Dominance of S. cerevisiae Commercial Starter Strains during Greco di Tufo and Aglianico Wine Fermentations and Evaluation of Oenological Performances of Some Indigenous/Residential Strains




Abstract
1. Introduction
2. Materials and Methods
2.1. Identification and Characterization of Commercial Starter Strains
2.2. Wine Fermentation Protocols
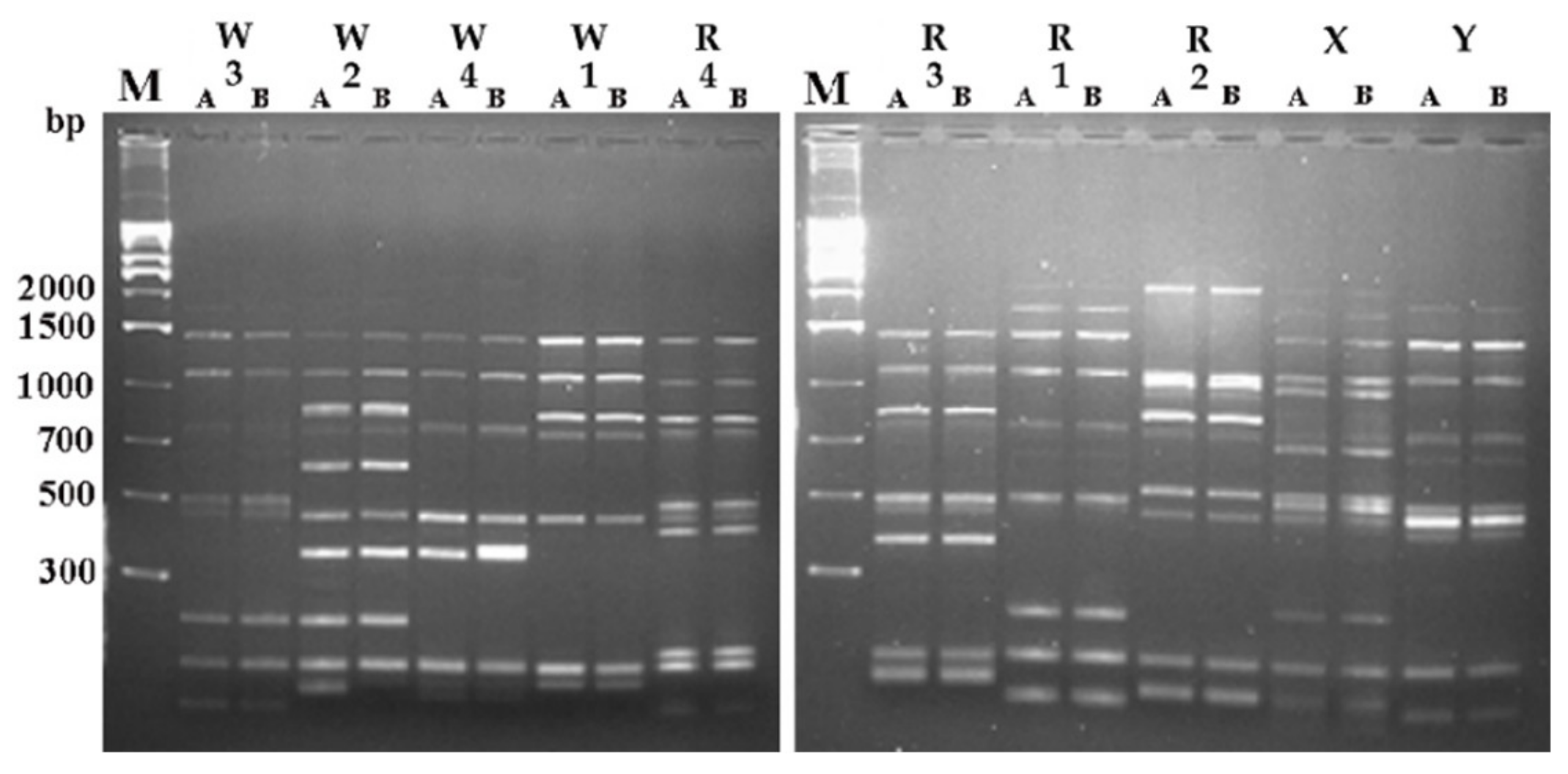
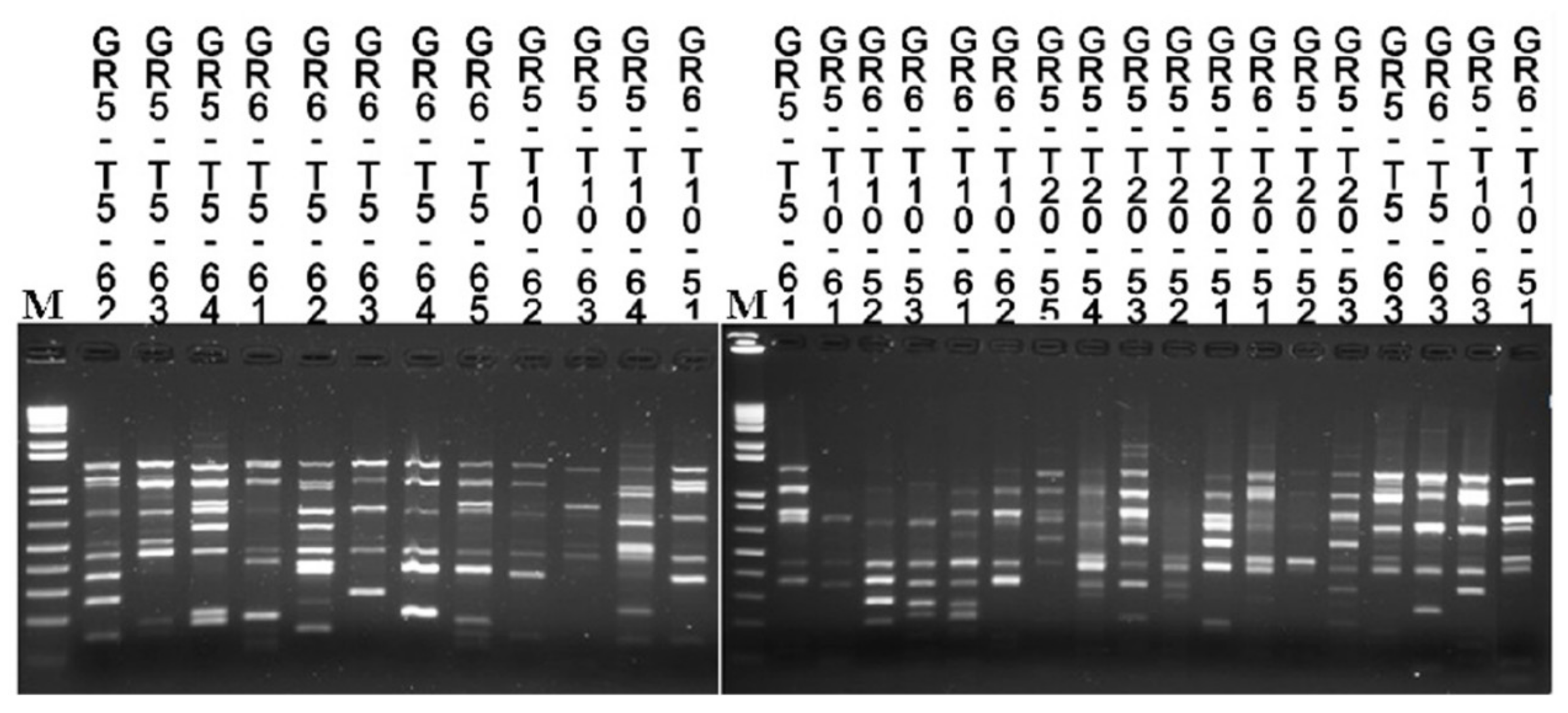
2.3. Microbial Counts, Yeasts Isolation, and Strain Typing
2.4. Technological Characterization of Strains Isolated from Greco Fermentations
2.5. Fermentation Performances of Selected Yeast Strains
2.6. Chemical Determinations
2.7. Statistical Analysis
3. Results
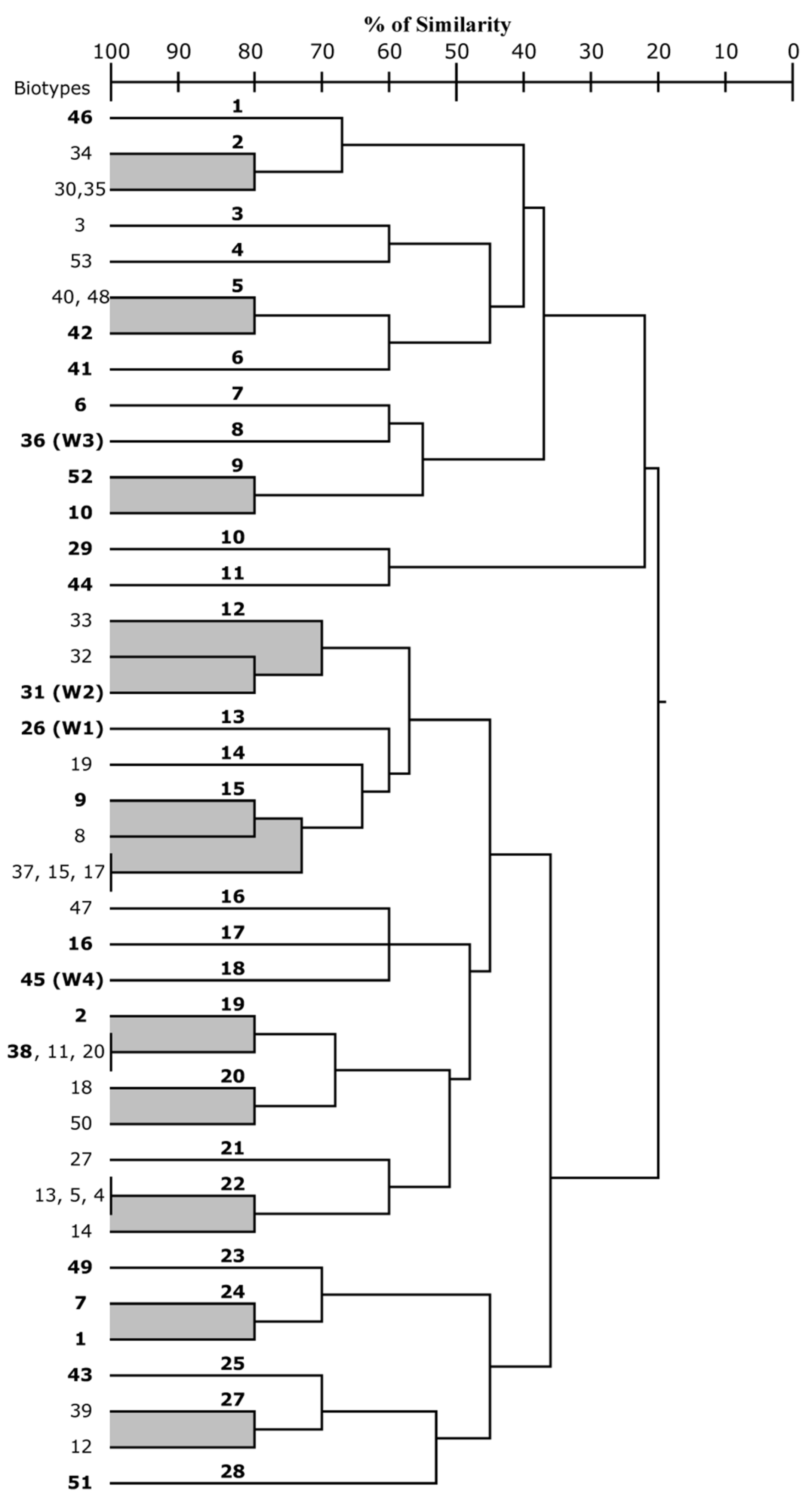
3.1. Commercial Yeast Strains Identification and Genetic Characterization
3.2. Starter Tracking during “Greco di Tufo” and “Aglianico Taurasi” Microvinifications
3.3. Yeast Strains Identification
3.4. Strains Tracking by Molecular Typing
3.5. Technological Characterization of Yeast Cultures from Greco di Tufo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fleet, G.H. Wine yeasts for the future. FEMS Yeast Res. 2008, 8, 979–995. [Google Scholar] [CrossRef] [PubMed]
- Mills, D.A.; Phister, T.; Neeley, E.; Johannsen, E. Wine fermentation. In Molecular Techniques in the Microbial Ecology of Fermented Foods; Cocolin, L., Ercolini, D., Eds.; Springer: New York, NY, USA, 2008; pp. 161–192. [Google Scholar]
- Bokulich, N.A.; Collins, T.S.; Masarweh, C.; Allen, G.; Heymann, H.; Ebeler, S.E.; Mills, D.A. Associations among wine grape microbiome, metabolome, and fermentation behavior suggest microbial contribution to regional wine characteristics. MBio 2016, 7, e00631-16. [Google Scholar] [CrossRef] [PubMed]
- Knight, S.; Klaere, S.; Fedrizzi, B.; Goddard, M.R. Regional microbial signatures positively correlate with differential wine phenotypes: Evidence for a microbial aspect to terroir. Sci. Rep. 2015, 5, 14233. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.E.; Infante, J.J.; Molina, M.; Domínguez, M.; Rebordinos, L.; Cantoral, J.M. Genomic characterization and selection of wine yeast to conduct industrial fermentations of a white wine produced in a SW Spain winery. J. Appl. Microbiol. 2010, 108, 1292–1302. [Google Scholar] [CrossRef] [PubMed]
- Barrajón, N.; Arévalo-Villena, M.; Rodríguez-Aragón, L.J.; Briones, A. Ecological study of wine yeast in inoculated vats from La Mancha Region. Food Control 2009, 20, 778–783. [Google Scholar] [CrossRef]
- Esteve-Zarzoso, B.; Gostincar, A.; Bobet, R.; Uruburu, F.; Querol, A. Selection and molecular characterization of wine yeasts isolated from the “El Penedes” area Spain. Food Microbiol. 2000, 17, 553–562. [Google Scholar] [CrossRef]
- Lopes, C.A.; Rodríguez, M.E.; Sangorrín, M.; Querol, A.; Caballero, A.C. Patagonian wines: Implantation of an indigenous strain of Saccharomyces cerevisiae in fermentations conducted in traditional and modern cellars. J. Ind. Microbiol. Biotechnol. 2007, 34, 139–149. [Google Scholar] [CrossRef]
- Raspor, P.; Cus, F.; Povhe, J.K.; Zagorc, T.; Cadez, N.; Nemanic, J. Yeast population dynamics in spontaneous and inoculated alcoholic fermentations of Zametovka must. Food Technol. Biotechnol. 2002, 40, 95–102. [Google Scholar]
- Liti, G.; Carter, D.M.; Moses, A.M.; Warringer, J.; Parts, L.; James, S.A.; Davey, R.P.; Roberts, I.N.; Burt, A.; Koufopanou, V.; et al. Population genomics of domestic and wild yeasts. Nature 2009, 458, 337–341. [Google Scholar] [CrossRef]
- Schacherer, J.; Shapiro, J.A.; Ruderfer, D.M.; Kruglyak, L. Comprehensive polymorphism survey elucidates population structure of Saccharomyces cerevisiae. Nature 2009, 458, 342–345. [Google Scholar] [CrossRef]
- Cromie, G.A.; Hyma, K.E.; Ludlow, C.L.; Garmendia-Torres, C.; Gilbert, T.L.; May, P.; Huang, A.A.; Dudley, A.M.; Fay, J.C. Genomic sequence diversity and population structure of Saccharomyces cerevisiae assessed by RAD-seq. G3 Genes Genomes Genet. 2013, 3, 2163–2171. [Google Scholar] [CrossRef] [PubMed]
- Martiniuk, J.T.; Pacheco, B.; Russell, G.; Tong, S.; Backstrom, I.; Measday, V. Impact of commercial strain use on Saccharomyces cerevisiae population structure and dynamics in Pinot Noir vineyards and spontaneous fermentations of a canadian winery. PLoS ONE 2016, 1, e0160259. [Google Scholar] [CrossRef]
- Aponte, M.; Blaiotta, G. Selection of an autochthonous Saccharomyces cerevisiae strain for the vinification of “Moscato di Saracena”, a southern Italy (Calabria Region) passito wine. Food Microbiol. 2016, 54, 30–39. [Google Scholar] [CrossRef]
- Cordero-Bueso, G.; Rodríguez, M.E.; Garrido, C.; Cantoral, J.M. Rapid and not culture-dependent assay based on multiplex PCR-SSR analysis for monitoring inoculated yeast strains in industrial wine fermentations. Arch. Microbiol. 2017, 199, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Capece, A.; Pietrafesa, R.; Siesto, G.; Romano, P. Biotechnological approach based on selected Saccharomyces cerevisiae starters for reducing the use of sulfur dioxide in wine. Microorganisms 2020, 8, 738. [Google Scholar] [CrossRef]
- Villano, C.; Carputo, D.; Frusciante, L.; Santoro, X.; Aversano, R. Use of SSR and retrotransposon-based markers to interpret the population structure of native grapevines from southern Italy. Mol. Biotechnol. 2014, 56, 1011–1020. [Google Scholar] [CrossRef]
- Boselli, M.; Scienza, A. Vini e Vitigni Della Campania: Tremila Anni di Storia; Prismi: Naples, Italy, 2003. [Google Scholar]
- Villano, C.; Rinaldi, A.; Lanzillo, C.; Moio, L.; Tamburrino, A.J.; Carputo, D.; Frusciante, L.; Aversano, R. Polyphenol content and differential expression of flavonoid biosynthetic pathway genes in berries of Aglianico. ISHS Ser. Acta Horticult. 2017, 1188, 141–148. [Google Scholar] [CrossRef]
- Rinaldi, A.; Villano, C.; Lanzillo, C.; Tamburrino, A.; Jourdes, M.; Teissedre, P.L.; Moio, L.; Frusciante, L.; Carputo, D.; Aversano, R. Metabolic and RNA profiling elucidates proanthocyanidins accumulation in Aglianico grape. Food Chem. 2017, 233, 52–59. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, F.; La Storia, A.; Blaiotta, G. Monitoring the mycobiota during Greco di Tufo and Aglianico wine fermentation by 18S rRNA gene sequencing. Food Microbiol. 2017, 63, 117–122. [Google Scholar] [CrossRef]
- Sangorrin, M.P.; Zajonskovsky, I.E.; Lopes, C.A.; Rodriguez, M.E.; Giraudo de Van Broock, M.R.; Caballero, A.C. Killer behaviour in wild wine yeasts associated with Merlot and Malbec type musts spontaneously fermented from Northwestern Patagonia (Argentina). J. Basic Microbiol. 2001, 41, 105–113. [Google Scholar] [CrossRef]
- Slifkin, M. Tween 80 opacity test responses of various Candida species. J. Clin. Microbiol. 2000, 38, 4626–4628. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, A.; Blaiotta, G.; Aponte, M.; Moio, L. Effect of yeast strain and some nutritional factors on tannin composition and potential astringency of model wines. Food Microbiol. 2016, 53, 128–134. [Google Scholar] [CrossRef]
- Upholt, W.B. Estimation of DNA sequence divergence from comparison of restriction endonuclease digests. Nucleic Acids Res. 1977, 4, 1257–1265. [Google Scholar] [CrossRef]
- Nei, M.; Li, W. Mathematical model for studying genetic variation in therms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, V.; Ferreira, M.M.; Monteiro, S.; Ferreira, R.B. The microbial community of grape berry. In The Biochemistry of the Grape Berry; Gerós, H., Chaves, M.M., Delrot, S., Eds.; Bentham Science Publishers: Sharjah, UAE, 2012. [Google Scholar]
- Aponte, M.; Blaiotta, G. Potential Role of Yeast Strains Isolated from Grapes in the Production of Taurasi DOCG. Front. Microbiol. 2016, 7, 809. [Google Scholar] [CrossRef]
- Goddard, M.R.; Anfang, N.; Tang, R.; Gardner, R.C.; Jun, C. A distinct population of Saccharomyces cerevisiae in New Zealand: Evidence for local dispersal by insects and human-aided global dispersal in oak barrels. Environ. Microbiol. 2010, 12, 63–73. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Ohta, M.; Richardson, P.M.; Mills, D.A. Monitoring seasonal changes in winery-resident microbiota. PLoS ONE 2013, 8, e66437. [Google Scholar] [CrossRef]
- Beltran, G.; Torija, M.J.; Novo, M.; Ferrer, N.; Poblet, M.; Guillamón, J.M.; Rozès, N.; Mas, A. Analysis of yeast populations during alcoholic fermentation: A six year follow-up study. Syst. Appl. Microbiol. 2002, 25, 287–293. [Google Scholar] [CrossRef]
- Lange, J.N.; Faasse, E.; Tantikachornkiat, M.; Gustafsson, F.S.; Halvorsen, L.C.; Kluftinger, A.; Ledderhof, D.; Durall, D.M. Implantation and persistence of yeast inoculum in Pinot noir fermentations at three Canadian wineries. Int. J. Food Microbiol. 2014, 180, 56–61. [Google Scholar] [CrossRef]
- Delteil, D. Aspectos prácticos del levadurado en condiciones mediterráneas. Técnica de inoculación y relación entre la población seleccionada y la población indígena. Rev. Fr. d’Oenologie 2004, 189, 1–13. [Google Scholar]
- Clavijo, A.; Calderon, I.L.; Paneque, P. Effect of the use of commercial Saccharomyces strains in a newly established winery in Rond (Malaga, Spain). Antonie Van Leeuwenhoek 2011, 99, 727–731. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; Capece, A.; Comitini, F.; Canonico, L.; Siesto, G.; Romano, P. Yeast interactions in inoculated wine fermentation. Front. Microbiol. 2016, 7, 555. [Google Scholar] [CrossRef] [PubMed]
- Capece, A.; Granchi, L.; Guerrini, S.; Mangani, S.; Romaniello, R.; Vincenzini, M.; Romano, P. Diversity of Saccharomyces cerevisiae strains isolated from two Italian wine-producing regions. Front. Microbiol. 2016, 7, 1018. [Google Scholar] [CrossRef] [PubMed]
- Capece, A.; Romaniello, R.; Siesto, G.; Romano, P. Diversity of Saccharomyces cerevisiae yeasts associated to spontaneously fermenting grapes from an Italian “heroic vine-growing area”. Food Microbiol. 2012, 31, 159–166. [Google Scholar] [CrossRef]
- Vigentini, I.; Fabrizio, V.; Faccincani, M.; Picozzi, C.; Comasio, A.; Foschino, R. Dynamics of Saccharomyces cerevisiae populations in controlled and spontaneous fermentations for Franciacorta D.O.C.G. base wine production. Ann. Microbiol. 2014, 64, 639–651. [Google Scholar] [CrossRef]
- Börlin, M.; Miot-Sertier, C.; Vinsonneau, E.; Becquet, S.; Salin, F.; Bely, M.; Lucas, P.; Albertin, W.; Legras, J.-L.; Masneuf-Pomarède, I. The “pied de cuve” as an alternative way to manage indigenous fermentation: Impact on the fermentative process and Saccharomyces cerevisiae diversity. OENO One 2020, 54, 435–442. [Google Scholar] [CrossRef]
- Moschetti, G.; Corona, O.; Gaglio, R.; Squadrito, M.; Parrinello, A.; Settanni, L.; Barone, E.; Francesca, N. Use of fortified pied de cuve as an innovative method to start spontaneous alcoholic fermentation for red winemaking. Aust. J. Grape Wine Res. 2016, 22, 36–45. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fermentations (Starter Strain) | Time (Days) | Microbiological and Oenochemical Parameters | ||||||
|---|---|---|---|---|---|---|---|---|
| Yeast Loads 1 | pH | TA 3 | TS 3 | ET 3 | GLY 3 | AA 3 | ||
| GR1/GR7 (W1) | 0 | 6.78 ± 0.02 a 2 | 3.29 ± 0.01 a | 5.95 ± 0.11 a | 218.51 ± 0.16 a | nd 4 | nd | nd |
| GR2/GR8 (W2) | 6.77 ± 0.01 a | 3.30 ± 0.01 a | 6.03 ± 0.04 a | 218.37 ± 0.06 a | nd | nd | nd | |
| GR3/GR9 (W3) | 6.22 ± 0.23 b | 3.32 ± 0.02 a | 5.92 ± 0.17 a | 218.08 ± 0.57 a | nd | nd | nd | |
| GR4/GR10 (W4) | 6.38 ± 0.04 ab | 3.29 ± 0.01 a | 6.05 ± 0.08 a | 217.80 ± 0.52 a | nd | nd | nd | |
| GR5/GR6 (Control) | 5.24 ± 0.06 c | 3.30 ± 0.01 a | 6.01 ± 0.01 a | 217.60 ± 0.08 a | nd | nd | nd | |
| GR1/GR7 (W1) | 5 | 7.61 ± 0.06 a | 3.25 ± 0.00 a | 7.18 ± 0.01 bc | 141.52 ± 0.59 c | 4.20 ± 0.02 ab | 3.47 ± 0.21 b | 0.13 ± 0.01 b |
| GR2/GR8 (W2) | 7.63 ± 0.03 a | 3.25 ± 0.00 a | 7.10 ± 0.01 cd | 136.58 ± 0.27 d | 4.53 ± 0.08 a | 3.85 ± 0.07 ab | 0.09 ± 0.01 c | |
| GR3/GR9 (W3) | 7.66 ± 0.05 a | 3.25 ± 0.01 a | 7.30 ± 0.01 a | 145.78 ± 0.58 b | 3.97 ± 0.00 b | 3.59 ± 0.08 ab | 0.16 ± 0.01 a | |
| GR4/GR10 (W4) | 7.50 ± 0.06 a | 3.27 ± 0.01 a | 7.07 ± 0.05 d | 154.26 ± 54.2 a | 3.43 ± 0.17 c | 3.62 ± 0.03 ab | 0.18 ± 0.01 a | |
| GR5/GR6 (Control) | 7.61 ± 0.02 a | 3.25 ± 0.00 a | 7.23 ± 0.00 ab | 154.66 ± 0.91 a | 3.39 ± 0.11 c | 3.97 ± 0.13 a | 0.15 ± 0.00 ab | |
| GR1/GR7 (W1) | 10 | 7.56 ± 0.03 ab | 3.28 ± 0.00 b | 7.51 ± 0.43 ab | 27.89 ± 1.90 b | 10.50 ± 0.50 ab | 6.97 ± 0.12 cd | 0.21 ± 0.04 ab |
| GR2/GR8 (W2) | 7.58 ± 0.00 ab | 3.32 ± 0.01 a | 7.32 ± 0.06 b | 19.92 ± 2.95 b | 11.17 ± 0.33 a | 7.39 ± 0.05 bc | 0.14 ± 0.06 | |
| GR3/GR9 (W3) | 7.64 ± 0.05 a | 3.27 ± 0.00 b | 8.24 ± 0.01 a | 40.85 ± 4.87 a | 9.51 ± 0.24 b | 6.80 ± 0.01 d | 0.19 ± 0.01 ab | |
| GR4/GR10 (W4) | 7.50 ± 0.01 b | 3.32 ± 0.01 a | 7.69 ± 0.01 ab | 28.39 ± 0.57 b | 10.27 ± 0.07 ab | 7.52 ± 0.20 | 0.29 ± 0.00 a | |
| GR5/GR6 (Control) | 7.58 ± 0.04 ab | 3.30 ± 0.01 ab | 8.14 ± 0.05 a | 27.24 ± 0.95 b | 10.25 ± 0.01 ab | 8.21 ± 0.01 a | 0.22 ± 0.00 ab | |
| GR1/GR7 (W1) | 21 | 6.32 ± 0.09 b | 3.30 ± 0.00 bc | 7.55 ± 0.09 b | 4.84 ± 0.12 ab | 12.68 ± 0.06 abc | 7.39 ± 0.15 c | 0.19 ± 0.01 c |
| GR2/GR8 (W2) | 6.07 ± 0.08 b | 3.32 ± 0.01 ab | 7.50 ± 0.01 b | 3.10 ± 0.88 b | 12.84 ± 0.07 a | 7.59 ± 0.06 bc | 0.10 ± 0.00 d | |
| GR3/GR9 (W3) | 6.77 ± 0.04 a | 3.29 ± 0.01 c | 8.01 ± 0.08 a | 7.68 ± 1.80 a | 12.45 ± 0.01 c | 7.44 ± 0.06 c | 0.20 ± 0.01 bc | |
| GR4/GR10 (W4) | 6.17 ± 0.03 b | 3.33 ± 0.00 a | 7.33 ± 0.07 b | 5.30 ± 0.95 ab | 12.58 ± 0.08 bc | 8.00 ± 0.15 b | 0.31 ± 0.01 a | |
| GR5/GR6 (Control) | 6.14 ± 0.09 b | 3.32 ± 0.00 a | 7.83 ± 0.01 a | 3.24 ± 0.62 b | 12.77 ± 0.04 ab | 8.62 ± 0.04 a | 0.22 ± 0.01 b | |
| Fermentations (Starter Strain) | Time (Days) | Microbiological and Oenochemical Parameters | ||||||
|---|---|---|---|---|---|---|---|---|
| Yeast loads 1 | pH | TA 3 | TS 3 | ET 3 | GLY 3 | AA 3 | ||
| AGL1/AGL7 (R1) | 0.5 | 7.2 ± 0.26 a 2 | 3.35 ± 0.04 a | 4.91 ± 0.16 a | 209.00 ± 1.41 a | nd 4 | nd | nd |
| AGL2/AGL8 (R2) | 6.57 ± 0.55 ab | 3.38 ± 0.02 a | 5.29 ± 0.05 a | 214.00 ± 2.83 a | nd | nd | nd | |
| AGL3/AGL9 (R3) | 7.20 ± 0.34 a | 3.38 ± 0.00 a | 5.06 ± 0.16 a | 217.50 ± 2.12 a | nd | nd | nd | |
| AGL4/AGL10 (R4) | 7.37 ± 0.15 a | 3.38 ± 0.00 a | 5.10 ± 0.10 a | 212.00 ± 2.83 a | nd | nd | nd | |
| AGL5/AGL6 (Control) | 5.36 ± 0.69 b | 3.33 ± 0.01 a | 4.99 ± 0.16 a | 217.00 ± 4.24 a | nd | nd | nd | |
| AGL1/AGL7 (R1) | 6.5 | 7.31 ± 0.04 a | 3.42 ± 0.04 a | 7.59 ± 0.18 abc | 18.29 ± 2.81 b | 11.42 ± 0.02 a | 7.29 ± 0.23 ab | 0.28 ± 0.02 a |
| AGL2/AGL8 (R2) | 6.93 ± 0.19 a | 3.44 ± 0.01 a | 7.03 ± 0.06 c | 26.14 ± 2.04 b | 11.23 ± 0.04 a | 6.96 ± 0.03 b | 0.21 ± 0.01 b | |
| AGL3/AGL9 (R3) | 7.18 ± 0.23 a | 3.44 ± 0.02 a | 7.65 ± 0.13 ab | 23.41 ± 12.91 b | 11.28 ± 0.65 a | 7.65 ± 0.49 ab | 0.29 ± 0.00 a | |
| AGL4/AGL10 (R4) | 7.10 ± 0.14 a | 3.45 ± 0.01 a | 7.72 ± 0.21 a | 18.78 ± 6.77 b | 11.40 ± 0.39 a | 8.60 ± 0.09 a | 0.20 ± 0.00 b | |
| AGL5/AGL6 (Control) | 7.28 ± 0.14 a | 3.38 ± 0.01 a | 7.06 ± 0.13 bc | 113.20 ± 10.82 a | 5.90 ± 0.47 b | 4.82 ± 0.64 c | 0.14 ± 0.01 c | |
| AGL1/AGL7 (R1) | 12.5 | 6.75 ± 0.34 a | 3.49 ± 0.03 a | 7.16 ± 0.17 abc | 2.03 ± 0.08 a | 11.82 ± 0.01 b | 8.83 ± 0.03 d | 0.30 ± 0.01 ab |
| AGL2/AGL8 (R2) | 6.28 ± 0.18 a | 3.40 ± 0.22 a | 6.73 ± 0.04 c | 1.82 ± 0.11 a | 12.46 ± 0.20 a | 9.16 ± 0.02 cd | 0.27 ± 0.01 bc | |
| AGL3/AGL9 (R3) | 6.73 ± 0.20 a | 3.53 ± 0.01 a | 7.13 ± 0.09 bc | 1.74 ± 0.06 a | 12.29 ± 0.08 ab | 9.50 ± 0.13 bc | 0.35 ± 0.01 a | |
| AGL4/AGL10 (R4) | 6.44 ± 0.06 a | 3.54 ± 0.05 a | 7.40 ± 0.27 ab | 1.81 ± 0.21 a | 12.20 ± 0.04 ab | 10.62 ± 0.21 a | 0.24 ± 0.02 c | |
| AGL5/AGL6 (Control) | 7.02 ± 0.09 a | 3.47 ± 0.01 a | 7.74 ± 0.04 a | 2.25 ± 0.17 a | 12.06 ± 0.26 ab | 9.90 ± 0.11 b | 0.25 ± 0.01 c | |
| Must | Fermentation (Starter Strain) | Days of Fermentation | Total N. of Isolates | N. of S. cerevisiae 1 | N. of Non-Saccharomyces Strains (Species) 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 5 | 10 | 20 | ||||||||
| Greco di Tufo | GR1 (W1) | 7 (1) 2 | 4 | 5 | 5 | 21 | 20 | 1 | (M. pulcherrima) | ||
| GR7 (W1) | 4 | 4 | 5 | 4 | 17 | 17 | - | ||||
| GR2 (W2) | 4 (1) | 4 | 4 | 5 | 17 | 16 | 1 | (M. pulcherrima) | |||
| GR8 (W2) | 3 | 5 | 5 | 3 | 16 | 16 | - | ||||
| GR3 (W3) | 5 (2) | 4 | 5 | 6 | 20 | 18 | 2 | (P. occidentalis; H. uvarum) | |||
| GR9 (W3) | 5 (1) | 4 | 4 | 3 | 16 | 15 | 1 | (M. pulcherrima) | |||
| GR4 (W4) | 6 (1) | 4 | 5 | 4 | 19 | 18 | 1 | (P. anomala) | |||
| GR10 (W4) | 3 (1) | 5 | 5 | 5 | 18 | 17 | 1 | (P. fermentans) | |||
| GR5 (Contr.) | 6 (6) | 4 | 4 | 5 | 19 | 13 | 6 | (P. fermentans; H. uvarum) | |||
| GR6 (Contr.) | 6 (6) | 5 | 5 | 3 | 19 | 13 | 6 | (P. fermentans) | |||
| 182 | 163 | 19 | |||||||||
| Must | Fermentation (Starter Strain) | Days of Fermentation | Total N. of Isolates | N. of S. cerevisiae1 | N. of Non-Saccharomyces Strains (Species) | ||||||
| 0.5 | 6.5 | 12.5 | |||||||||
| Aglianico | AGL1 (R1) | 5 (1) | 3 | 3 | 11 | 10 | 1 | (St.la bacillaris) | |||
| AGL7 (R1) | 3 (3) | 3 (1) | 3 | 9 | 5 | 4 | ( P. fermentans, St.la bacillaris) | ||||
| AGL2 (R2) | 6 (3) | 4 | 3 | 13 | 10 | 3 | (M. pulcherrima; St.la bacillaris) | ||||
| AGL8 (R2) | 4 (4) | 3 | 5 | 12 | 8 | 4 | ( H. uvarum) | ||||
| AGL3 (R3) | 4 | 4 | 3 | 11 | 11 | - | |||||
| AGL9 (R3) | 5 (4) | 4 | 4 | 13 | 9 | 4 | ( H. uvarum) | ||||
| AGL4 (R4) | 5 (1) | 3 | 4 | 12 | 11 | 1 | (P. fermentans) | ||||
| AGL10 (R4) | 6 (1) | 4 | 4 | 14 | 13 | 1 | (H. uvarum) | ||||
| AGL5 (Contr.) | 3 (2) | 5 (1) | 4 | 12 | 9 | 3 | (P. fermentans; H. uvarum; St.la bacillaris) | ||||
| AGL6 (Contr.) | 3 (3) | 4 | 5 | 12 | 9 | 3 | (St.la bacillaris; H. uvarum) | ||||
| 119 | 95 | 24 | |||||||||
| Must | Fermentation (Starter Strain) | Days of Fermentation | |||||
|---|---|---|---|---|---|---|---|
| 0 | 5 | 10 | 20 | ||||
| GR | GR1 (W1) | 6 * (6a) ** | 4 (4a) | 5(5a) | 5 (3a, 1b, 1c) | ||
| GR7 (W1) | 4 (4a) | 4 (3a, 1d) | 5 (3a, 2e) | 4 (4a) | |||
| W1 % of dominance | 100.0 | 87.5 | 80.0 | 77.8 | |||
| GR2 (W2) | 4 (3a) | 4 (3a, 1b) | 4 (4a) | 5 (4a, 1c) | |||
| GR8 (W2) | 3 (2a, 1d) | 5 (5a) | 5 (5a) | 3 (2a, 1e) | |||
| W2 % of dominance | 83.3 | 88.9 | 100.0 | 75.0 | |||
| GR3 (W3) | 3 (2a, 1b) | 4 (3a, 1c) | 5 (3a, 1d, 1e) | 6 (4a, 1e; 1f) | |||
| GR9 (W3) | 4 (4a) | 4 (2a, 1g, 1h) | 4 (3a, 1i) | 3 (3a) | |||
| W1 % of dominance | 85.7 | 62.5 | 66.7 | 77.8 | |||
| GR4 (W4) | 5 (5a) | 4 (2a, 2b) | 5 (2a, 2c, 1d) | 4 (3a, 1e) | |||
| GR10 (W4) | 2 (2a) | 5 (4a, 1f) | 6 (3a, 1f, 1g) | 5 (3a, 1h, 1i) | |||
| W4 % of dominance | 100.0 | 66.7 | 45.5 | 66.7 | |||
| GR5 (Contr.) | - | 4 (a–d) | 4 (e–h) | 5 (i–m) | |||
| GR6 (Contr.) | - | 5 (n–r) | 5 (a, s–v) | 3 (w–y) | |||
| Must | Fermentation (Starter Strain) | Days of Fermentation | |||||
| 0.5 | 6.5 | 12.5 | |||||
| AGL | AGL1 (R1) | 4 * (4a) ** | 3 (2a; 1b) | 3 (2a; 1c) | |||
| AGL7 (R1) | - | 2 (2a) | 2 (2a) | ||||
| R1 % of dominance | 50.0 | 80.0 | 80.0 | ||||
| AGL2 (R2) | 3 (3a) | 4 (3a; 1b) | 2 (2a) | ||||
| AGL8 (R2) | - | 3 (3a) | 5 (3a; 1c; 1d) | ||||
| R2 % of dominance | 50.0 | 85.7 | 71.4 | ||||
| AGL3 (R3) | 4 (4a) | 4 (3a; 1b) | 3 (3a) | ||||
| AGL9 (R3) | 1 (1a) | 4 (4a) | 4 (4a) | ||||
| R3 % of dominance | 100.0 | 87.5 | 100.0 | ||||
| AGL4 (R4) | 4 (4a) | 3 (1a; 2b) | 4 (2a; 1b; 1c) | ||||
| AGL10 (R4) | 5 (4a, 1b) | 4 (3a, 1d) | 4 (2d; 2e) | ||||
| R4 % of dominance | 88.9 | 57.1 | 25.0 | ||||
| AGL5 (Contr.) | - | 4 (2a; 1b; 1c) | 4 (1d; 1e; 1f; 1g) | ||||
| AGL6 (Contr.) | - | 4 (1h; 1i; 1l; 1m) | 5 (1h; 1i; 1n; 1o; 1p) | ||||
| ID | a Strain | b Ethanol Tolerance (%) | c H2S Production | d Antagonistic Activity | e Growth in SM at 14 °C | |
|---|---|---|---|---|---|---|
| °Brix Reduction | Type of Growth | |||||
| 1 | GR5-T5-61 | 15 | 2.5 | + | 3.85 | S |
| 2 | GR5-T5-62 | 15 | 1.5 | − | 3.25 | S |
| 3 | GR5-T5-63 | 14 | 2.0 | − | 2.45 | D |
| 4 | GR6-T5-61 | 14 | 2.0 | − | 2.85 | S |
| 5 | GR6-T5-63 | 14 | 2.0 | − | 2.55 | S |
| 6 | GR6-T5-64 | 15 | 3.5 | ++ | 2.45 | D |
| 7 | GR6-T5-65 | 15 | 2.5 | + | 2.55 | S |
| 8 | GR5-T10-61 | 14 | 2.5 | − | 2.95 | S |
| 9 | GR5-T10-63 | 14 | 2.5 | − | 4.05 | S |
| 10 | GR5-T10-64 | 15 | 3.0 | ++ | 3.05 | D |
| 11 | GR6-T10-51 | 14 | 1.0 | − | 3.35 | S |
| 12 | GR6-T10-52 | 15 | 3.5 | ++ | 2.65 | S |
| 13 | GR6-T10-53 | 14 | 2.0 | − | 2.55 | S |
| 14 | GR6-T10-61 | 14 | 2.0 | + | 2.85 | S |
| 15 | GR5-T20-51 | 14 | 2.5 | − | 2.40 | S |
| 16 | GR5-T20-52 | 15 | 2.0 | − | 4.15 | S |
| 17 | GR5-T20-53 | 14 | 2.5 | − | 2.35 | S |
| 18 | GR6-T20-51 | 14 | 1.5 | − | 2.75 | S |
| 19 | GR6-T20-53 | 14 | 2.5 | ± | 2.85 | S |
| 20 | GR5-T10-62 | 14 | 1.5 | − | 3.25 | S |
| 26 | W1 | 14 | 2.5 | + | 3.35 | S |
| 27 | GR1-T20-53 | 14 | 4.5 | ++ | 2.85 | S |
| 29 | GR7-T5-64 | 13 | 4.0 | ++ | 4.35 | D |
| 30 | GR7-T10-61 | 14 | 3.0 | + | 2.55 | D |
| 31 | W2 | 13 | 2.5 | + | 2.35 | S |
| 32 | GR2-T5-61 | 13 | 2.5 | − | 2.35 | S |
| 33 | GR2-T20-55 | 13 | 2.5 | − | 3.15 | S |
| 34 | GR8-T0-51 | 13 | 3.0 | + | 2.85 | D |
| 35 | GR8-T20-42 | 14 | 3.0 | + | 2.75 | D |
| 36 | W3 | 15 | 2.5 | ++ | 2.85 | D |
| 37 | GR3-T0-54 | 14 | 2.5 | − | 2.45 | S |
| 38 | GR3-T5-62 | 14 | 1.5 | − | 3.35 | S |
| 39 | GR3-T10-61 | 15 | 1.5 | ++ | 2.95 | S |
| 40 | GR3-T10-64 | 15 | 3.0 | − | 2.85 | D |
| 41 | GR3-T20-55 | 15 | 5.0 | − | 3.55 | D |
| 42 | GR9-T5-63 | 15 | 3.0 | − | 3.35 | D |
| 43 | GR9-T5-64 | 15 | 1.5 | ++ | 3.15 | S |
| 44 | GR9-T10-63 | 12 | 4.0 | − | 4.35 | S |
| 45 | W4 | 15 | 5.0 | − | 3.35 | S |
| 46 | GR4-T5-61 | 13 | 3.0 | + | 3.15 | D |
| 47 | GR4-T10-63 | 15 | 4.5 | − | 2.85 | S |
| 48 | GR4-T10-65 | 15 | 3.0 | − | 2.85 | D |
| 49 | GR4-T20-53 | 15 | 3.5 | + | 2.75 | S |
| 50 | GR10-T5-61 | 13 | 1.5 | − | 2.75 | S |
| 51 | GR10-T10-65 | 14 | 3.5 | ++ | 3.35 | S |
| 52 | GR10-T20-43 | 15 | 3.5 | ++ | 2.95 | D |
| 53 | GR10-T20-44 | 13 | 2.0 | − | 2.95 | D |
| ID | Strains | FV 1 | FP 2 | Content of | pH | TA 4 | Wine Color e | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| AA 3 | RS 3 | GLY 3 | ET 3 | I | H | ||||||
| 46 | GR4-T5-61 | 0.95 ± 0.05 defg 5 | 7.63 ± 0.04 abc | 0.15 ± 0.00 e | 1.75 ± 1.07 ef | 5.16 ± 0.41 bcdef | 11.13 ± 1.03 a | 3.08 ± 0.01 a | 9.77 ± 0.02 c | 1.03 ± 0.04 bcd | 3.94 ± 0.07 ab |
| 42 | GR9-T5-63 | 1.47 ± 0.08 ab | 7.60 ± 0.20 abc | 0.36 ± 0.01 abcd | 6.41 ± 2.19 bcde | 4.81 ± 0.18 cdefgh | 11.47 ± 1.11 a | 3.09 ± 0.04 a | 9.90 ± 0.21 bc | 1.07 ± 0.07 bcd | 3.83 ± 0.23 ab |
| 6 | GR6-T5-64 | 1.16 ± 0.05 bcdef | 7.44 ± 0.02 abc | 0.22 ± 0.04 de | 6.47 ± 1.20 bcde | 4.65 ± 0.30 efgh | 11.12 ± 0.99 a | 3.05 ± 0.05 a | 11.53 ± 0.66 ab | 1.19 ± 0.06 abc | 3.49 ± 0.21 bcd |
| 29 | GR7-T5-64 | 1.29 ± 0.06 abcd | 7.65 ± 0.06 abc | 0.36 ± 0.01 abcd | 3.14 ± 0.62 def | 4.56 ± 0.19 efgh | 11.72 ± 0.65 a | 3.07 ± 0.01 a | 9.85 ± 0.07 c | 1.31 ± 0.15 ab | 3.07 ± 0.07 cd |
| 38 | GR3-T5-62 | 1.20 ± 0.11 bcdef | 7.63 ± 0.05 abc | 0.49 ± 0.04 a | 2.34 ± 1.19 ef | 5.36 ± 0.20 bcd | 11.44 ± 0.23 a | 3.13 ± 0.00 a | 9.43 ± 0.18 c | 1.06 ± 0.02 bcd | 3.86 ± 0.12 ab |
| 2 | GR5-T5-62 | 0.88 ± 0.01 fg | 7.61 ± 0.03 abc | 0.34 ± 0.00 abcd | 2.60 ± 1.55 ef | 6.60 ± 0.05 a | 11.43 ± 0.22 a | 3.08 ± 0.01 a | 10.74 ± 0.18 abc | 1.12 ± 0.05 abcd | 3.75 ± 0.12 abc |
| 7 | GR6-T5-65 | 1.35 ± 0.10 abc | 7.46 ± 0.04 abc | 0.33 ± 0.00 bcd | 8.33 ± 3.15 abcd | 4.46 ± 0.13 gh | 11.99 ± 0.09 a | 3.06 ± 0.01 a | 10.21 ± 0.12 bc | 1.08 ± 0.05 bcd | 3.85 ± 0.12 ab |
| 1 | GR5-T5-61 | 1.24 ± 0.08 bcde | 7.44 ± 0.04 abc | 0.36 ± 0.02 abcd | 9.44 ± 3.04 abc | 4.78 ± 0.14 defgh | 11.44 ± 0.42 a | 3.04 ± 0.04 a | 10.76 ± 0.06 abc | 1.03 ± 0.00 bcd | 4.01 ± 0.07 ab |
| 43 | GR9-T5-64 | 0.96 ± 0.03 defg | 7.65 ± 0.04 abc | 0.33 ± 0.01 bcd | 4.57 ± 1.32 cdef | 4.37 ± 0.06 h | 11.67 ± 0.69 a | 3.09 ± 0.01 a | 10.16 ± 0.11 bc | 1.09 ± 0.01 bcd | 3.89 ± 0.08 ab |
| 10 | GR5-T10-64 | 1.39 ± 0.07 ab | 7.65 ± 0.22 abc | 0.22 ± 0.10 de | 0.50 ± 0.00 f | 4.50 ± 0.05 fgh | 11.17 ± 2.08 a | 3.05 ± 0.01 a | 9.65 ± 0.07 c | 1.14 ± 0.06 abcd | 3.61 ± 0.17 bc |
| 44 | GR9-T10-63 | 0.77 ± 0.13 g | 7.80 ± 0.04 ab | 0.42 ± 0.00 ab | 2.94 ± 1.34 def | 5.49 ± 0.10 ab | 11.82 ± 0.58 a | 3.07 ± 0.04 a | 10.11 ± 0.35 bc | 1.22 ± 0.15 abc | 2.80 ± 0.23 d |
| 9 | GR5-T10-63 | 1.60 ± 0.00 a | 7.71 ± 0.13 ab | 0.36 ± 0.05 abcd | 0.50 ± 0.00 f | 5.24 ± 0.18 bcde | 12.14 ± 1.08 a | 3.07 ± 0.03 a | 10.09 ± 0.05 bc | 0.87 ± 0.02 d | 4.36 ± 0.06 a |
| 51 | GR10-T10-65 | 0.95 ± 0.02 defg | 7.38 ± 0.04 abc | 0.36 ± 0.01 abcd | 10.75 ± 0.64 ab | 4.94 ± 0.03 bcdefgh | 11.58 ± 0.58 a | 3.04 ± 0.03 a | 10.84 ± 0.47 abc | 1.04 ± 0.06 bcd | 3.95 ± 0.2 ab |
| 41 | GR3-T20-55 | 1.41 ± 0.05 ab | 7.76 ± 0.08 ab | 0.39 ± 0.01 ab | 0.50 ± 0.00 f | 6.26 ± 0.19 a | 11.56 ± 0.56 a | 3.11 ± 0.01 a | 10.14 ± 0.03 bc | 1.03 ± 0.09 bcd | 3.54 ± 0.48 bc |
| 52 | GR10-T20-43 | 1.01 ± 0.13 cdefg | 7.65 ± 0.07 abc | 0.22 ± 0.11 cde | 0.50 ± 0.00 f | 4.69 ± 0.00 defgh | 11.65 ± 0.63 a | 3.05 ± 0.01 a | 9.69 ± 0.03 c | 1.04 ± 0.01 bcd | 3.71 ± 0.26 abc |
| 16 | GR5-T20-52 | 1.42 ± 0.00 ab | 7.80 ± 0.12 ab | 0.37 ± 0.05 abcd | 0.50 ± 0.00 f | 5.52 ± 0.01 b | 11.20 ± 0.50 a | 3.05 ± 0.01 a | 10.87 ± 0.00 abc | 1.07 ± 0.03 bcd | 3.84 ± 0.12 ab |
| 49 | GR4-T20-53 | 1.31 ± 0.01 abc | 7.08 ± 0.04 c | 0.38 ± 0.01 abc | 12.78 ± 0.35 a | 4.83 ± 0.06 bcdefgh | 11.23 ± 0.58 a | 3.14 ± 0.13 a | 12.03 ± 1.52 a | 1.05 ± 0.08 bcd | 3.76 ± 0.11 abc |
| 26 | W1 | 0.80 ± 0.06 g | 7.21 ± 0.08 bc | 0.34 ± 0.02 abcd | 10.85 ± 0.55 ab | 5.10 ± 0.21 bcdefg | 11.24 ± 0.31 a | 3.07 ± 0.03 a | 10.37 ± 0.29 abc | 1.04 ± 0.04 bcd | 3.89 ± 0.10 ab |
| 31 | W2 | 0.93 ± 0.08 efg | 7.67 ± 0.03 ab | 0.15 ± 0.00 e | 0.50 ± 0.00 f | 6.87 ± 0.24 a | 11.55 ± 0.70 a | 3.07 ± 0.01 a | 9.79 ± 0.11 c | 1.11 ± 0.15 bcd | 3.82 ± 0.15 ab |
| 36 | W3 | 1.05 ± 0.16 cdefg | 7.84 ± 0.20 a | 0.32 ± 0.00 bcd | 1.72 ± 1.02 ef | 4.52 ± 0.08 fgh | 11.43 ± 1.25 a | 3.06 ± 0.01 a | 10.12 ± 0.00 bc | 1.40 ± 0.06 a | 3.91 ± 0.03 ab |
| 45 | W4 | 0.95 ± 0.15 defg | 7.60 ± 0.22 abc | 0.31 ± 0.03 bcde | 3.67 ± 1.79 def | 5.02 ± 0.15 bcdefgh | 11.88 ± 0.76 a | 3.05 ± 0.00 a | 10.22 ± 0.45 bc | 1.01 ± 0.02 cd | 3.96 ± 0.10 ab |
| Average values | Total | 1.15 ± 0.24 | 7.58 ± 0.19 | 0.32 ± 0.09 | 4.32 ± 3.99 | 5.13 ± 0.70 | 11.52 ± 0.29 | 3.07 ± 0.03 | 10.30 ± 0.64 | 1.10 ± 0.11 | 3.75 ± 0.33 |
| Wild | 1.20 ± 0.24 | 7.58 ± 0.18 | 0.33 ± 0.08 | 4.35 ± 3.98 | 5.07 ± 0.63 | 11.52 ± 0.30 | 3.07 ± 0.03 | 10.34 ± 0.70 | 1.08 ± 0.10 | 3.72 ± 0.36 | |
| Commercial | 0.93 ± 0.10 | 7.58 ± 0.27 | 0.28 ± 0.09 | 4.19 ± 4.63 | 5.38 ± 1.03 | 11.53 ± 0.27 | 3.06 ± 0.01 | 10,13 ± 0.25 | 1.14 ± 0.18 | 3.90 ± 0.06 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aponte, M.; Romano, R.; Villano, C.; Blaiotta, G. Dominance of S. cerevisiae Commercial Starter Strains during Greco di Tufo and Aglianico Wine Fermentations and Evaluation of Oenological Performances of Some Indigenous/Residential Strains. Foods 2020, 9, 1549. https://doi.org/10.3390/foods9111549
Aponte M, Romano R, Villano C, Blaiotta G. Dominance of S. cerevisiae Commercial Starter Strains during Greco di Tufo and Aglianico Wine Fermentations and Evaluation of Oenological Performances of Some Indigenous/Residential Strains. Foods. 2020; 9(11):1549. https://doi.org/10.3390/foods9111549
Chicago/Turabian StyleAponte, Maria, Raffaele Romano, Clizia Villano, and Giuseppe Blaiotta. 2020. "Dominance of S. cerevisiae Commercial Starter Strains during Greco di Tufo and Aglianico Wine Fermentations and Evaluation of Oenological Performances of Some Indigenous/Residential Strains" Foods 9, no. 11: 1549. https://doi.org/10.3390/foods9111549
APA StyleAponte, M., Romano, R., Villano, C., & Blaiotta, G. (2020). Dominance of S. cerevisiae Commercial Starter Strains during Greco di Tufo and Aglianico Wine Fermentations and Evaluation of Oenological Performances of Some Indigenous/Residential Strains. Foods, 9(11), 1549. https://doi.org/10.3390/foods9111549