Food Authentication: Identification and Quantitation of Different Tuber Species via Capillary Gel Electrophoresis and Real-Time PCR

Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Material
2.2. DNA Isolation
2.3. Preparation of Spiked Sample Material
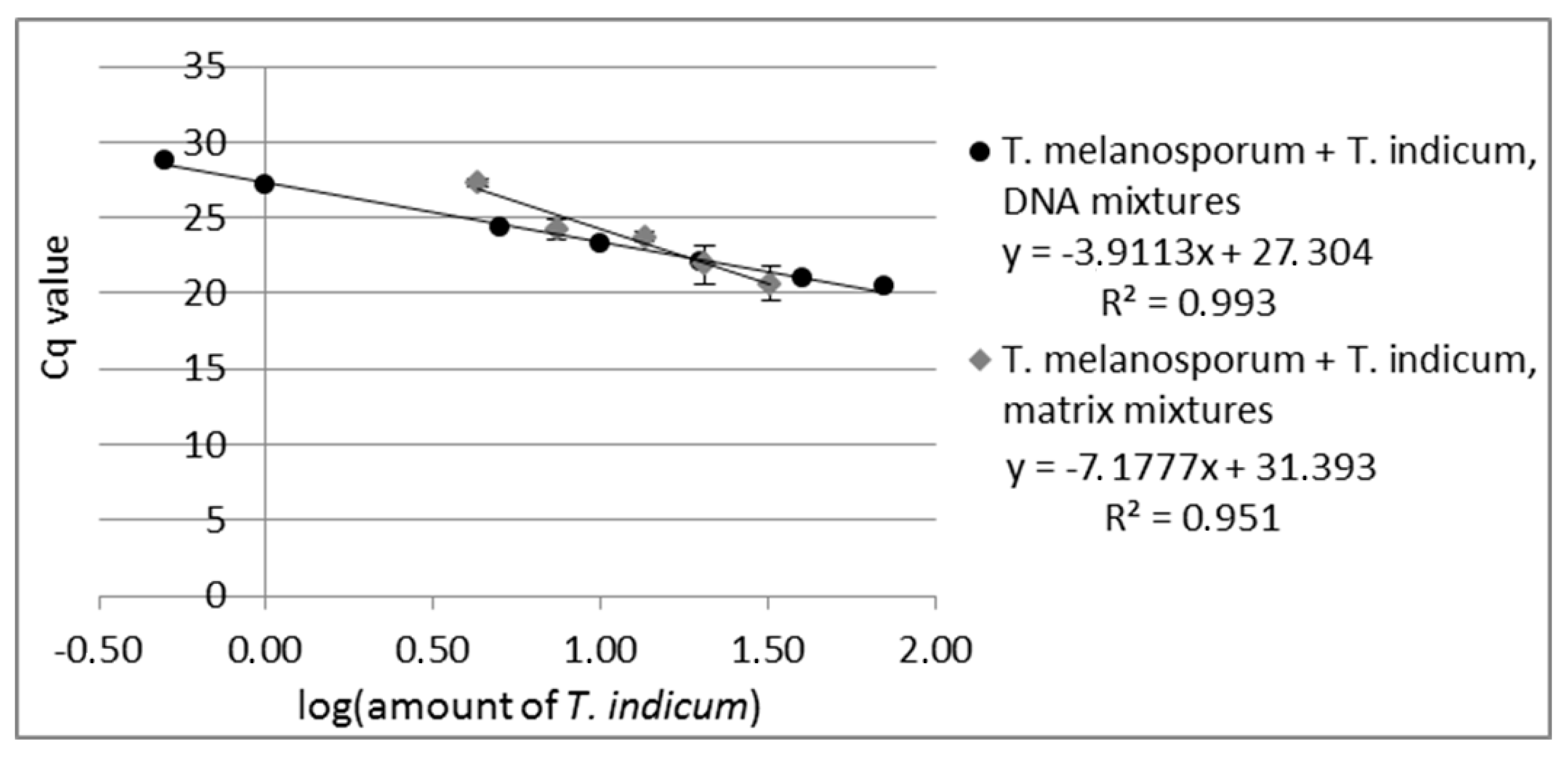
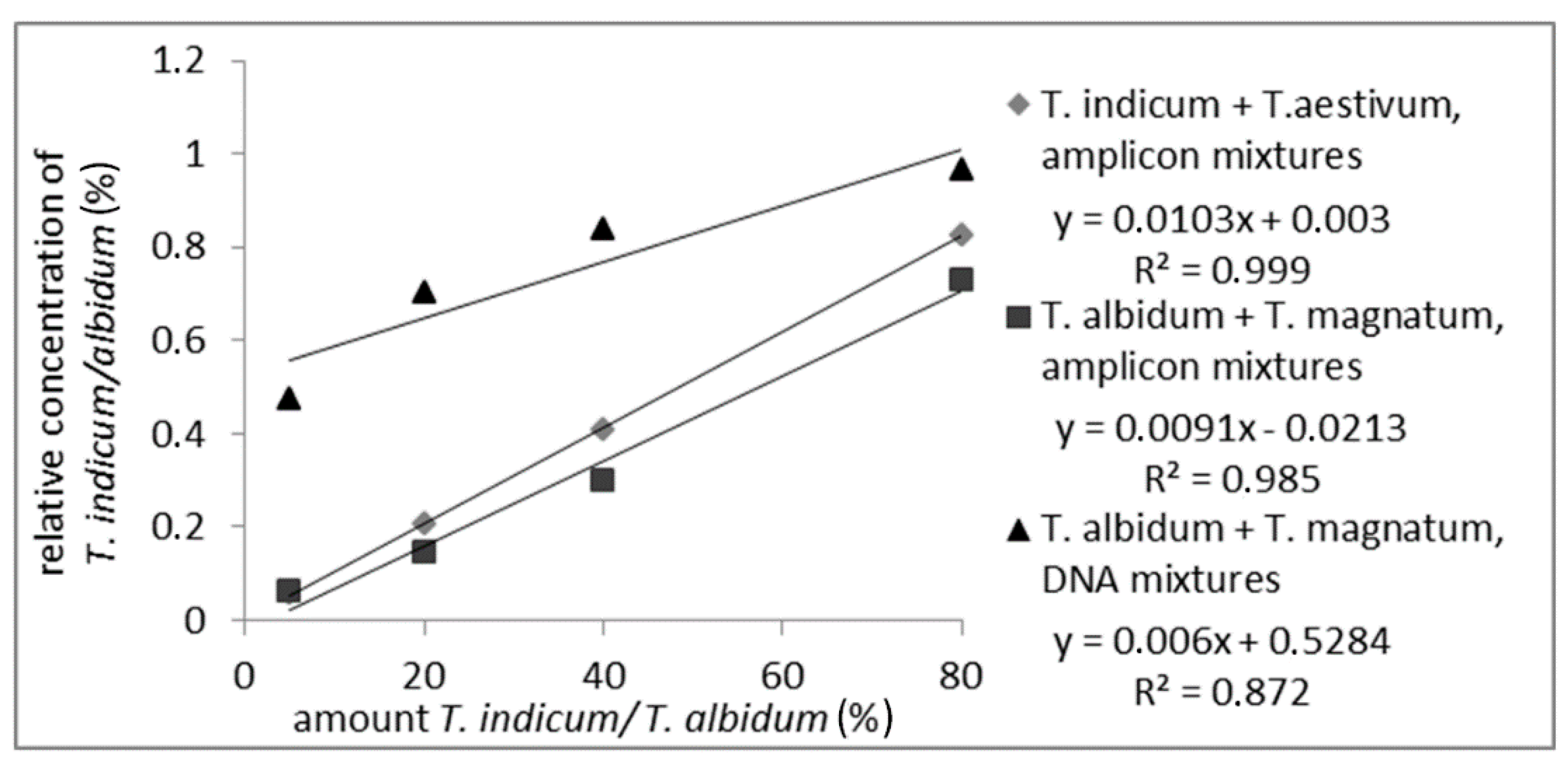
2.3.1. DNA Mixtures
2.3.2. Matrix Mixtures of Fruiting Bodies
2.4. Real-Time PCR
Practical Determination of LoD
2.5. Isolation of DNA Fragments from Agarose Gels
2.6. RFLP and CGE
3. Results and Discussion
3.1. Real-Time PCR
3.1.1. Primer Specificity
3.1.2. Quantitation
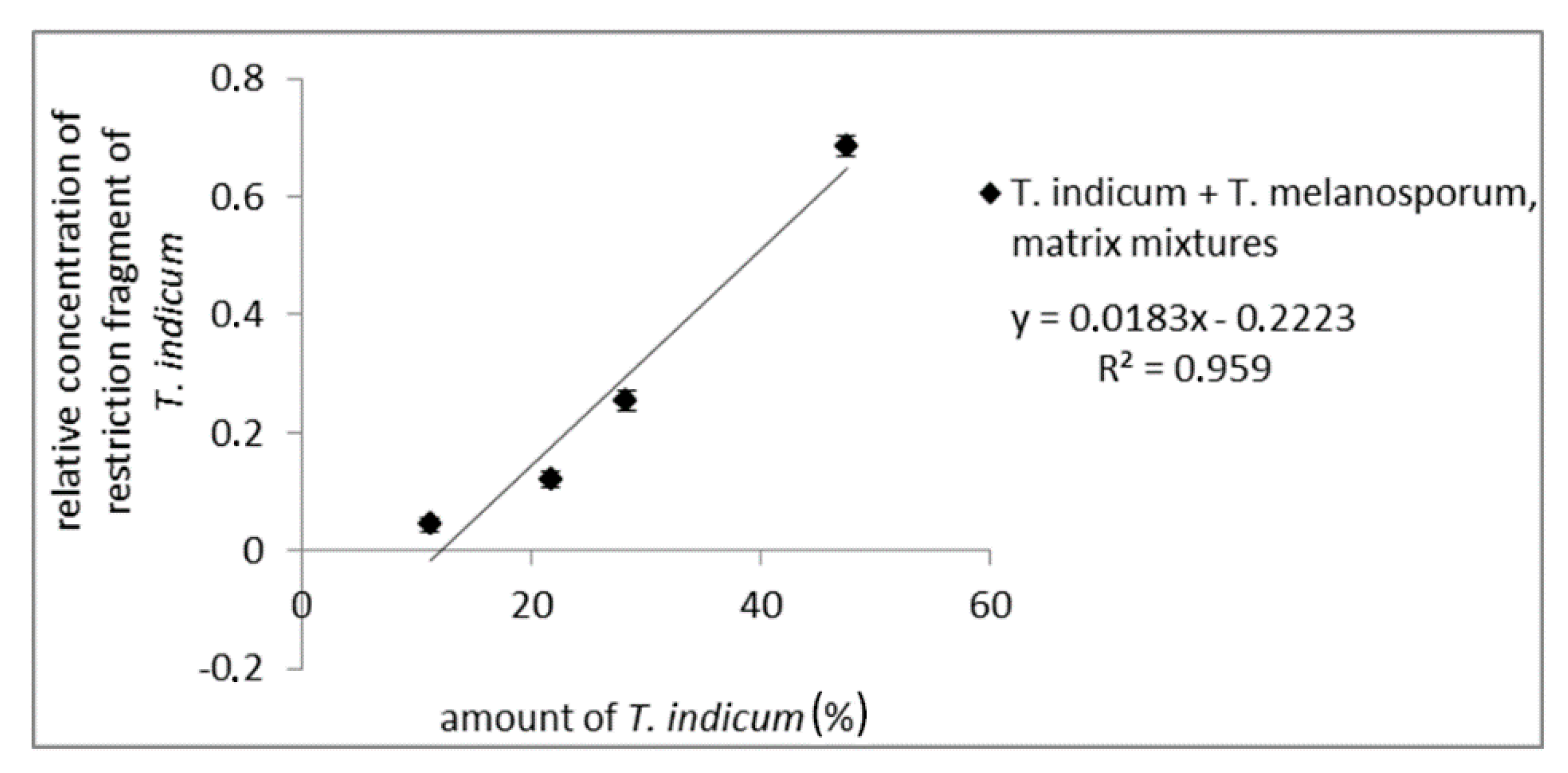
3.2. RFLP and CGE
3.2.1. PCR-Amplification of the ITS Region, Evaluation via CGE
3.2.2. RFLP of the ITS Region, Evaluation via CGE
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Trappe, J.M.; Molina, R.; Luoma, D.L.; Cázares, E.; Pilz, D.; Smith, J.E.; Castellano, M.A.; Miller, S.L. Diversity, ecology, and conservation of truffle fungi in forests of the Pacific Northwest. 2009. Available online: https://www.fs.fed.us/pnw/pubs/pnw_gtr772.pdf (accessed on 21 March 2020).
- Læssøe, T.; Hansen, K. Truffle trouble: What happened to the Tuberales? Mycol. Res. 2007, 111, 1075–1099. [Google Scholar] [CrossRef] [PubMed]
- Harley, J.L.; Smith, S.E. Mycorrhizal symbiosis; Academic Press Inc.: Lodon, UK, 1983. [Google Scholar]
- Giovannetti, G.; Fontana, A. Mycorrhizal synthesis between Cistaceae and Tuberaceae. New Phytol. 1982, 92, 533–537. [Google Scholar] [CrossRef]
- Mello, A.; Fontana, A.; Meotto, F.; Comandini, O.; Bonfante, P. Molecular and morphological characterization of Tuber magnatum mycorrhizas in a long-term survey. Microbiol. Res. 2001, 155, 279–284. [Google Scholar] [CrossRef]
- Zambonelli, A.; Iotti, M.; Murat, C. True Truffle (Tuber Spp.) in the World: Soil Ecology, Systematics and Biochemistry; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Bougher, N.L.; Lebel, T. Sequestrate (truffle-like) fungi of Australia and New Zealand. Aust. Syst. Bot. 2001, 14, 439–484. [Google Scholar] [CrossRef]
- Ceruti, A.; Fontana, A.; Nosenzo, C. Le specie europee del genere Tuber: Una revisione storica; Museo Regionale di Scienze Naturali di Torino: Torino, Italy, 2003. [Google Scholar]
- Chevalier, G.; Frochot, H. Ecology and possibility of culture in Europe of the Burgundy truffle (Tuber uncinatum Chatin). Agric. Ecosyst. Environ. 1990, 28, 71–73. [Google Scholar] [CrossRef]
- Pegler, D. Useful fungi of the world: Morels and truffles. Mycologist 2003, 17, 174–175. [Google Scholar] [CrossRef]
- Marjanovic, Z.; Saljnikov, E.; Milenkovic, M.; Grebenc, T. Ecological features of Tuber magnatum Pico in the conditions of West Balkans–soil characterization. In Proceedings of the 3rd international congress on truffles, Spoleto, Italy, 25–28 November 2008. [Google Scholar]
- Lazzari, B.; Gianazza, E.; Viotti, A. Molecular characterization of some truffle species. In Biotechnology of Ectomycorrhizae; Plenum Press: New York, NY, USA, 1995; pp. 161–169. [Google Scholar]
- Hall, I.R.; Yun, W.; Amicucci, A. Cultivation of edible ectomycorrhizal mushrooms. Trends Biotechnol. 2003, 21, 433–438. [Google Scholar] [CrossRef]
- Mello, A.; Murat, C.; Bonfante, P. Truffles: Much more than a prized and local fungal delicacy. FEMS Microbiol. Lett. 2006, 260, 1–8. [Google Scholar] [CrossRef]
- Favre, J.; Parguey Leduc, A.; Sejalon Delmas, N.; Dargent, R.; Kulifaj, M. The ascocarp of Tuber indicum (Chinese truffle) recently introduced in France: Preliminary study. C. r. hebd. séances Acad. sci. Serie 3 Sciences de la Vie (France) 1996, 319, 517–521. [Google Scholar]
- Bonito, G.; Trappe, J.M.; Donovan, S.; Vilgalys, R. The Asian black truffle Tuber indicum can form ectomycorrhizas with North American host plants and complete its life cycle in non-native soils. Fungal Ecol. 2011, 4, 83–93. [Google Scholar] [CrossRef]
- García-Montero, L.G.; Díaz, P.; Di Massimo, G.; García-Abril, A. A review of research on Chinese Tuber species. Mycol Prog. 2010, 9, 315–335. [Google Scholar] [CrossRef]
- Murat, C.; Zampieri, E.; Vizzini, A.; Bonfante, P. Is the Perigord black truffle threatened by an invasive species? We dreaded it and it has happened! New Phytol. 2008, 178, 699–702. [Google Scholar] [CrossRef] [PubMed]
- Paolocci, F.; Rubini, A.; Riccioni, C.; Granetti, B.; Arcioni, S. Cloning and characterization of two repeated sequences in the symbiotic fungus Tuber melanosporum Vitt. FEMS Microbiol. Ecol. 2000, 34, 139–146. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Séjalon-Delmas, N.; Roux, C.; Martins, M.; Kulifaj, M.; Bécard, G.; Dargent, R. Molecular tools for the identification of Tuber melanosporum in agroindustry. J. Agric. Food Chem. 2000, 48, 2608–2613. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Xu, J. Fungal DNA barcoding. Genome 2016, 59, 913–932. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- Haase, I.; Brüning, P.; Matissek, R.; Fischer, M. Real-time PCR assays for the quantitation of rDNA from apricot and other plant species in marzipan. J. Agric. Food Chem. 2013, 6, 3414–3418. [Google Scholar] [CrossRef]
- Johnson, S.M.; Carlson, E.L.; Pappagianis, D. Determination of ribosomal DNA copy number and comparison among strains of Coccidioides. Mycopathologia 2015, 179, 45–51. [Google Scholar] [CrossRef]
- López-Calleja, I.M.; de la Cruz, S.; Pegels, N.; González, I.; García, T.; Martín, R. High resolution TaqMan real-time PCR approach to detect hazelnut DNA encoding for ITS rDNA in foods. Food Chem. 2013, 141, 1872–1880. [Google Scholar] [CrossRef]
- Bertini, L.; Potenza, L.; Zambonelli, A.; Amicucci, A.; Stocchi, V. Restriction fragment length polymorphism species-specific patterns in the identification of white truffles. FEMS Microbiol. Lett. 1998, 164, 397–401. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gandeboeuf, D.; Dupre, C.; Chevalier, G.; Nicolas, P.; Roeckel-Drevet, P. Typing Tuber ectomycorrhizae by polymerase chain amplification of the internal transcribed spacer of rDNA and the sequence characterized amplified region markers. Can. J. Microbiol. 1997, 43, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Henrion, B.; Chevalier, G.; Martin, F. Typing truffle species by PCR amplification of the ribosomal DNA spacers. Mycol. Res. 1994, 98, 37–43. [Google Scholar] [CrossRef]
- Mabru, D.; Dupré, C.; Douet, J.; Leroy, P.; Ravel, C.; Ricard, J.; Medina, B.; Castroviejo, M.; Chevalier, G. Rapid molecular typing method for the reliable detection of Asiatic black truffle (Tuber indicum) in commercialized products: Fruiting bodies and mycorrhizal seedlings. Mycorrhiza. 2001, 11, 89–94. [Google Scholar] [CrossRef]
- Murat, C.; Vizzini, A.; Bonfante, P.; Mello, A. Morphological and molecular typing of the below-ground fungal community in a natural Tuber magnatum truffle-ground. FEMS Microbiol. Lett. 2005, 245, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Paolocci, F.; Rubini, A.; Granetti, B.; Arcioni, S. Typing Tuber melanosporum and Chinese black truffle species by molecular markers. FEMS Microbiol. Lett. 1997, 153, 255–260. [Google Scholar] [CrossRef]
- Paolocci, F.; Rubini, A.; Granetti, B.; Arcioni, S. Rapid molecular approach for a reliable identification of Tuber spp. ectomycorrhizae. FEMS Microbiol. Ecol. 1999, 28, 23–30. [Google Scholar] [CrossRef]
- Douet, J.; Castroviejo, M.; Mabru, D.; Chevalier, G.; Dupré, C.; Bergougnoux, F.; Ricard, J.; Médina, B. Rapid molecular typing of Tuber melanosporum, T. brumale and T. indicum from tree seedlings and canned truffles. Anal. Bioanal. Chem. 2004, 379, 668–673. [Google Scholar] [CrossRef]
- Gryndler, M.; Trilčová, J.; Hršelová, H.; Streiblová, E.; Gryndlerová, H.; Jansa, J. Tuber aestivum Vittad. mycelium quantified: Advantages and limitations of a qPCR approach. Mycorrhiza 2013, 23, 341–348. [Google Scholar] [CrossRef]
- Zampieri, E.; Rizzello, R.; Bonfante, P.; Mello, A. The detection of mating type genes of Tuber melanosporum in productive and non productive soils. Appl Soil Ecol. 2012, 57, 9–15. [Google Scholar] [CrossRef]
- Iotti, M.; Leonardi, M.; Lancellotti, E.; Salerni, E.; Oddis, M.; Leonardi, P.; Perini, C.; Pacioni, G.; Zambonelli, A. Spatio-temporal dynamic of Tuber magnatum mycelium in natural truffle grounds. PLoS ONE 2014, 9, e115921. [Google Scholar] [CrossRef] [PubMed]
- Salerni, E.; Iotti, M.; Leonardi, P.; Gardin, L.; D’Aguanno, M.; Perini, C.; Pacioni, P.; Zambonelli, A. Effects of soil tillage on Tuber magnatum development in natural truffières. Mycorrhiza 2014, 24, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Parladé, J.; De la Varga, H.; De Miguel, A.M.; Sáez, R.; Pera, J. Quantification of extraradical mycelium of Tuber melanosporum in soils from truffle orchards in northern Spain. Mycorrhiza 2013, 23, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, R.; Zampieri, E.; Vizzini, A.; Autino, A.; Cresti, M.; Bonfante, P.; Mello, A. Authentication of prized white and black truffles in processed products using quantitative real-time PCR. Food Res. Int. 2012, 48, 792–797. [Google Scholar] [CrossRef]
- Holland, P.M.; Abramson, R.D.; Watson, R.; Gelfand, D.H. Detection of specific polymerase chain reaction product by utilizing the 5’–3’exonuclease activity of Thermus aquaticus DNA polymerase. Proc. Natl. Acad. Sci. USA 1991, 88, 7276–7280. [Google Scholar] [CrossRef]
- Haunshi, S.; Pattanayak, A.; Bandyopadhaya, S.; Saxena, S.C.; Bujarbaruah, K.M. A simple and quick DNA extraction procedure for rapid diagnosis of sex of chicken and chicken embryos. J. Poult. Sci. 2008, 45, 75–81. [Google Scholar] [CrossRef]
- Bruüning, P.; Haase, I.; Matissek, R.; Fischer, M. Marzipan: Polymerase chain reaction-driven methods for authenticity control. J. Agric. Food Chem. 2011, 59, 11910–11917. [Google Scholar] [CrossRef] [PubMed]
- Debode, F.; Marien, A.; Janssen, É.; Bragard, C.; Berben, G. Influence of the amplicon length on real-time PCR results. Biotechnol., Agron., Soc. Environ. 2017, 21, 3–11. [Google Scholar]
- Cankar, K.; Štebih, D.; Dreo, T.; Žel, J.; Gruden, K. Critical points of DNA quantification by real-time PCR–effects of DNA extraction method and sample matrix on quantification of genetically modified organisms. BMC Biotechnol. 2006, 6, 37. [Google Scholar] [CrossRef]
- Paolocci, F.; Rubini, A.; Riccioni, C.; Topini, F.; Arcioni, S. Tuber aestivum and Tuber uncinatum: Two morphotypes or two species? FEMS Microbiol. Lett. 2004, 235, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Paolocci, F.; Angelini, P.; Cristofari, E.; Granetti, B.; Arcioni, S. Identification of Tuber spp and corresponding ectomycorrhizae through molecular markers. J. Agric. Food Chem. 1995, 69, 511–517. [Google Scholar] [CrossRef]
- Bonito, G.M.; Gryganskyi, A.P.; Trappe, J.M.; Vilgalys, R. A global meta-analysis of Tuber ITS rDNA sequences: Species diversity, host associations and long-distance dispersal. Mol. Ecol. 2010, 19, 4994–5008. [Google Scholar] [CrossRef] [PubMed]
- Roux, C.; Séjalon-Delmas, N.; Martins, M.; Parguey-Leduc, A.; Dargent, R.; Bécard, G. Phylogenetic relationships between European and Chinese truffles based on parsimony and distance analysis of ITS sequences. FEMS Microbiol. Lett. 1999, 180, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Qiao, P.; Tian, W.; Liu, P.; Yu, F.; Chen, J.; Deng, X.; Wan, S.; Wang, R.; Wang, Y.; Guo, H. Phylogeography and population genetic analyses reveal the speciation of the Tuber indicum complex. Fungal Genet. Biol. 2018, 113, 14–23. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tuber Species | Geographical Origin | Fruiting Bodies Analyzed | |
|---|---|---|---|
| Numbers with Regard to the Origin | Total Number | ||
| T. albidum Pico | Italy | 5 | 5 |
| T. indicum | China | 5 | 5 |
| T. himalayense | Dali, Yunnan, China | 20 | 20 |
| T. brumale | Sarrion, Teruel, Spain | 2 | 2 |
| T. melanosporum | Marche, Italy | 2 | |
| France | 1 | ||
| Australia | 2 | ||
| Sarrion, Teruel, Spain | 8 | ||
| Castello, Valencia, Spain | 6 | ||
| unknown | 1 | 20 | |
| T. magnatum | Romagna, Italy | 2 | |
| Buzet, Croatia | 1 | ||
| Turin, Piemonte, Italy | 1 | ||
| Italy | 5 | ||
| L’Aquila, Abruzzo, Italy | 1 | ||
| Perugia, Umbria, Italy | 1 | ||
| Rome, Lazio, Italy | 1 | ||
| Naples, Campania, Italy | 1 | ||
| Ancona, Marche, Italy | 1 | ||
| Campobasso, Molise, Italy | 1 | 15 | |
| T. aestivum | unknown | 19 | |
| Romania | 15 | ||
| Italy | 11 | ||
| Hungary | 3 | ||
| Toscana, Florence, Italy | 2 | 50 | |
| Processed food containing truffle: | |||
| T. melanosporum fruiting bodies canned in saltwater | 6 | ||
| salt with dried T. aestivum | 1 | ||
| T. brumale chopped and cooked in sherry port wine stock | 1 | ||
| Primer/Probes | Name | Sequence 5′–3′ | Product Size (bp) |
|---|---|---|---|
| specific for T. melanosporum | Primer Mela-fw Mela-rv Probe | ACGACGGACTTTATAAACGGTTATAA AGCGGGTATCCCTCCCTGATT Cy5–GACCTGGATCAGTCACAAGTCTTGTCTGGT-BHQ2 | 141 |
| specific for T. indicum/ T. himalayense | Primer Indi-fw ITS4LNG * Probe | AACAACAGACTTTGTAAAGGGTT TGATATGCTTAAGTTCAGCGGG HEX-GGACCTAGATCAGTCACAAGTCATGTCTGG-BHQ2 | 146 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schelm, S.; Siemt, M.; Pfeiffer, J.; Lang, C.; Tichy, H.-V.; Fischer, M. Food Authentication: Identification and Quantitation of Different Tuber Species via Capillary Gel Electrophoresis and Real-Time PCR. Foods 2020, 9, 501. https://doi.org/10.3390/foods9040501
Schelm S, Siemt M, Pfeiffer J, Lang C, Tichy H-V, Fischer M. Food Authentication: Identification and Quantitation of Different Tuber Species via Capillary Gel Electrophoresis and Real-Time PCR. Foods. 2020; 9(4):501. https://doi.org/10.3390/foods9040501
Chicago/Turabian StyleSchelm, Stefanie, Melanie Siemt, Janin Pfeiffer, Christina Lang, Hans-Volker Tichy, and Markus Fischer. 2020. "Food Authentication: Identification and Quantitation of Different Tuber Species via Capillary Gel Electrophoresis and Real-Time PCR" Foods 9, no. 4: 501. https://doi.org/10.3390/foods9040501
APA StyleSchelm, S., Siemt, M., Pfeiffer, J., Lang, C., Tichy, H.-V., & Fischer, M. (2020). Food Authentication: Identification and Quantitation of Different Tuber Species via Capillary Gel Electrophoresis and Real-Time PCR. Foods, 9(4), 501. https://doi.org/10.3390/foods9040501