Potential of Germination in Selected Conditions to Improve the Nutritional and Bioactive Properties of Moringa (Moringa oleifera L.)

 , ,
, ,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Chemicals and Reagents
2.3. Germination Experiments
2.4. Proximate Composition
2.5. Thiamine and Riboflavin
2.6. Total and Individual GLS
2.7. GABA
2.8. Free Phenolic Compounds
2.9. Antioxidant Activity
2.10. Data Modelling and Optimization of Germination Conditions
2.11. Statistical Analysis
3. Results and Discussion
3.1. Effect of Germination Conditions on Proximate Composition of Moringa Sprouts
3.2. Effect of Germination Conditions on Vitamin B1 and B2 Contents in Moringa Sprouts
3.3. Effect of Germination Conditions on Total and Individual GLS Content in Moringa Sprouts
3.4. Effect of Germination Conditions on GABA Content in Moringa Sprouts
3.5. Effect of Germination Conditions on the Content of Free Phenolic Compounds (FPC) in Moringa Sprouts
3.6. Effect of Germination Conditions on Antioxidant Activity in Moringa Sprouts
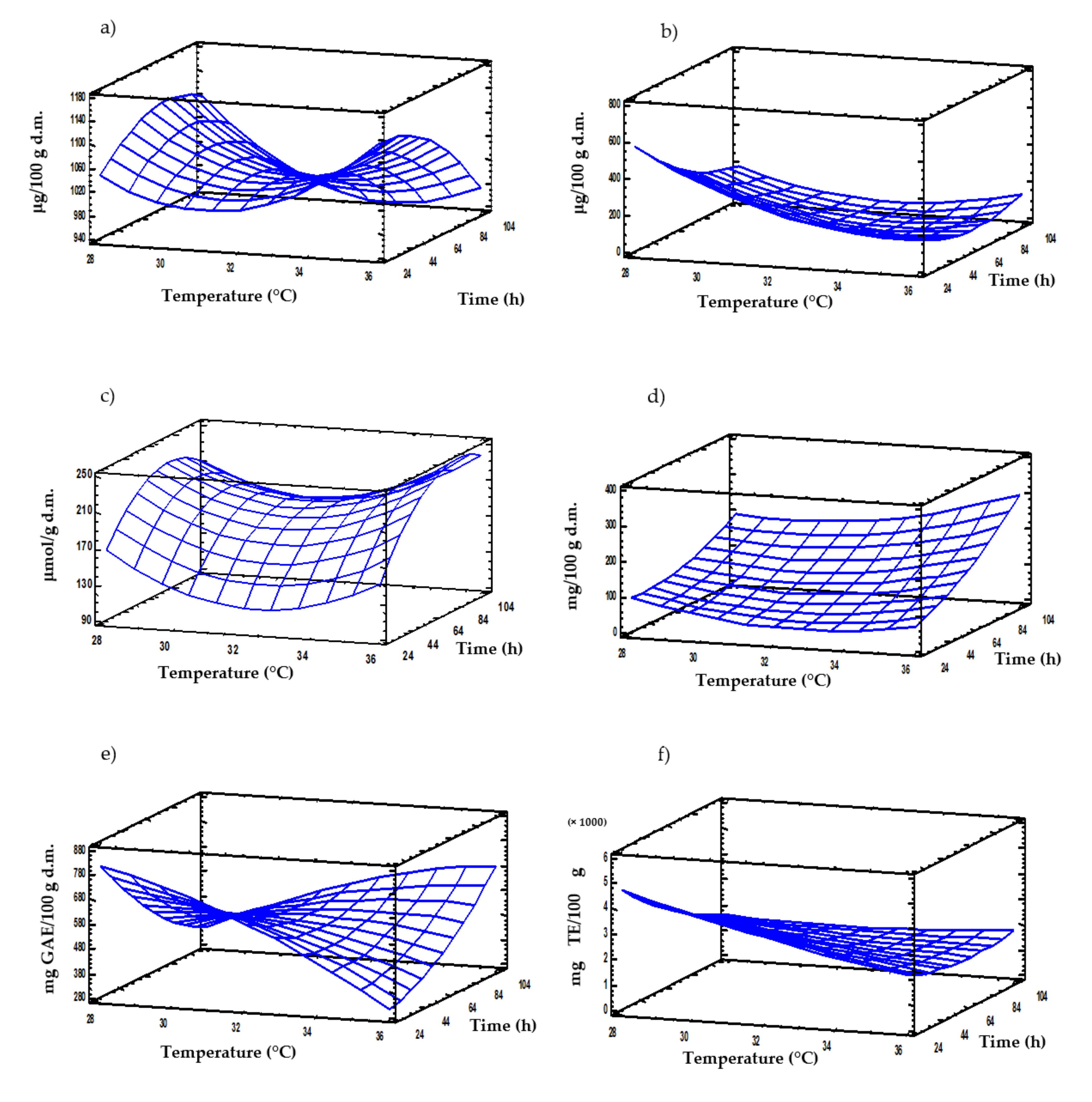
3.7. Multi-Response Modelling of the Influence Germination Conditions on Nutritional and Bioactive Properties of Moringa Sprouts
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gupta, S.; Jain, R.; Kachhwaha, S.; Kothari, S. Nutritional and Medicinal Applications of Moringa oleifera Lam.—Review of Current Status and Future Possibilities. J. Herb. Med. 2018, 11, 1–11. [Google Scholar] [CrossRef]
- Jaja-Chimedza, A.; Graf, B.L.; Simmler, C.; Kim, Y.; Kuhn, P.; Pauli, G.F.; Raskin, I. Biochemical Characterization and Anti-Inflammatory Properties of an Isothiocyanate-Enriched Moringa (Moringa oleifera) Seed Extract. PLoS ONE 2017, 12, e0182658. [Google Scholar] [CrossRef]
- Mehta, K.; Balaraman, R.; Amin, A.; Bafna, P.; Gulati, O. Effect of Fruits of Moringa oleifera on the Lipid Profile of Normal and Hypercholesterolaemic Rabbits. J. Ethnopharmacol. 2003, 86, 191–195. [Google Scholar] [CrossRef]
- Zabetakis, I.; Lordan, R.; Norton, C.; Tsoupras, A. COVID-19: The Inflammation Link and the Role of Nutrition in Potential Mitigation. Nutrients 2020, 12, 1466. [Google Scholar] [CrossRef]
- Guzmán-Albores, J.M.; Ramírez-Merchant, M.L.; Interiano, E.C.; Ruiz-Valdiviezo, V.M. Mebabolomic and Proteomic Analysis of Moringa oleifera Cultivated with Vermicompost and Phosphate Rock Under Water Stress Conditions. Int. J. Agric. Biol. 2019, 21, 786–794. [Google Scholar]
- Raman, J.K.; Alves, C.M.; Gnansounou, E. A Review on Moringa Tree and Vetiver Grass–Potential Biorefinery Feedstocks. Biores. Technol. 2018, 249, 1044–1051. [Google Scholar] [CrossRef]
- Falowo, A.B.; Mukumbo, F.E.; Idamokoro, E.M.; Lorenzo, J.M.; Afolayan, A.J.; Muchenje, V. Multi-Functional Application of Moringa oleifera Lam. in Nutrition and Animal Food Products: A Review. Food Res. Int. 2018, 106, 317–334. [Google Scholar] [CrossRef] [PubMed]
- Saa, R.W.; Fombang, E.N.; Ndjantou, E.B.; Njintang, N.Y. Treatments and Uses of Moringa oleifera Seeds in Human Nutrition: A Review. Food Sci. Nutr. 2019, 7, 1911–1919. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Wang, X.-J.; Zhang, Y.-Y.; Xing, Y.; Yang, L.; Ni, H.; Li, H.-H. Simultaneous Extraction and Separation of Oil, Proteins, and Glucosinolates from Moringa oleifera Seeds. Food Chem. 2019, 300, 125162. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yin, Q.; Yang, Y. Comprehensive Investigation of Moringa oleifera from Different Regions by Simultaneous Determination of 11 Polyphenols Using UPLC-ESI-MS/MS. Molecules 2020, 25, 676. [Google Scholar] [CrossRef]
- Nonogaki, H.; Bassel, G.W.; Bewley, J.D. Germination—Still a Mystery. Plant Sci. 2010, 179, 574–581. [Google Scholar] [CrossRef]
- Gan, R.-Y.; Lui, W.-Y.; Wu, K.; Chan, C.-L.; Dai, S.-H.; Sui, Z.-Q.; Corke, H. Bioactive Compounds and Bioactivities of Germinated Edible Seeds and Sprouts: An Updated Review. Trends Food Sci. Technol. 2017, 59, 1–14. [Google Scholar] [CrossRef]
- Rico, D.; Peñas, E.; García, M.C.; Martínez-Villaluenga, C.; Rai, D.K.; Birsan, R.I.; Frias, J.; Martin-Diana, A.B. Sprouted Barley Flour as a Nuttitious and Functional Ingredient. Foods 2020, 9, 296. [Google Scholar] [CrossRef] [PubMed]
- Cáceres, P.J.; Martínez-Villaluenga, C.; Amigo, L.; Frias, J. Maximising Thephytochemical Content and Antioxidant Activity of Ecuadorian Brown Rice Sprouts Through Optimal Germination Conditions. Food Chem. 2014, 152, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Aparicio-García, N.; Martínez-Villaluenga, C.; Frias, J.; Peñas, E. Changes in Protein Profile, Bioactive Potential and Enzymatic Activities of Gluten-Free Flours Obtained from Hulled and Dehulled Oat Varieties as Affected by Germination Conditions. LWT 2020, 134, 109955. [Google Scholar] [CrossRef]
- Paucar-Menacho, L.M.; Martínez-Villaluenga, C.; Dueñas, M.; Frias, J.; Peñas, E. Response Surface Optimisation of Germination Conditions to Improve the Accumulation of Bioactive Compounds and the Antioxidant Activity in Quinoa. Int. J. Food Sci. Technol. 2017, 53, 516–524. [Google Scholar] [CrossRef]
- Tomé-Sánchez, I.; Martín-Diana, A.B.; Peñas, E.; Bautista-Expósito, S.; Efrias, J.; Rico, D.; González-Maillo, L.; Martinez-Villaluenga, C. Soluble Phenolic Composition Tailored by Germination Conditions Accompany Antioxidant and Anti-Inflammatory Properties of Wheat. Antioxidants 2020, 9, 426. [Google Scholar] [CrossRef]
- Ijarotimi, O.S.; Adeoti, O.A.; Ariyo, O. Comparative Study on Nutrient Composition, Phytochemical, and Functional Characteristics of Raw, Germinated, and Fermented Moringa oleifera Seed Flour. Food Sci. Nutr. 2013, 1, 452–463. [Google Scholar] [CrossRef]
- Román, S.; Sánchez-Siles, L.M.; Siegrist, M. The Importance of Food Naturalness for Consumers: Results of a Systematic Review. Trends Food Sci. Technol. 2017, 67, 44–57. [Google Scholar] [CrossRef]
- Cáceres, P.J.; Martínez-Villaluenga, C.; Amigo, L.; Frias, J. Assessment on Proximate Composition, Dietary Fiber, Phytic Acid and Protein Hydrolysis of Germinated Ecuatorian Brown Rice. Plant Foods Hum. Nutr. 2014, 69, 261–267. [Google Scholar] [CrossRef]
- Frias, J.; Prodanov, M.; Sierra, I.; Vidal-Valverde, C. Effect of Light on Carbohydrates and Hydrosoluble Vitamins of Lentils during Soaking. J. Food Prot. 1995, 58, 692–695. [Google Scholar] [CrossRef] [PubMed]
- Kliebenstein, D.J.; Kroymann, J.; Brown, P.; Figuth, A.; Pedersen, D. Genetic control of natural variation in Arabidopsis glucosinolate accumulation. Plant Physiol. 2001, 126, 811–825. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Wang, C.; Li, S.; Chu, X.; Sun, K. Nutritional Compositions of Indian Moringa oleifera Seed and Antioxidant Activity of Its Polypeptides. Food Sci. Nutr. 2019, 7, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Olagbemide, P.T.; Alikwe, P.C.N. Proximate Analysis and Chemical Composition of Raw and Defatted Moringa oleifera Kernel. Adv. Life Sci. Technol. 2014, 24, 92–99. [Google Scholar]
- Eme, P.; Mbah, B.; Ogbusu, O. Effect of Cooking Methods (Boiling and Roasting) on Nutrients and Anti-nutrients Content of Moringa oleifera Seeds. Pak. J. Nutr. 2012, 11, 211–215. [Google Scholar] [CrossRef]
- Leone, A.; Spada, A.; Battezzati, A.; Schiraldi, A.; Aristil, J.; Bertoli, S. Moringa oleifera Seeds and Oil: Characteristics and Uses for Human Health. Int. J. Mol. Sci. 2016, 17, 2141. [Google Scholar] [CrossRef] [PubMed]
- Mbithi-Mwikya, S.; Van Camp, J.; Yiru, Y.; Huyghebaert, A. Nutrient and Antinutrient Changes in Finger Millet (Eleusine coracan) During Sprouting. LWT 2000, 33, 9–14. [Google Scholar] [CrossRef]
- Gopalakrishnan, L.; Doriya, K.; Kumar, D.S. Moringa oleifera: A Review on Nutritive Importance and Its Medicinal Application. Food Sci. Hum. Wellness 2016, 5, 49–56. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on Dietary Reference Values for Carbohydartes and Dietary Fibre. EFSA J. 2010, 8, 1462. [Google Scholar]
- Lemmens, E.; Moroni, A.V.; Pagand, J.; Heirbaut, P.; Ritala, A.; Karlen, Y.; Lê, K.; Broeck, H.C.V.D.; Brouns, F.J.; De Brier, N.; et al. Impact of Cereal Seed Sprouting on Its Nutritional and Technological Properties: A Critical Review. Compr. Rev. Food Sci. Food Saf. 2018, 18, 305–328. [Google Scholar] [CrossRef]
- Moongngarm, A.; Saetung, N. Comparison of Chemical Compositions and Bioactive Compounds of Germinated Rough Rice and Brown Rice. Food Chem. 2010, 122, 782–788. [Google Scholar] [CrossRef]
- Zieliński, H.; Frias, J.; Piskuła, M.K.; Kozłowska, H.; Vidal-Valverde, C. The Effect of Germination Process on the Superoxide Dismutase-Like Activity and Thiamine, Riboflavin and Mineral Contents of Rapeseeds. Food Chem. 2006, 99, 516–520. [Google Scholar] [CrossRef]
- Rahmatullina, Y.R.; Doronin, A.F.; Vrzhesinskaya, O.; Kodentsova, V. Content of Vitamins B1 and B2 in Germinating Grain. Bull. Exp. Biol. Med. 2013, 154, 628–630. [Google Scholar] [CrossRef] [PubMed]
- Bennett, R.N.; Mellon, F.A.; Foidl, N.; Pratt, J.H.; Dupont, M.S.; Perkins, L.; Kroon, P.A. Profiling Glucosinolates and Phenolics in Vegetative and Reproductive Tissues of the Multi-Purpose Trees Moringa oleifera L. (Horseradish Tree) and Moringa stenopetala L. J. Agric. Food Chem. 2003, 51, 3546–3553. [Google Scholar] [CrossRef]
- Förster, N.; Ulrichs, C.; Schreiner, M.; Müller, C.T.; Mewis, I. Development of a Reliable Extraction and Quantification Method for Glucosinolates in Moringa oleifera. Food Chem. 2015, 166, 456–464. [Google Scholar] [CrossRef]
- Maldini, M.; Maksoud, S.A.; Natella, F.; Montoro, P.; Petretto, G.L.; Foddai, M.; De Nicola, G.R.; Chessa, M.; Pintore, G. Moringa oleifera: Study of Phenoics and Glucosinaltes by Mass Spectrometry. J. Mass. Spectr. 2014, 49, 900–910. [Google Scholar] [CrossRef]
- Sikorska-Zimny, K.; Beneduce, L. The Glucosinolates and Their Bioactive Derivatives in Brassica: A Review on Classification, Biosynthesis and Content in Plant Tissues, Fate During and After Processing, Effect on the Human Organism and Interaction with the Gut Microbiota. Crit. Rev. Food Sci. Nutr. 2020, 1–28. [Google Scholar] [CrossRef]
- Miękus, N.; Marszałek, K.; Podlacha, M.; Iqbal, A.; Puchalski, C.; Swiergiel, A.H. Health Benefits of Plant-Derived Sulfur Compounds, Glucosinolates, and Organosulfur Compounds. Molecules 2020, 25, 3804. [Google Scholar] [CrossRef]
- Bellostas, N.; Sørensen, A.D.; Sørensen, J.C.; Sørensen, H. Genetic Variation and Metabolism of Glucosinolates. Adv. Bot. Res. 2007, 45, 369–415. [Google Scholar] [CrossRef]
- Olszewska, M.A.; Granica, S.; Kolodziejczyk-Czepas, J.; Magiera, A.; Czerwińska, M.E.; Nowak, P.; Rutkowska, M.; Wasiński, P.; Owczarek, A. Variability of Sinapic Acid Derivatives During Germination and Their Contribution to Antioxidant and Anti-Inflammatory Effects of Broccoli Sprouts on Human Plasma and Human Peripheral Blood Mononuclear Cells. Food Funct. 2020, 11, 7231. [Google Scholar] [CrossRef]
- Ragusa, L.; Picchi, V.; Tribulato, A.; Cavallaro, C.; Scalzo, R.L.; Branca, F. The Effect of the Germination Temperature on the Phytochemical Content of Broccoli and Rocket Sprouts. Int. J. Food Sci. Nutr. 2016, 68, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Andini, S.; Dekker, P.; Gruppen, H.; Araya-Cloutier, C.; Vincken, J.P. Modulation of Glucosinolate Composition in Brassicaceae Seeds by Germinatin and Fungal Elicitaion. J. Agric. Food Chem. 2019, 67, 12770–12779. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, Y.; Leng, C.; Li, X.; Wang, P.; Gu, Z.; Yang, R. Glucosinolates Metabolism and Redox State of Rocket (Eruca sativa Mill.) During Germination. J. Food Process. Preserv. 2019, 43, e14019. [Google Scholar] [CrossRef]
- Li, L.; Song, S.; Nirasawa, S.; Hung, Y.C.; Jiang, Z.; Liu, H. Slightly Acidic Electrolyzed Water Treatment Enhances the Main Bioctive Phytochemicals Content in Broccoli Sprouts via Changing Metabolism. J. Agric. Food Chem. 2019, 67, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Maina, S.; Misinzo, G.; Bakari, G.; Kim, H.-Y. Human, Animal and Plant Health Benefits of Glucosinolates and Strategies for Enhanced Bioactivity: A Systematic Review. Molecules 2020, 25, 3682. [Google Scholar] [CrossRef]
- Galuppo, M.; Giacoppo, S.; De Nicola, G.R.; Iori, R.; Navarra, M.; Lombardo, G.E.; Bramanti, P.; Mazzon, E. Antiinflammatory Activity of Glucomoringin Isothocyanate in a Mouse Model of Experimental Autoimmune Encephalomyelitis. Fitoterapia 2014, 95, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Galuppo, M.; De Nicola, G.; Iori, R.; Dell’Utri, P.; Bramanti, P.; Mazzon, E. Antibacterial Activity of Glucomoringin Bioactivated with Myrosinase against Two Important Pathogens Affecting the Health of Long-Term Patients in Hospitals. Molecules 2013, 18, 14340–14348. [Google Scholar] [CrossRef]
- Boscaro, V.; Boffa, L.; Binello, A.; Amisano, G.; Fornasero, S.; Cravotto, G.; Gallicchio, M. Antiproliferative, Proapoptotic, Antioxidant and Antimicrobial Effects of Sinapis nigra L. and Sinapis alga L. Extracts. Molecules 2018, 23, 3004. [Google Scholar] [CrossRef]
- Cho, H.J.; Lim, D.Y.; Kwon, G.T.; Kim, J.H.; Huang, Z.; Song, H.; Oh, Y.S.; Kang, Y.H.; Lee, K.W.; Dong, Z.; et al. Benzyl Isothiocyanate Inhibits Prostate Cancer Dvelopment in the Transgenic Adenocarcinoma Mouse Prostate (TRAMP) Model, Which Is Associated with the Induction of Cell Cycle G1 Arrest. Int. J. Mol. Sci. 2016, 17, 264. [Google Scholar] [CrossRef]
- Kim, E.J.; Hong, J.E.; Eom, S.J.; Lee, J.Y.; Park, J.H.Y. Oral Administration of Benzyl-Isothiocyanate Inhibits Solid Tumor Grow and Lung Metastasis of 4t1 Murine Mammary Carcinoama Cells in BALB/c Mice. Breast Cancer Res. Treat. 2011, 130, 61–71. [Google Scholar] [CrossRef]
- Luo, X.; Wang, Y.; Li, Q.; Wang, D.; Xing, C.; Zhang, L.; Xu, T.; Fang, F.; Wang, F. Accumulating Mechanism of γ-Aminobutyric Acid in Soybean (Glycine Max L.) During Germination. Int. J. Food Sci. Technol. 2017, 53, 106–111. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, E.J.; Lim, S.-T.; Han, J.-A. Self-enhancement of GABA in Rice Bran Using Various Stress Treatments. Food Chem. 2015, 172, 657–662. [Google Scholar] [CrossRef] [PubMed]
- León-López, L.; Bañuelos-Piña, A.M.; Reyes-Moreno, C.; Milán-Carrillo, J.; Contreras-Andrade, I.; Sánchez-Magaña, L.M.; Cuevas-Rodríguez, E.O. Optimisation of Temperature and Time for the Dark Germination Bioprocess of Moringa oleifera Seeds to Boost Nutritional Value, Total Phenolic Content and Antioxidant Activity. Int. Food Res. J. 2019, 26, 831–839. [Google Scholar]
- Pająk, P.; Socha, R.; Broniek, J.; Królikowska, K.; Fortuna, T. Antioxidant Properties, Phenolic and Mineral Composition of Germinated Chia, Golden Flax, Evening Primrose, Phacelia and Fenugreek. Food Chem. 2019, 275, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F. Chemical Composition and Food Uses of Teff (Eragrostis tef). Food Chem. 2018, 239, 402–415. [Google Scholar] [CrossRef]
- Cho, D.-H.; Lim, S.-T. Changes in Phenolic Acid Composition and Associated Enzyme Activity in Shoot and Kernel Fractions of Brown Rice During Germination. Food Chem. 2018, 256, 163–170. [Google Scholar] [CrossRef]
- Tesfay, S.Z.; Modi, A.; Mohammed, F. The Effect of Temperature in Moringa Seed Phytochemical Compounds and Carbohydrate Mobilization. South Afr. J. Bot. 2016, 102, 190–196. [Google Scholar] [CrossRef]
- Sharma, S.; Saxena, D.C.; Riar, C.S. Changes in the GABA And Polyphenols Contents of Foxtail Millet on Germination and Their Relationship with In Vitro Antioxidant Activity. Food Chem. 2018, 245, 863–870. [Google Scholar] [CrossRef]
- Chen, K.; Arora, R. Dynamics of the Antioxidant System During Seed Osmopriming, Post-Priming Germination, and Seedling Establishment in Spinach (Spinacia oleracea). Plant Sci. 2011, 180, 212–220. [Google Scholar] [CrossRef]
- Limmongkon, A.; Nopprang, P.; Chaikeandee, P.; Somboon, T.; Wongshaya, P.; Pilaisangsuree, V. LC-MS/MS Profiles and Interrelationships Between the Anti-Inflammatory Activity, Total Phenolic Content and Antioxidant Potential of Kalasin 2 Cultivar Peanut Sprout Crude Extract. Food Chem. 2018, 239, 569–578. [Google Scholar] [CrossRef]
- Meier, K.; Ehbrecht, M.D.; Wittstock, U. Glucosinolate Content in Dormant and Germinating Arabidopsis thaliana Seeds Is Affected by Non-Functional Alleles of Classical Myrosinase and Nitrile-Specifier Protein Genes. Front. Plant Sci. 2019, 10, 10. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | T (°C) | t (h) | Ash | Protein | Lipids | Fiber | Total Carbohydrates |
|---|---|---|---|---|---|---|---|
| Seed | - | - | 3.34 ± 0.00a | 28.01 ± 0.63e | 28.73 ± 0.04cd | 24.41 ± 1.24d | 35.32 ± 0.83a |
| Sprouts | 28 | 24 | 2.98 ± 0.03c | 29.59 ± 0.66bc | 30.13 ± 0.04bc | 28.32 ± 1.59ab | 34.45 ± 0.73a |
| 60 | 3.14 ± 0.03b | 30.64 ± 0.50b | 31.89 ± 0.30ab | 29.72 ± 2.09a | 29.25 ± 0.24bc | ||
| 96 | 2.99 ± 0.02c | 28.43 ± 0.97e | 29.26 ± 1.05cd | 26.92 ± 0.05c | 34.79 ± 2.05a | ||
| 32 | 24 | 3.19 ± 0.06b | 30.04 ± 0.70bc | 32.08 ± 1.14ab | 26.12 ± 1.56bcd | 29.54 ± 1.84bcd | |
| 60 | 3.20 ± 0.40b | 31.86 ± 0.73a | 33.39 ± 1.29a | 26.20 ± 2.37bcd | 28.00 ± 1.14cd | ||
| 96 | 3.17 ± 0.11b | 32.21 ± 0.46a | 33.18 ± 0.50a | 27.11 ± 1.61abcd | 26.84 ± 1.02d | ||
| 36 | 24 | 2.83 ± 0.05d | 28.80 ± 0.06de | 30.40 ± 1.33bc | 28.01 ± 0.74ab | 34.04 ± 1.28a | |
| 60 | 3.13 ± 0.02b | 31.79 ± 0.43a | 30.27 ± 0.38bc | 23.97 ± 0.27d | 30.11 ± 0.42b | ||
| 96 | 3.14 ± 0.00b | 28.99 ± 0.20cde | 27.87 ± 1.53d | 27.77 ± 1.75abc | 35.01 ± 1.31a |
| Sample | T (°C) | t (h) | Thiamine | Riboflavin |
|---|---|---|---|---|
| Seed | - | - | 1113.5 ± 18.8a | 179.11 ± 6.4de |
| Sprouts | 28 | 24 | 1090.8 ± 35.3a | 625.20 ± 52.1a |
| 60 | 1099.4 ± 5.0a | 221.27 ± 10.8b | ||
| 96 | 1112.0 ± 7.3a | 215.54 ± 8.4bc | ||
| 32 | 24 | 995.4 ± 8.6b | 152.80 ± 13.0d | |
| 60 | 1005.7 ± 50.3b | 153.95 ± 12.5d | ||
| 96 | 979.7 ± 8.0b | 81.50 ± 10.9f | ||
| 36 | 24 | 1113.7 ± 51.9a | 193.33 ± 17.1cd | |
| 60 | 1090.2 ± 30.3a | 115.10 ± 13.6e | ||
| 96 | 1001.9 ± 27.4b | 65.40 ± 3.8g |
| Sample | T (°C) | t (h) | Glucomoringin | Sinalbin | Glucotropaeolin | Total GLS |
|---|---|---|---|---|---|---|
| Seed | - | - | 197.4 ± 0.2d | 7.10 ± 0.29a | NDf | 204.5 ± 0.1c |
| Sprouts | 28 | 24 | 90.0 ± 1.2g | 4.11 ± 0.05b | NDf | 94.1 ± 1.2e |
| 60 | 176.7 ± 1.9e | 3.23 ± 0.46c | 0.90 ± 0.02d | 180.8 ± 2.4d | ||
| 96 | 205.1 ± 0.9c | 2.62 ± 0.03d | 1.28 ± 0.16c | 209.02 ± 0.8c | ||
| 32 | 24 | 234.9 ± 0.6a | 2.64 ± 0.02d | NDf | 237.5 ± 0.6a | |
| 60 | 214.5 ± 3.3b | 2.56 ± 0.08de | 0.38 ± 0.11e | 217.5 ± 3.2b | ||
| 96 | 170.2 ± 3.5f | 2.51 ± 0.20de | 3.69 ± 0.70b | 176.4 ± 3.0d | ||
| 36 | 24 | 202.4 ± 6.9cd | 2.47 ± 0.08de | NDf | 204.9 ± 6.9c | |
| 60 | 232.9 ± 6.5a | 2.58 ± 0.10de | 0.39 ± 0.01e | 235.8 ± 6.6a | ||
| 96 | 171.4 ± 1.1ef | 2.23 ± 0.13e | 4.88 ± 0.31a | 181.3 ± 5.5d |
| Sample | T (°C) | t (h) | GABA | FPC | Antioxidant Activity |
|---|---|---|---|---|---|
| Seed | - | - | 50.1 ± 2.2f | 418.0 ± 32.0e | 1531.4 ± 129.1g |
| Sprouts | 28 | 24 | 97.6 ± 9.6d | 753.5 ± 61.0a | 5172.9 ± 522.3a |
| 60 | 101.8 ± 4.8d | 646.9 ± 40.2b | 2199.9 ± 170.9de | ||
| 96 | 197.6 ± 19.9b | 335.7 ± 22.6f | 1904.7 ± 171.4f | ||
| 32 | 24 | 33.9 ± 3.6g | 593.5 ± 50.7c | 2541.9 ± 245.4b | |
| 60 | 129.2 ± 7.8c | 562.0 ± 46.8d | 2467.0 ± 237.3bc | ||
| 96 | 136.2 ± 20.5c | 576.1 ± 52.8cd | 2333.4 ± 196.0cd | ||
| 36 | 24 | 75.0 ± 7.3e | 340.8 ± 33.3f | 2124.1 ± 203.1ef | |
| 60 | 107.0 ± 9.3d | 324.2 ± 21.4f | 2097.3 ± 133.1ef | ||
| 96 | 291.2 ± 19.9a | 645.7 ± 35.9f | 1108.1 ± 111.1h |
| Optimal Conditions | ||||||
|---|---|---|---|---|---|---|
| Response | Predicted Model | R2 | T (°C) | t (h) | Predicted Values | Experimental Values |
| Thiamine | Y(T,t) = 5978.36 − 325.97T + 8.72 t + 5.37T2 − 0.29T × t | 0.83 | 36 | 24 | 1156.6 | 1113.7 ± 51.9 |
| Riboflavin | Y(T,t) = 10,034.0 − 536.61T − 26.47 t + 7.41T2 + 0.54 T × t + 0.05t2 | 0.83 | 28 | 24 | 577.7 | 625.2 ± 52.1 |
| GLS | Y(T,t) = 3057.2 − 187.21T + 1.72 t + 2.83T2 + 0.07 T × t − 0.03t2 | 0.87 | 36 | 96 | 246.9 | 178.55 ± 0.6 |
| GABA | Y(T,t) = 3494.6 − 204.17T − 6.84t + 3.01T2 + 0.20T × t + 0.02t2 | 0.83 | 36 | 96 | 255.7 | 291.2 ± 19.9 |
| FPC | Y(T,t) = 846.17 + 91.88T − 48.63t − 2.98T2 + 1.35T × t + 0.04t2 | 0.86 | 28 | 24 | 840.4 | 753.5 ± 61.0 |
| AO | Y(T,t) = 23,028.9 − 755.53T − 178.11t + 5.26T2 + 4.25T × t + 0.17t2 | 0.76 | 28 | 24 | 4671 | 5172 ± 522 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coello, K.E.; Frias, J.; Martínez-Villaluenga, C.; Cartea, M.E.; Abilleira, R.; Peñas, E. Potential of Germination in Selected Conditions to Improve the Nutritional and Bioactive Properties of Moringa (Moringa oleifera L.). Foods 2020, 9, 1639. https://doi.org/10.3390/foods9111639
Coello KE, Frias J, Martínez-Villaluenga C, Cartea ME, Abilleira R, Peñas E. Potential of Germination in Selected Conditions to Improve the Nutritional and Bioactive Properties of Moringa (Moringa oleifera L.). Foods. 2020; 9(11):1639. https://doi.org/10.3390/foods9111639
Chicago/Turabian StyleCoello, Karín E., Juana Frias, Cristina Martínez-Villaluenga, María Elena Cartea, Rosaura Abilleira, and Elena Peñas. 2020. "Potential of Germination in Selected Conditions to Improve the Nutritional and Bioactive Properties of Moringa (Moringa oleifera L.)" Foods 9, no. 11: 1639. https://doi.org/10.3390/foods9111639
APA StyleCoello, K. E., Frias, J., Martínez-Villaluenga, C., Cartea, M. E., Abilleira, R., & Peñas, E. (2020). Potential of Germination in Selected Conditions to Improve the Nutritional and Bioactive Properties of Moringa (Moringa oleifera L.). Foods, 9(11), 1639. https://doi.org/10.3390/foods9111639