Degree of Hydrolysis Affects the Techno-Functional Properties of Lesser Mealworm Protein Hydrolysates


Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Samples
2.2. Enzymatic Hydrolysis for Protein Extraction
2.2.1. Preliminary Enzymatic Assisted Extraction
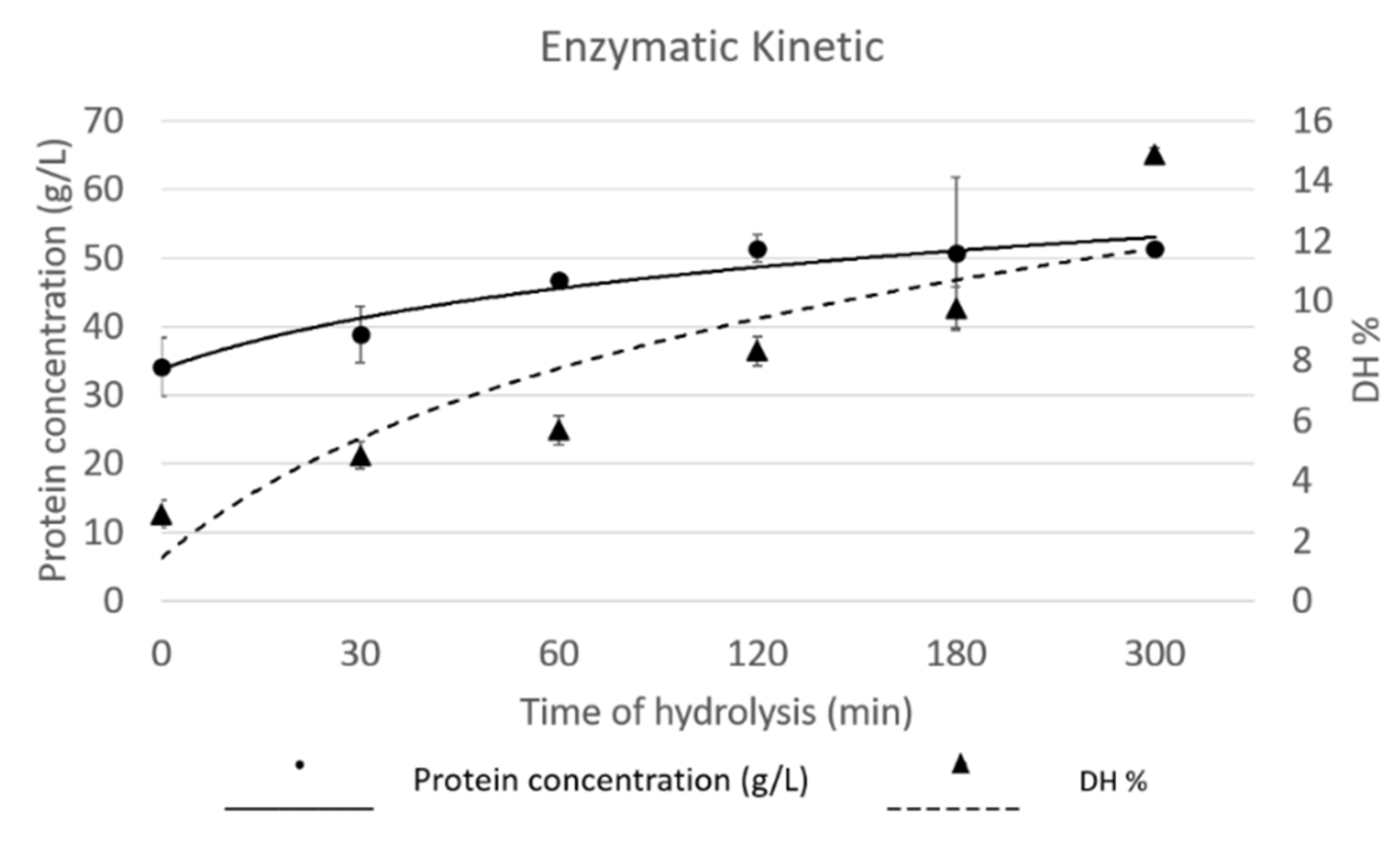
2.2.2. Set of Protein Hydrolysates Collected at Different Time-Points
2.2.3. Bulk Composition
2.2.4. Degree of Hydrolysis
2.3. Techno Functional Properties
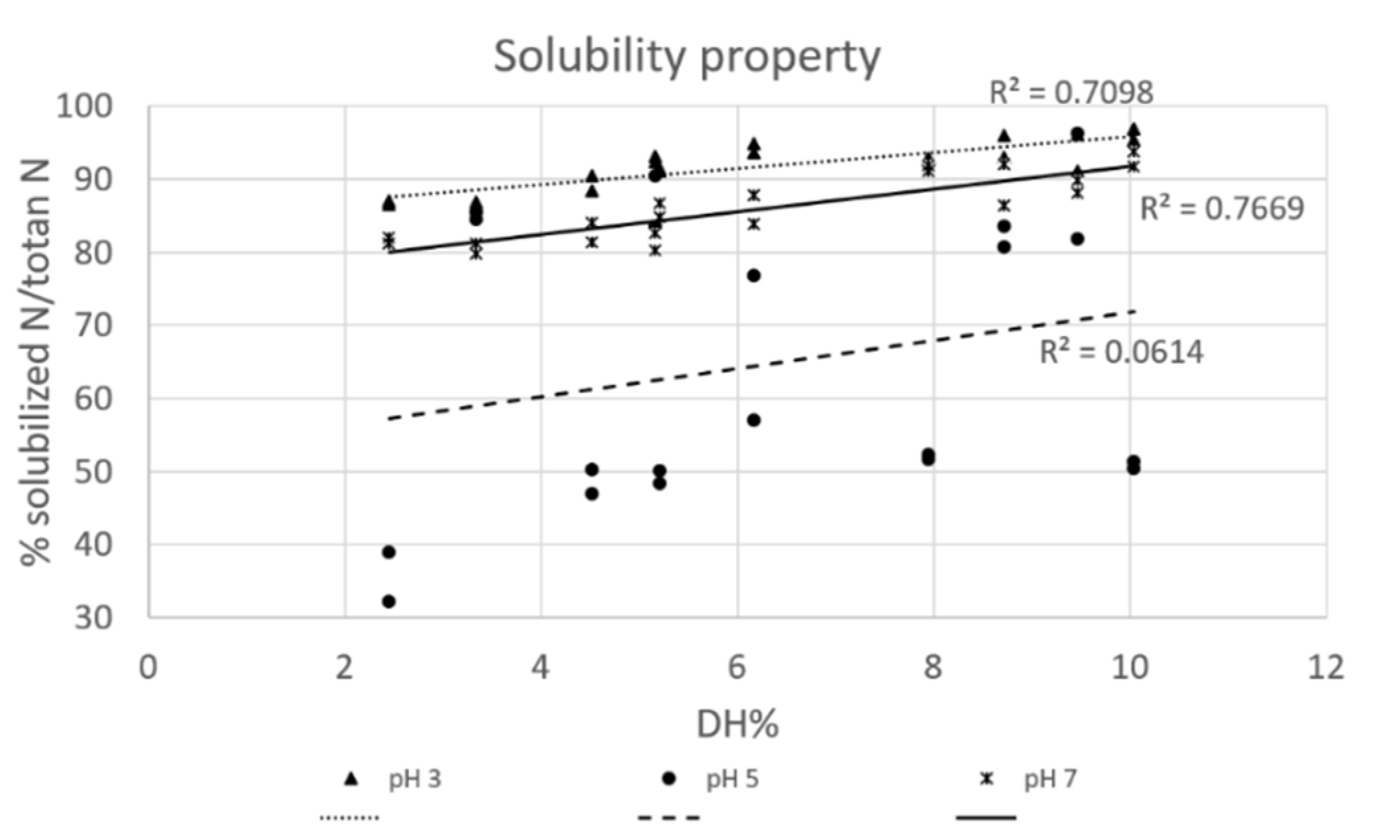
2.3.1. Solubility
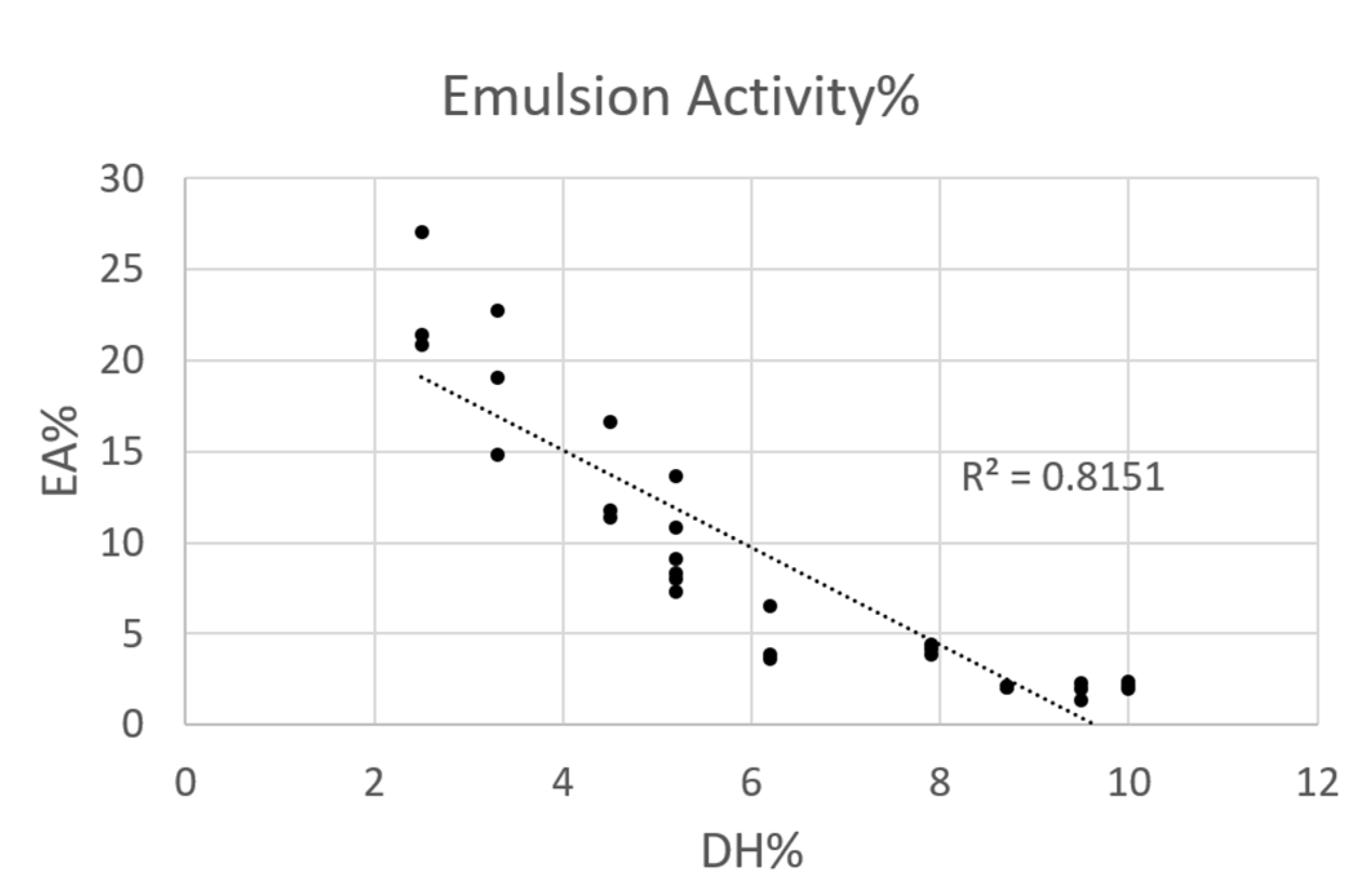
2.3.2. Emulsification Properties
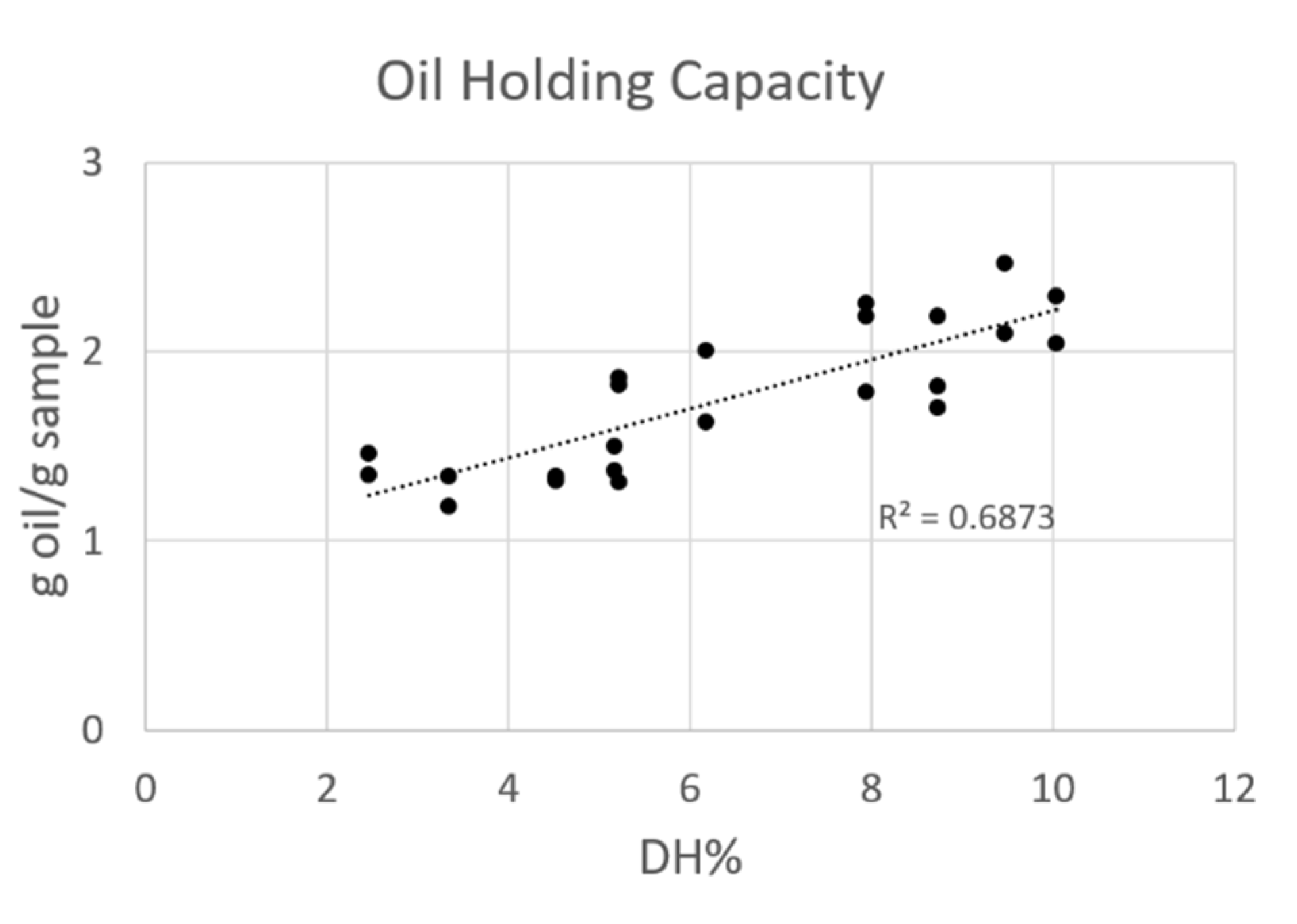
2.3.3. Oil Holding Capacity
2.3.4. Foaming Capacity
2.4. Statistical Analysis
3. Results
3.1. Techno Functional Assay of Protein Hydrolysate
3.2. Generation of Protein Hydrolysates with Different Degrees of Hydrolysis
3.3. Functional Properties
3.3.1. Solubility
3.3.2. Emulsifying Activity
3.3.3. Oil Holding Capacity
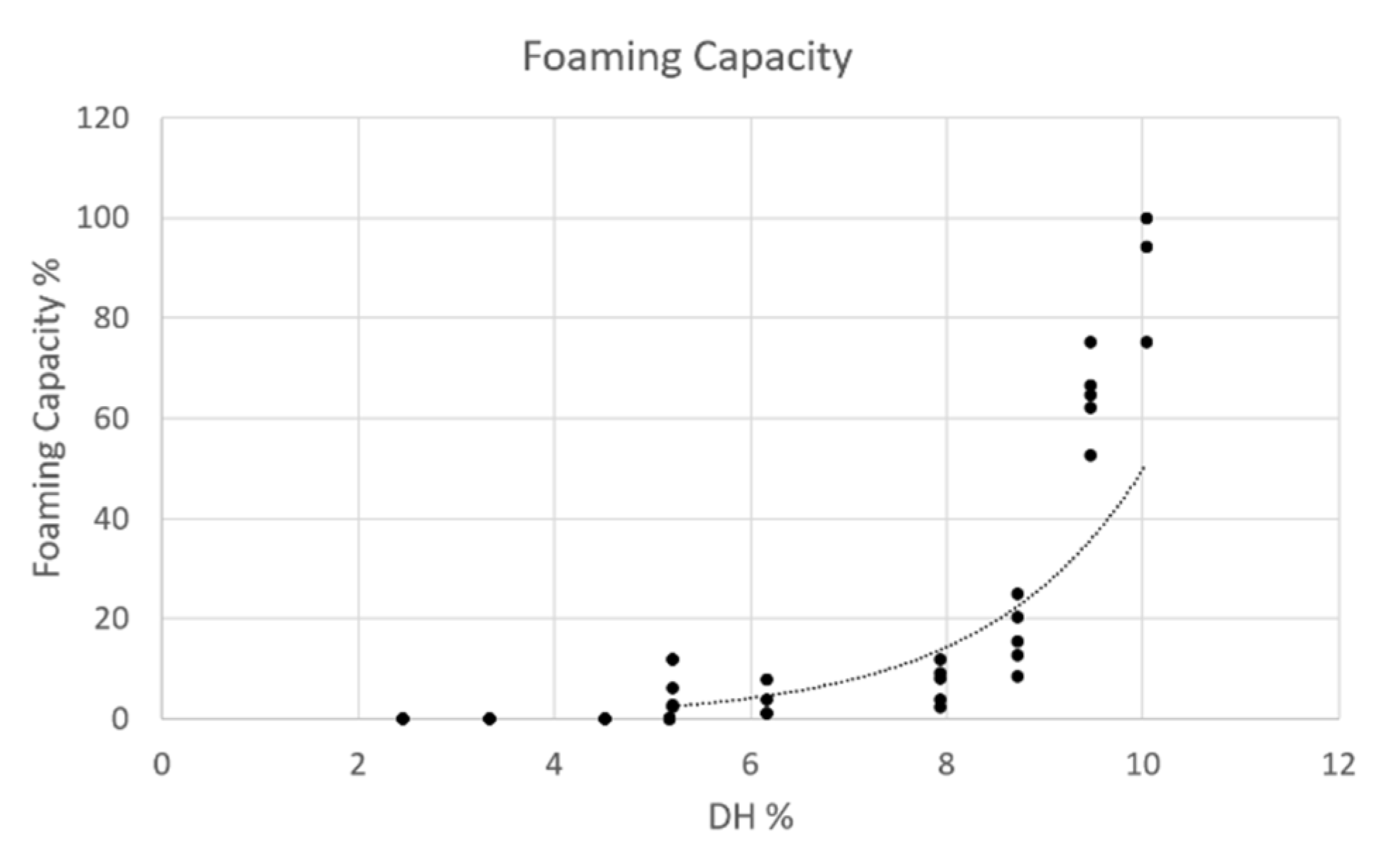
3.3.4. Foaming Capacity
3.4. Potential Application
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. How to Feed the World in 2050. Insights Expert Meet FAO 2009, 1, 1–35. [Google Scholar] [CrossRef]
- Boland, M.J.; Rae, A.N.; Vereijken, J.M.; Meuwissen, M.P.M.; Fischer, A.R.H.; van Boekel, M.A.J.S.; Hendriks, W.H. The future supply of animal-derived protein for human consumption. Trends Food Sci. Technol. 2013, 29, 62–73. [Google Scholar] [CrossRef]
- Yi, L.; Lakemond, C.M.M.; Sagis, L.M.C.; Eisner-Schadler, V.; van Huis, A.; Boekel, M.A.J.S.V. Extraction and characterisation of protein fractions from five insect species. Food Chem. 2013, 141, 3341–3348. [Google Scholar] [CrossRef] [PubMed]
- Caligiani, A.; Marseglia, A.; Leni, G.; Baldassarre, S.; Maistrello, L.; Dossena, A.; Sforza, S. Composition of black soldier fly prepupae and systematic approaches for extraction and fractionation of proteins, lipids and chitin. Food Res. Int. 2018, 105, 812–820. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Potential and challenges of insects as an innovative source for food and feed production. Innov. Food Sci. Emerg. Technol. 2013, 17, 1–11. [Google Scholar] [CrossRef]
- Regulation (EU) 2015/2283 of the European Parliament and of the Council of 25 November 2015 on novel foods, amending Regulation (EU) No 1169/2011 of the European Parliament and of the Council and repealing Regulation (EC) No 258/97 of the European Parliament and of the Council and Commission Regulation (EC) No 1852/2001. Off. J. Eur. Union 2015, L 327, 1–22. Available online: http://data.europa.eu/eli/reg/2015/2283/oj (accessed on 20 August 2019).
- Commission Regulation (EU) 2017/893 of 24 May 2017 amending Annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council and Annexes X, XIV and XV to Commission Regulation (EU) No 142/2011 as regards the provisions on processed animal protein. Off. J. Eur. Union 2017, L 138, 92–115.
- Regulation (EU) No 56/2013 of 16 January 2013 Amending Annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council Laying Down Rules for the Prevention, Control and Eradication of Certain Transmissible Spongiform Encephalopathies. Off. J. Eur. Union 2013, L 21, 3–16. Available online: http://data.europa.eu/eli/reg/2013/56/oj (accessed on 20 August 2019).
- Yi, L.; Van Boekel, M.A.J.S.; Lakemond, C.M.M. Extracting Tenebrio molitor protein while preventing browning: Effect of pH and NaCl on protein yield. J. Insects Food Feed 2017, 3, 21–31. [Google Scholar] [CrossRef]
- Bußler, S.; Rumpold, B.A.; Jander, E.; Rawel, H.M.; Schlüter, O.K. Recovery and techno-functionality of flours and proteins from two edible insect species: Mealworm (Tenebrio molitor) and black soldier fly (Hermetia illucens) larvae. Heliyon 2016, 2, e00218. [Google Scholar] [CrossRef]
- Zhao, X.; Vázquez-gutiérrez, J.L.; Johansson, D.P.; Landberg, R.; Langton, M. Yellow Mealworm Protein for Food Purposes—Extraction and Functional Properties. PLoS ONE 2016, 11, e0147791. [Google Scholar] [CrossRef] [PubMed]
- Soetemans, L.; Uyttebroek, M.; D’Hondt, E.; Bastiaens, L. Use of Organic Acids to Improve Fractionation of the Black Soldier Fly larvae juice into Lipid and Protein Enriched Fractions. Eur. Food Res. Technol. 2019, 245, 2257–2267. [Google Scholar] [CrossRef]
- Nielsen, P.N.; Olsen, H.S. Enzymic modification of food protein. In Enzymes in Food Technol.; Whitehurst, R.J., van Oort, M., Eds.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2002. [Google Scholar]
- Leni, G.; Soetemans, L.; Jacobs, J.; Depraetere, S.; Gianotten, N.; Bastiaens, L.; Caligiani, A.; Sforza, S. Protein hydrolysates from Alphitobius diaperinus and Hermetia illucens larvae treated with commercial proteases. J. Insects Food Feed. Accepted (2 March 2020).
- Tavano, O.L. Protein hydrolysis using proteases: An important tool for food biotechnology. J. Mol. Catal. B Enzym 2013, 90, 1–11. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Unlocking the biological potential of proteins from edible insects through enzymatic hydrolysis: A review. Innov. Food Sci. Emerg. Technol. 2017, 43, 239–252. [Google Scholar] [CrossRef]
- Purschke, B.; Meinlschmidt, P.; Horn, C.; Rieder, O.; Jäger, H. Improvement of techno-functional properties of edible insect protein from migratory locust by enzymatic hydrolysis. Eur. Food Res. Technol. 2018, 244, 999–1013. [Google Scholar] [CrossRef]
- Hall, F.G.; Jones, O.G.; O’Haire, M.E.; Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. 2017, 224, 414–422. [Google Scholar] [CrossRef]
- Butré, C.I.; Sforza, S.; Gruppen, H.; Wierenga, P.A. Introducing enzyme selectivity: A quantitative parameter to describe enzymatic protein hydrolysis. Anal. Bioanal. Chem. 2014, 406, 5827–5841. [Google Scholar] [CrossRef]
- Zayas, J.F. Solubility of Proteins. In Functionality of Proteins in Food; Springer: Berlin/Heidelberg, Germany, 1997. [Google Scholar]
- Ghribi, A.M.; Gafsi, I.M.; Sila, A.; Blecker, C.; Danthine, S.; Attia, H.; Bougatef, A.; Besbes, S. Effects of enzymatic hydrolysis on conformational and functional properties of chickpea protein isolate. Food Chem. 2015, 187, 322–330. [Google Scholar] [CrossRef]
- Gbogouri, G.A.; Linder, M.; Fanni, J.; Parmenter, M.C. Food Chemistry and Toxicology Influence of Hydrolysis Degree. J. Food Sci. 2004, 69, 615–622. [Google Scholar] [CrossRef]
- Bao, Z.; Zhao, Y.; Wang, X.; Chi, Y.J. Effects of degree of hydrolysis (DH) on the functional properties of egg yolk hydrolysate with alcalase. J. Food Sci. Technol. 2017, 54, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Nisov, A.; Ercili-Cura, D.; Nordlund, E. Limited hydrolysis of rice endosperm protein for improved techno-functional properties. Food Chem. 2020, 302, 125274. [Google Scholar] [CrossRef] [PubMed]
- Souissi, N.; Bougatef, A.; Triki-Ellouz, Y.; Nasri, M. Biochemical and functional properties of sardinella (Sardinetta aurita) by-product hydrolysates. Food Technol. Biotechnol. 2007, 45, 187–194. [Google Scholar]
- Van der Ven, C.; Gruppen, H.; De Bont, D.B.A.; Voragen, A.G.J. Emulsion properties of casein and whey protein hydrolysates and the relation with other hydrolysate characteristics. J. Agric. Food Chem. 2001, 49, 5005–5012. [Google Scholar] [CrossRef] [PubMed]
- Quaglia, G.B.; Orban, E. Influence of Enzymatic Hydrolysis on Structure and Emulsifying Properties of Sardine (Sardina pilchardus) Protein Hydrolysates. J. Food Sci. 1990, 55, 1571–1573. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Rasco, B.A. Fish protein hydrolysates: Production, biochemical and functional prop-erties. Crit. Rev. Food Sci. Nutr. 2000, 40, 43–81. [Google Scholar] [CrossRef] [PubMed]
- Mune, M.A. Functional Properties of Hydrolysates. J. Food Process. Preserv. 2015, 39, 2386–2392. [Google Scholar] [CrossRef]
- Wouters, A.G.B.; Rombouts, I.; Fierens, E.; Brijs, K.; Delcour, J.A. Relevance of the Functional Properties of Enzymatic Plant Protein Hydrolysates in Food Systems. Compr. Rev. Food Sci. Food Saf. 2016, 15, 786–800. [Google Scholar] [CrossRef]
- Kinsella, J.E.; Melachouris, N. Functional properties of proteins in foods: A survey. Crit. Rev. Food Sci. Nutr. 1976, 7, 219–280. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hydrolysis Time | Protein% on DM | Lipid% on DM | Ash% on DM | DH%° | Protein Concentration (g/L) |
|---|---|---|---|---|---|
| Preliminary test | |||||
| 300 min * | 58.2 ± 1.3 a | nd | nd | 14.9 ± 0.2 e | 51.39 ± 0.02 b |
| Detailed test | |||||
| Control (0 min) | 68.8 ± 4.9 b | 16.3 ± 0.9 ab | 10.7 ± 0.1 a | 2.9 ± 0.5 a | 34.2 ± 4.3 a |
| 30 min | 69.2 ± 4.5 bc | 10.9 ± 6.3 a | 9.3 ± 1.3 a | 4.8 ± 0.5 b | 38.9 ± 4.1 a |
| 60 min | 65.41 ± 1.05 bc | 12.7 ± 2.3 ab | 9.6 ± 0.8 a | 5.7 ± 0.5 b | 46.72 ± 1.01 ab |
| 120 min | 63.5 ± 1.7 cd | 16.04 ± 2.36 ab | 8.66 ± 1.98 a | 8.3 ± 0.5 c | 51.4 ± 1.9 ab |
| 180 min | 62.1 ± 0.3 d | 17.3 ± 2.6 b | 9.1 ± 0.6 a | 9.8 ± 0.7 d | 50.78 ± 10.96 ab |
| Sample | Protein Solubility% | Emulsification Activity% | Oil Holding Capacity g oil/g Sample | Foaming Capacity% | ||
|---|---|---|---|---|---|---|
| pH 3 | pH 5 | pH 7 | ||||
| Preliminary test | ||||||
| 300 min * | 94.8 ± 4.8 ab | 96.1 ± 4.2 b | 94.3 ± 0.6 d | 1.2 ± 0.3 a ** | 6.7 ± 0.6 d | 0 |
| Detailed test | ||||||
| Control | 86.7 ± 0.3 a | 60.3 ± 28.6 a | 81.04 ± 0.95 a | 20.99 ± 4.06 e ** | 1.4 ± 0.1 a | 0 |
| 30 min | 91.1 ± 2.1 ab | 67.8 ± 22.4 a | 82.1 ± 1.6 ab | 11.8 ± 3.1 d ** | 1.4 ± 0.1 a | 0 |
| 60 min | 92.7 ± 1.8 b | 58.1 ± 12.9 a | 85.8 ± 1.7 bc | 6.7 ± 2.7 c ** | 1.7 ± 0.3 ab | 5.3 ± 4.2 a |
| 120 min | 93.6 ± 1.6 b | 67.1 ± 17.4 a | 90.4 ± 2.7 cd | 3.1 ± 1.1 bc ** | 1.99 ± 0.25 bc | 12.59 ± 7.02 a |
| 180 min | 94.9 ± 2.6 b | 69.9 ± 2.6 a | 90.9 ± 2.5 d | 2.03 ± 0.36 b ** | 2.2 ± 0.2 c | 73.6 ± 16.1 c |
| Standard protein | ||||||
| Casein | nd | nd | nd | 34.3 ± 2.3 f | 1.52 ± 0.03 a | 43 ± 5 b |
| Egg white | 85.9 ± 2.6 a | 80.4 ± 4.3 a | 86.4 ± 1.5 bc | 52.5 ± 3.5 g | 1.1 ± 0.2 a | 123 ± 24 d |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leni, G.; Soetemans, L.; Caligiani, A.; Sforza, S.; Bastiaens, L. Degree of Hydrolysis Affects the Techno-Functional Properties of Lesser Mealworm Protein Hydrolysates. Foods 2020, 9, 381. https://doi.org/10.3390/foods9040381
Leni G, Soetemans L, Caligiani A, Sforza S, Bastiaens L. Degree of Hydrolysis Affects the Techno-Functional Properties of Lesser Mealworm Protein Hydrolysates. Foods. 2020; 9(4):381. https://doi.org/10.3390/foods9040381
Chicago/Turabian StyleLeni, Giulia, Lise Soetemans, Augusta Caligiani, Stefano Sforza, and Leen Bastiaens. 2020. "Degree of Hydrolysis Affects the Techno-Functional Properties of Lesser Mealworm Protein Hydrolysates" Foods 9, no. 4: 381. https://doi.org/10.3390/foods9040381
APA StyleLeni, G., Soetemans, L., Caligiani, A., Sforza, S., & Bastiaens, L. (2020). Degree of Hydrolysis Affects the Techno-Functional Properties of Lesser Mealworm Protein Hydrolysates. Foods, 9(4), 381. https://doi.org/10.3390/foods9040381