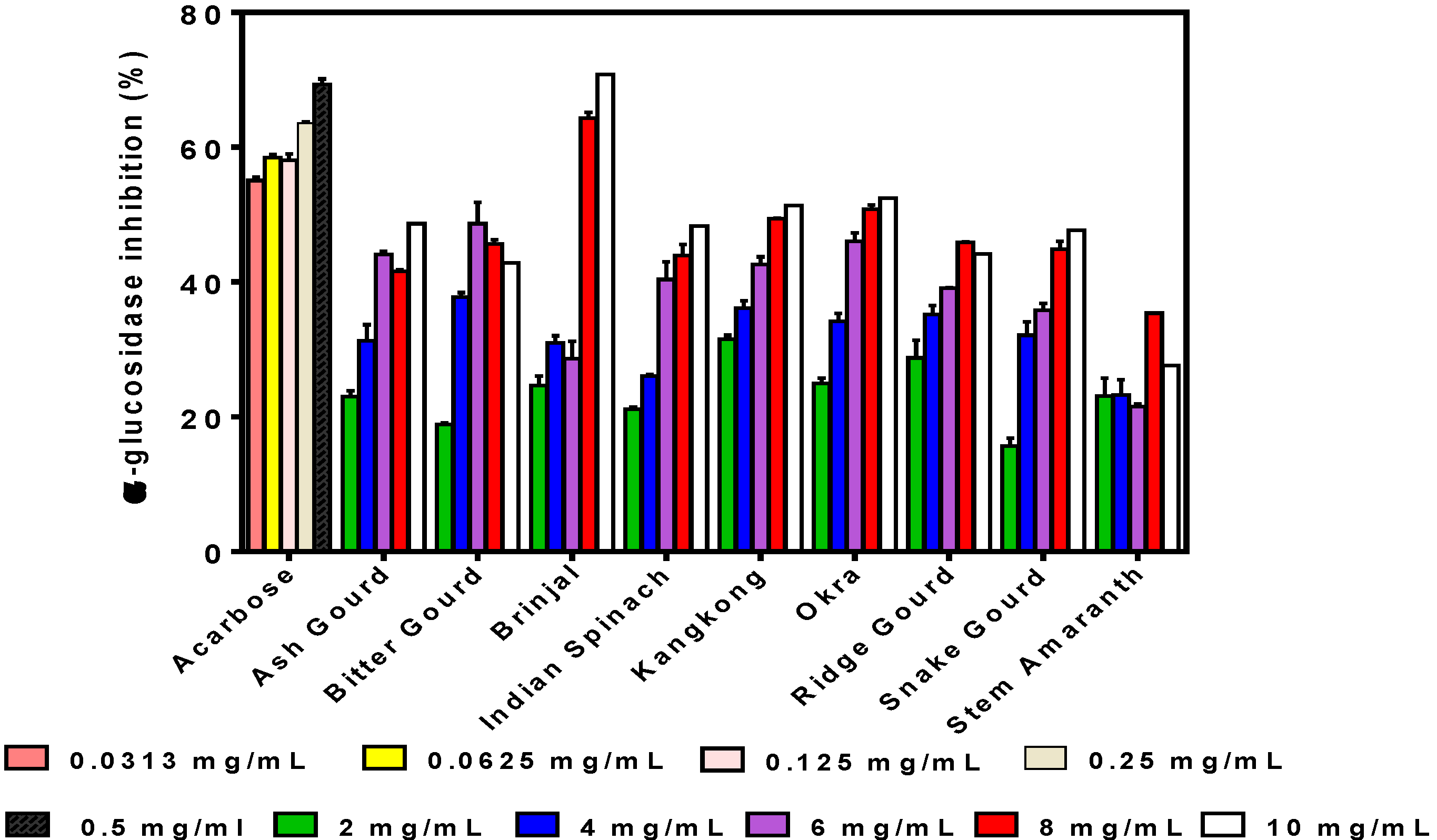
3.3. α-Glucosidase Inhibition
3.4. Pancreatic Lipase (PL)-Inhibitory Activity
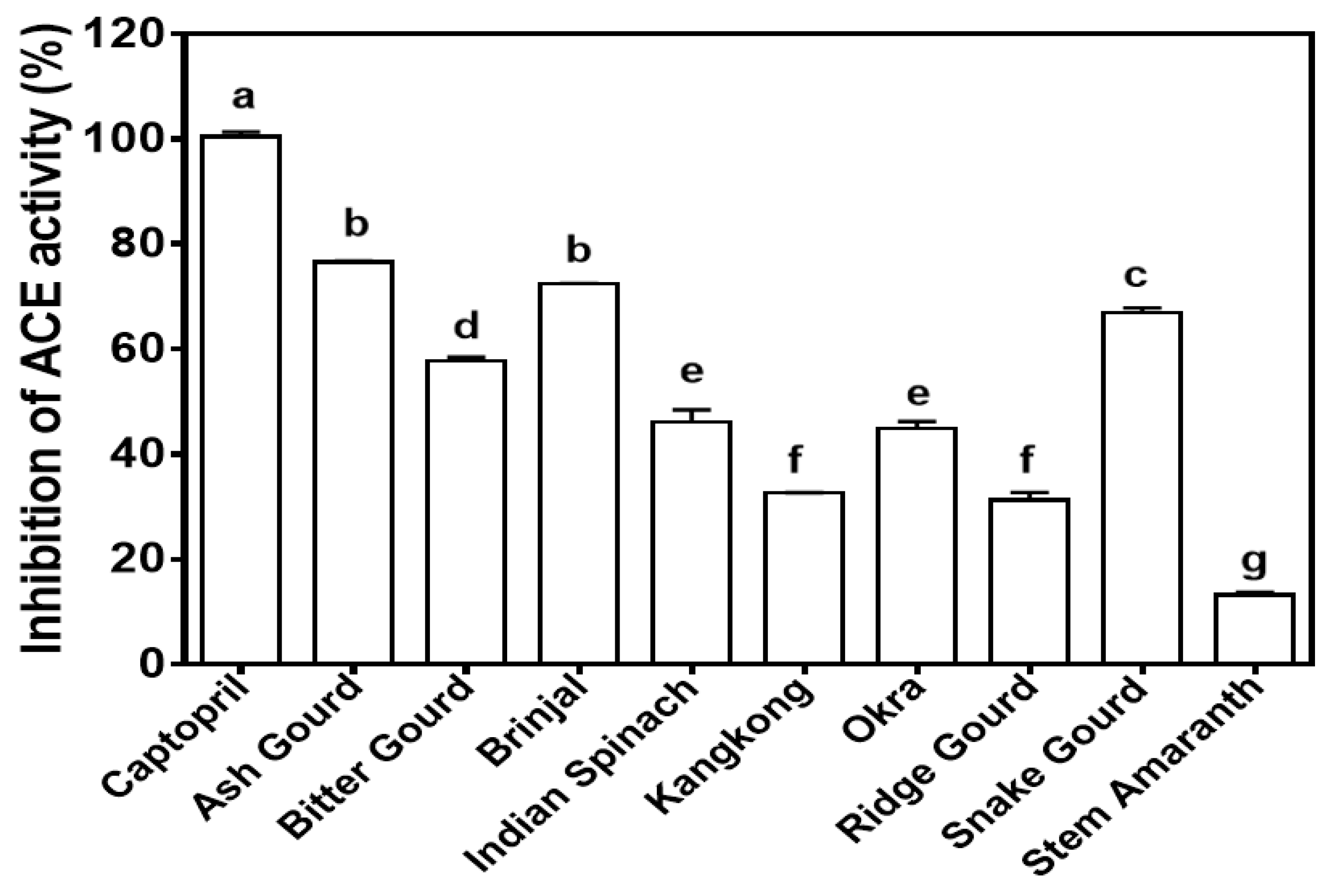
3.5. Angiotensin-Converting Enzyme (ACE)-Inhibitory Activity
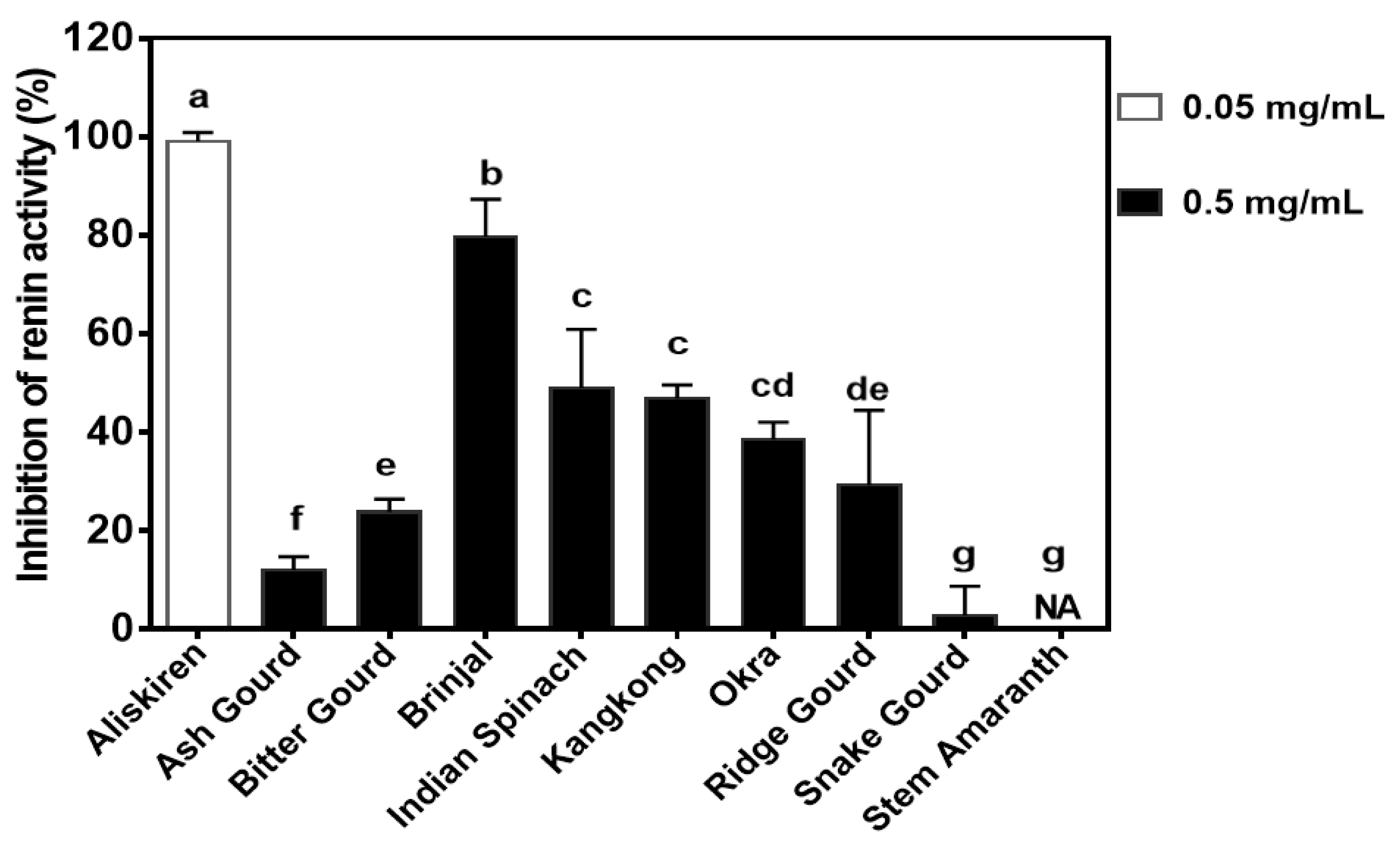
3.6. Renin-Inhibitory Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Miglani, N.; Bains, K. Interplay between proteins and metabolic syndrome—A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 2483–2496. [Google Scholar] [CrossRef]
- Handy, D.E.; Castro, R.; Loscalzo, J. Epigenetic modifications: Basic mechanisms and role in cardiovascular disease. Circulation 2011, 123, 2145–2156. [Google Scholar] [CrossRef]
- Tucci, S.A.; Boyland, E.J.; Halford, J.C. The role of lipid and carbohydrate digestive enzyme inhibitors in the management of obesity: A review of current and emerging therapeutic agents. Diabetes Metab. Syndr. Obes. 2010, 3, 125–143. [Google Scholar] [CrossRef]
- Poovitha, S.; Parani, M. In vitro and in vivo α-amylase and α-glucosidase inhibiting activities of the protein extracts from two varieties of bitter gourd (Momordica charantia L.). BMC Complement. Altern. Med. 2016, 16, 185. [Google Scholar] [CrossRef] [PubMed]
- Wongsa, P.; Chaiwarit, J.; Zamaludien, A. In vitro screening of phenolic compounds, potential inhibition against α-amylase and α-glucosidase of culinary herbs in Thailand. Food Chem. 2012, 131, 964–971. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, S.; Sweet, R.M.; Suh, S.W. Crystallization and a preliminary X-ray crystallographic study of α-amylase from Bacillus licheniformis. Arch. Biochem. Biophys. 1991, 291, 255–257. [Google Scholar] [CrossRef]
- Patil, P.; Mandal, S.; Tomar, S.K.; Anand, S. Food protein-derived bioactive peptides in management of type 2 diabetes. Eur. J. Nutr. 2015, 54, 863–880. [Google Scholar] [CrossRef] [PubMed]
- Thanakosai, W.; Phuwapraisirisan, P. First identification of α-glucosidase inhibitors from okra (Abelmoschus esculentus) seeds. Nat. Prod. Commun. 2013, 8, 1085–1088. [Google Scholar] [CrossRef] [PubMed]
- De La Garza, A.L.; Milagro, F.I.; Boque, N.; Campión, J.; Martínez, J.A. Natural inhibitors of pancreatic lipase as new players in obesity treatment. Planta Med. 2011, 77, 773–785. [Google Scholar] [CrossRef]
- Ombra, M.N.; d’Acierno, A.; Nazzaro, F.; Spigno, P.; Riccardi, R.; Zaccardelli, M.; Fratianni, F. Alpha-amylase, α-glucosidase and lipase inhibiting activities of polyphenol-rich extracts from six common bean cultivars of Southern Italy, before and after cooking. Int. J. Food Sci. Nutr. 2018, 69, 824–834. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Shimoda, H.; Nishida, N.; Takada, M.; Matsuda, H. Salacia reticulata and its polyphenolic constituents with lipase inhibitory and lipolytic activities have mild antiobesity effects in rats. J. Nutr. 2002, 132, 1819–1824. [Google Scholar] [CrossRef] [PubMed]
- Ballinger, A.; Peikin, S.R. Orlistat: Its current status as an anti-obesity drug. Eur. J. Pharmacol. 2002, 440, 109–117. [Google Scholar] [CrossRef]
- Drew, B.S.; Dixon, A.F.; Dixon, J.B. Obesity management: Update on orlistat. Vasc. Health Risk Manag. 2007, 3, 817–821. [Google Scholar] [PubMed]
- Chiou, S.Y.; Sung, J.M.; Huang, P.W.; Lin, S.D. Antioxidant, antidiabetic, and antihypertensive properties of Echinacea purpurea flower extract and caffeic acid derivatives using in vitro models. J. Med. Food 2017, 20, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Aluko, R.E. Food protein-derived renin-inhibitory peptides: In vitro and in vivo properties. J. Food Biochem. 2019, 43, e12648. [Google Scholar] [CrossRef]
- Da-Costa-Rocha, I.; Bonnlaender, B.; Sievers, H.; Pischel, I.; Heinrich, M. Hibiscus sabdariffa L.—A phytochemical and pharmacological review. Food Chem. 2014, 165, 424–443. [Google Scholar] [CrossRef]
- Ademiluyi, A.O.; Oboh, G.; Ogunsuyi, O.B.; Oloruntoba, F.M. A comparative study on antihypertensive and antioxidant properties of phenolic extracts from fruit and leaf of some guava (Psidium guajava L.) varieties. Comp. Clin. Pathol. 2016, 25, 363–374. [Google Scholar] [CrossRef]
- Chiang, Y.C.; Chen, C.L.; Jeng, T.L.; Sung, J.M. In vitro inhibitory effects of cranberry bean (Phaseolus vulgaris L.) extracts on aldose reductase, α-glucosidase and α-amylase. Int. J. Food Sci. Technol. 2014, 49, 1470–1479. [Google Scholar] [CrossRef]
- Chiou, S.Y.; Lai, J.Y.; Liao, J.A.; Sung, J.M.; Lin, S.D. In vitro inhibition of lipase, α-amylase, α-glucosidase, and angiotensin-converting enzyme by defatted rice bran extracts of red-pericarp rice mutant. Cereal Chem. 2018, 95, 167–176. [Google Scholar] [CrossRef]
- Gondoin, A.; Grussu, D.; Stewart, D.; McDougall, G.J. White and green tea polyphenols inhibit pancreatic lipase in vitro. Food Res. Int. 2010, 43, 1537–1544. [Google Scholar] [CrossRef]
- Alashi, A.M.; Taiwo, K.A.; Oyedele, D.; Adebooye, O.C.; Aluko, R.E. Antihypertensive properties of aqueous extracts of vegetable leaf-fortified bread after oral administration to spontaneously hypertensive rats. Int. J. Food Sci. Technol. 2018, 53, 1705–1716. [Google Scholar] [CrossRef]
- Oluwagunwa, O.A.; Alashi, A.M.; Aluko, R.E. Solanum Macrocarpon leaf extracts reduced blood pressure and heart rate after oral administration to spontaneously hypertensive rats. Curr. Top. Nutraceut. Res. 2019, 17, 282–290. [Google Scholar] [CrossRef]
- Olarewaju, O.A.; Alashi, A.M.; Aluko, R.E. Antihypertensive effect of aqueous polyphenol extracts of Amaranthus viridis and Telfairia occidentalis leaves in spontaneously hypertensive rats. J. Food Bioact. 2018, 1, 166–173. [Google Scholar] [CrossRef][Green Version]
- Ranilla, L.G.; Kwon, Y.I.; Apostolidis, E.; Shetty, K. Phenolic compounds, antioxidant activity and in vitro inhibitory potential against key enzymes relevant for hyperglycemia and hypertension of commonly used medicinal plants, herbs and spices in Latin America. Bioresour. Technol. 2010, 101, 4676–4689. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhang, B.; Li, X.; Chen, P.X.; Zhang, H.; Liu, R.; Tsao, R. Bound phenolics of quinoa seeds released by acid, alkaline, and enzymatic treatments and their antioxidant and α-glucosidase and pancreatic lipase inhibitory effects. J. Agric. Food Chem. 2016, 64, 1712–1719. [Google Scholar] [CrossRef]
- Sonklin, C.; Alashi, M.A.; Laohakunjit, N.; Kerdchoechuen, O.; Aluko, R.E. Identification of antihypertensive peptides from mung bean protein hydrolysate and their effects in spontaneously hypertensive rats. J. Funct. Foods 2019, 64, 103635. [Google Scholar] [CrossRef]
- Verdu, C.F.; Gatto, J.; Freuze, I.; Richomme, P.; Laurens, F.; Guilet, D. Comparison of two methods, UHPLC-UV and UHPLC-MS/MS, for the quantification of polyphenols in cider apple juices. Molecules 2013, 18, 10213–10227. [Google Scholar] [CrossRef]
- Vijayaraj, P.; Nakagawa, H.; Yamaki, K. Cyanidin and cyanidin-3-glucoside derived from Vigna unguiculata act as noncompetitive inhibitors of pancreatic lipase. J. Food Biochem. 2019, 43, e12774. [Google Scholar] [CrossRef]
- Chizoba, E.F.G.; Suneetha, J.W.; Maheswari, K.; Sucharitha, D.S.; Prasad, T.N.V.K.V. In vitro carbohydrate digestibility of copper nanoparticulated bitter gourd extract. J. Nutr. Food Sci. 2016, 6, 1000482. [Google Scholar] [CrossRef]
- Pinto, M.D.S.; Ranilla, L.G.; Apostolidis, E.; Lajolo, F.M.; Genovese, M.I.; Shetty, K. Evaluation of antihyperglycemia and antihypertension potential of native Peruvian fruits using in vitro models. J. Med. Food 2009, 12, 278–291. [Google Scholar] [CrossRef]
- Cheplick, S.; Kwon, Y.I.; Bhowmik, P.; Shetty, K. Phenolic-linked variation in strawberry cultivars for potential dietary management of hyperglycemia and related complications of hypertension. Bioresour. Technol. 2010, 101, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Akinyemi, A.J.; Ademiluyi, A.O. Inhibition of α-amylase and α-glucosidase activities by ethanolic extract of Telfairia occidentalis (fluted pumpkin) leaf. Asian Pac. J. Trop. Biomed. 2012, 2, 733–738. [Google Scholar] [CrossRef]
- Awosika, T.O.; Aluko, R.E. Inhibition of the in vitro activities of α-amylase, α-glucosidase and pancreatic lipase by yellow field pea (Pisum sativum L.) protein hydrolysates. Int. J. Food Sci. Technol. 2019, 54, 2021–2034. [Google Scholar] [CrossRef]
- Nanok, K.; Sansenya, S. α-Glucosidase, α-amylase, and tyrosinase inhibitory potential of capsaicin and dihydrocapsaicin. J. Food Biochem. 2020, 44, e13099. [Google Scholar] [CrossRef]
- Nwanna, E.; Ibukun, E.O.; Oboh, G. Inhibitory effects of methanolic extracts of two eggplant species from South-western Nigeria on starch hydrolysing enzymes linked to type-2 diabetes. Afr. J. Pharm. Pharmacol. 2013, 7, 1575–1584. [Google Scholar] [CrossRef]
- Sulaiman, S.F.; Ooi, K.L.; Supriatno. Antioxidant and α-glucosidase inhibitory activities of cucurbit fruit vegetables and identification of active and major constituents from phenolic-rich extracts of Lagenaria siceraria and Sechium edule. J. Agric. Food Chem. 2013, 61, 10080–10090. [Google Scholar] [CrossRef]
- Patil, M.; Patil, R.; Bhadane, B.; Mohammad, S.; Maheshwari, V. Pancreatic lipase inhibitory activity of phenolic inhibitor from endophytic Diaporthe arengae. Biocat. Agric. Biotechnol. 2017, 10, 234–238. [Google Scholar] [CrossRef]
- Embleton, J.K.; Pouton, C.W. Structure and function of gastro-intestinal lipases. Adv. Drug Deliv. Rev. 1997, 25, 15–32. [Google Scholar] [CrossRef]
- Seyedan, A.; Alshawsh, M.A.; Alshagga, M.A.; Koosha, S.; Mohamed, Z. Medicinal plants and their inhibitory activities against pancreatic lipase: A review. Evid. Based Complement. Altern. Med. 2015, 2015, 973143. [Google Scholar] [CrossRef]
- Cooper, W.O.; Hernandez-Diaz, S.; Arbogast, P.G.; Dudley, J.A.; Dyer, S.; Gideon, P.S.; Ray, W.A. Major congenital malformations after first-trimester exposure to ACE inhibitors. N. Engl. J. Med. 2006, 354, 2443–2451. [Google Scholar] [CrossRef]
- Actis-Goretta, L.; Ottaviani, J.I.; Fraga, C.G. Inhibition of angiotensin converting enzyme activity by flavanol-rich foods. J. Agric. Food Chem. 2006, 54, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Afonso, J.; Passos, C.P.; Coimbra, M.A.; Silva, C.M.; Soares-da-Silva, P. Inhibitory effect of phenolic compounds from grape seeds (Vitis vinifera L.) on the activity of angiotensin I converting enzyme. LWT Food Sci. Technol. 2013, 54, 265–270. [Google Scholar] [CrossRef]
- Al Shukor, N.; Van Camp, J.; Gonzales, G.B.; Staljanssens, D.; Struijs, K.; Zotti, M.J.; Smagghe, G. Angiotensin-converting enzyme inhibitory effects by plant phenolic compounds: A study of structure activity relationships. J. Agric. Food Chem. 2013, 61, 11832–11839. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, L.; Castillo, J.; Quiñones, M.; Garcia-Vallvé, S.; Arola, L.; Pujadas, G.; Muguerza, B. Inhibition of angiotensin-converting enzyme activity by flavonoids: Structure-activity relationship studies. PLoS ONE 2012, 7, e49493. [Google Scholar] [CrossRef]
- Ojeda, D.; Jiménez-Ferrer, E.; Zamilpa, A.; Herrera-Arellano, A.; Tortoriello, J.; Alvarez, L. Inhibition of angiotensin convertin enzyme (ACE) activity by the anthocyanins delphinidin- and cyanidin-3-O-sambubiosides from Hibiscus sabdariffa. J. Ethnopharmacol. 2010, 127, 7–10. [Google Scholar] [CrossRef]
- Pechanova, O.; Barta, A.; Koneracka, M.; Zavisova, V.; Kubovcikova, M.; Klimentova, J.; Cebova, M. Protective effects of nanoparticle-loaded aliskiren on cardiovascular system in spontaneously hypertensive rats. Molecules 2019, 24, 15. [Google Scholar] [CrossRef]
- Wood, J.M.; Maibaum, J.; Rahuel, J.; Grutter, M.G.; Cohen, N.-C.; Rasetti, V.; Bedigian, M.P. Structure-based design of aliskiren, a novel orally effective renin inhibitor. Biochem. Biophys. Res. Commun. 2003, 308, 698–705. [Google Scholar] [CrossRef]
- Tavassoli, Z.; Taghdir, M.; Ranjbar, B. Renin inhibition by soyasaponin I: A potent native anti-hypertensive compound. J. Biomol. Struct. Dyn. 2018, 36, 166–176. [Google Scholar] [CrossRef]
- Hiwatashi, K.; Shirakawa, H.; Hori, K.; Yoshiki, Y.; Suzuki, N.; Hokari, M.; Komai, M.; Takahashi, S. Reduction of blood pressure by soybean saponins, renin inhibitors from soybean, in spontaneously hypertensive rats. Biosci. Biotechnol. Biochem. 2010, 74, 2310–2312. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Polyphenolic Compounds | Retention | m/z | MS/MS | Concentration | |
|---|---|---|---|---|---|---|
| Time (min) | (Da) | (Da) | (mg/g) | |||
| A 1 | B 2 | |||||
| Ash Gourd | Vitexin ramnoside | 3.2 | 579 | 433, 313, 283 | 1.23 | 0.77 |
| Brinjal | Chlorogenic acid | 2.4 | 355 | 163 | 5.63 | 4.07 |
| Indian spinach | Vitexin arabinoside | 3.0 | 465 | 433, 313, 283 | 8.96 | 3.40 |
| Vitexin | 3.2 | 433 | 313, 283 | 2.82 | 1.07 | |
| Kaempferol O-rutinoside | 3.4 | 595 | 449, 287 | 0.12 | 0.05 | |
| Kangkong | Dicaffeoyl quinic acid | 3.5 | 517 | 163 | 6.92 | 3.72 |
| Quercetin hexoside | 3.4 | 465 | 163 | 4.46 | 2.40 | |
| Chlorogenic, isochlorogenic acid | 2.5 | 355 | 163 | 10.12 | 5.43 | |
| Okra | Quercetin O-sophoroside | 3.0 | 627 | 301 | 3.33 | 2.28 |
| Quercetin O-hexoside | 3.4 | 465 | 301 | 2.63 | 1.80 | |
| Quercetin malonyl O-hexoside | 3.5 | 551 | 301 | 0.66 | 0.45 | |
| Snake gourd | Kaempferol O-sophoroside | 3.1 | 611 | 287 | 81.35 | 57.15 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sultana, R.; Alashi, A.M.; Islam, K.; Saifullah, M.; Haque, C.E.; Aluko, R.E. Inhibitory Activities of Polyphenolic Extracts of Bangladeshi Vegetables against α-Amylase, α-Glucosidase, Pancreatic Lipase, Renin, and Angiotensin-Converting Enzyme . Foods 2020, 9, 844. https://doi.org/10.3390/foods9070844
Sultana R, Alashi AM, Islam K, Saifullah M, Haque CE, Aluko RE. Inhibitory Activities of Polyphenolic Extracts of Bangladeshi Vegetables against α-Amylase, α-Glucosidase, Pancreatic Lipase, Renin, and Angiotensin-Converting Enzyme . Foods. 2020; 9(7):844. https://doi.org/10.3390/foods9070844
Chicago/Turabian StyleSultana, Razia, Adeola M. Alashi, Khaleda Islam, Md Saifullah, C. Emdad Haque, and Rotimi E. Aluko. 2020. "Inhibitory Activities of Polyphenolic Extracts of Bangladeshi Vegetables against α-Amylase, α-Glucosidase, Pancreatic Lipase, Renin, and Angiotensin-Converting Enzyme " Foods 9, no. 7: 844. https://doi.org/10.3390/foods9070844
APA StyleSultana, R., Alashi, A. M., Islam, K., Saifullah, M., Haque, C. E., & Aluko, R. E. (2020). Inhibitory Activities of Polyphenolic Extracts of Bangladeshi Vegetables against α-Amylase, α-Glucosidase, Pancreatic Lipase, Renin, and Angiotensin-Converting Enzyme . Foods, 9(7), 844. https://doi.org/10.3390/foods9070844