Potential Application of Apilactobacillus kunkeei for Human Use: Evaluation of Probiotic and Functional Properties

 ,
, 
_Di_Marco.png) ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Biochemical Characterization
2.3. Antibiotic Susceptibility
2.4. Antimicrobial Activity
2.5. Osmotic Tolerance Assay in Sugar Syrup
2.6. Simulated Gastrointestinal Conditions
2.7. Screening for Cholesterol Lowering
2.8. Statistical Analysis
3. Results
3.1. Biochemical Characterization
3.2. Antibiotic Susceptibility and Antimicrobial Activity
3.3. Bacterial Viability in Sugar Syrup
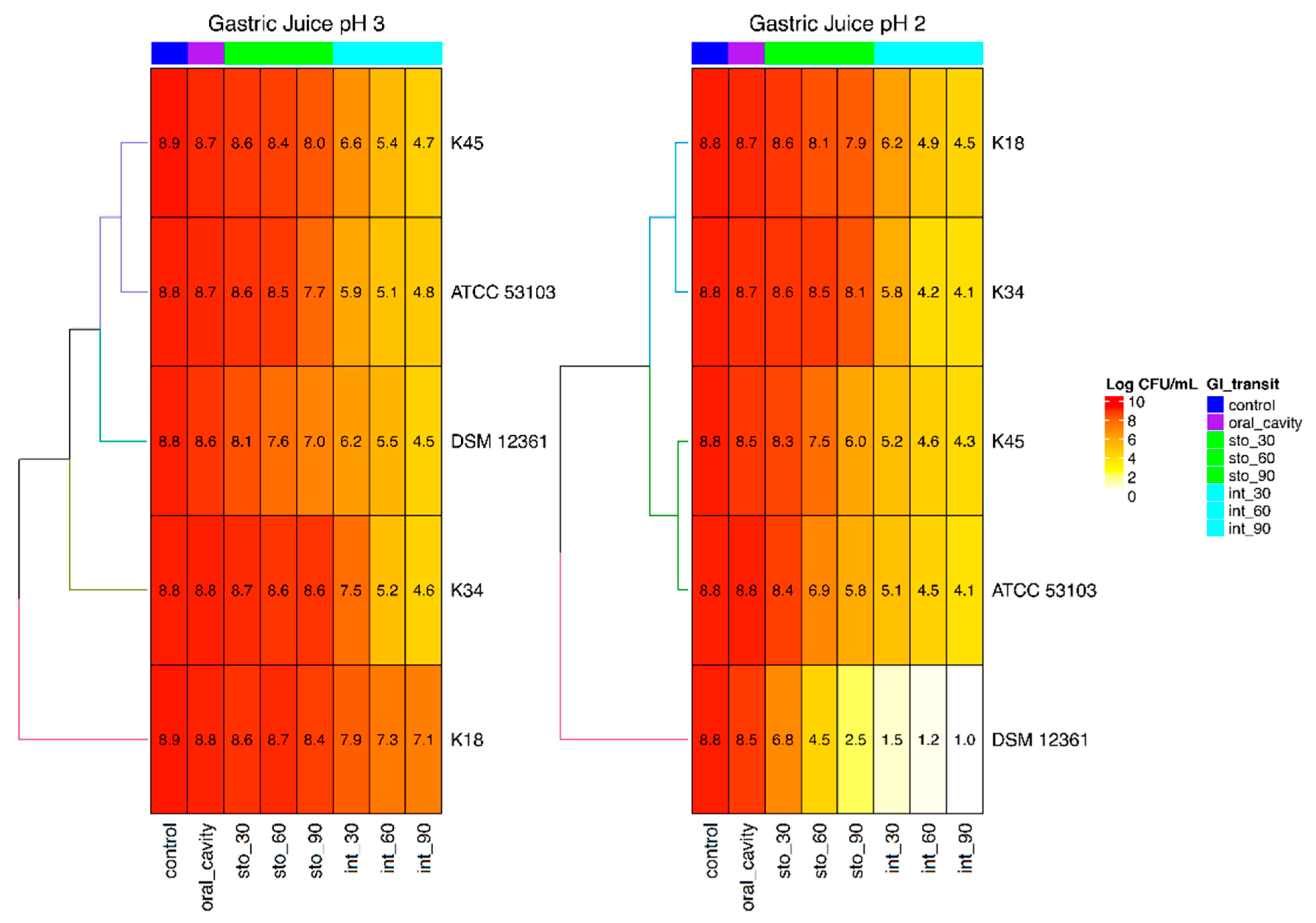
3.4. Viability during Simulated Gastrointestinal Transit
3.5. Cholesterol Lowering Capacity
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vasiljevic, T.; Shah, N.P. Probiotics—From Metchnikoff to bioactives. Int. Dairy J. 2008, 18, 714–728. [Google Scholar] [CrossRef]
- Sornplang, P.; Piyadeatsoontorn, S. Probiotic isolates from unconventional sources: A review. J. Anim. Sci. Technol. 2016, 58, 26. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; Filannino, P.; Gobbetti, M. Novel fermented fruit and vegetable-based products. In Novel Food Fermentation Technologies; Springer: Cham, Switzerland, 2016; pp. 279–291. [Google Scholar] [CrossRef]
- Vitali, B.; Minervini, G.; Rizzello, C.G.; Spisni, E.; Maccaferri, S.; Brigidi, P.; Gobbettii, M.; Di Cagno, R. Novel probiotic candidates for humans isolated from raw fruits and vegetables. J. Food Microbiol. 2012, 31, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Pennacchia, C.; Vaughan, E.E.; Villani, F. Potential probiotic Lactobacillus strains from fermented sausages: Further investigations on their probiotic properties. Meat Sci. 2006, 73, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.H.; Shim, Y.Y.; Cha, S.-K.; Chee, K.M. Probiotic characteristics of lactic acid bacteria isolated from kimchi. J. Appl. Microbiol. 2010, 109, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Honey Chandran, C.; Keerthi, T.R. Probiotic potency of Lactobacillus plantarum KX519413 and KX519414 isolated from honey bee gut. FEMS Microbiol. Lett. 2018, 365, fnx285. [Google Scholar]
- Sakandar, H.A.; Kubow, S.; Sadiq, F.A. Isolation and in-vitro probiotic characterization of fructophilic lactic acid bacteria from Chinese fruits and flowers. LWT 2019, 104, 70–75. [Google Scholar] [CrossRef]
- Nagpal, R.; Kumar, A.; Kumar, M. Fortification and fermentation of fruit juices with probiotic lactobacilli. Ann. Microbiol. 2012, 62, 1573–1578. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Calder, P.C. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. 2014, 11, 506. [Google Scholar] [CrossRef]
- Brodmann, T.; Endo, A.; Gueimonde, M.; Vinderola, G.; Kneifel, W.; de Vos, W.M.; Gómez-Gallego, C. Safety of novel microbes for human consumption: Practical examples of assessment in the European Union. Front. Microbiol. 2017, 8, 1725. [Google Scholar] [CrossRef]
- Endo, A.; Salminen, S. Honeybees and beehives are rich sources for fructophilic lactic acid bacteria. Syst. Appl. Microbiol. 2013, 36, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Kwong, W.K.; Moran, N.A. Gut microbial communities of social bees. Nat. Rev. Microbiol. 2016, 14, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Vásquez, A.; Forsgren, E.; Fries, I.; Paxton, R.J.; Flaberg, E.; Szekely, L.; Olofsson, T.C. Symbionts as major modulators of insect health: Lactic acid bacteria and honeybees. PLoS ONE 2012, 7, 33188. [Google Scholar] [CrossRef]
- Endo, A.; Maeno, S.; Tanizawa, Y.; Kneifel, W.; Arita, M.; Dicks, L.; Salminen, S. Fructophilic lactic acid bacteria, a unique group of fructose-fermenting microbes. Appl. Environ. Microbiol. 2018, 84, 01290-18. [Google Scholar] [CrossRef]
- Filannino, P.; Di Cagno, R.; Tlais, A.Z.A.; Cantatore, V.; Gobbetti, M. Fructose-rich niches traced the evolution of lactic acid bacteria toward fructophilic species. Crit. Rev. Microbiol. 2019, 45, 65–81. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.M.; Mattarelli, P.; Watanabe, K. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Neveling, D.P.; Endo, A.; Dicks, L.M. Fructophilic Lactobacillus kunkeei and Lactobacillus brevis isolated from fresh flowers, bees and beehives. Curr. Microbiol. 2012, 65, 507–515. [Google Scholar] [CrossRef]
- Butler, E.; Oien, R.F.; Lindholm, C.; Olofsson, T.C.; Nilson, B.; Vasquez, A. A pilot study investigating lactic acid bacterial symbionts from the honeybee in inhibiting human chronic wound pathogens. Int. Wound J. 2016, 13, 729–737. [Google Scholar] [CrossRef]
- Olofsson, T.C.; Butler, E.; Markowicz, P.; Lindholm, C.; Larsson, L.; Vasquez, A. Lactic acid bacterial symbionts in honeybees—An unknown key to honey’s antimicrobial and therapeutic activities. Int. Wound J. 2016, 13, 668–679. [Google Scholar] [CrossRef]
- Berrios, P.; Fuentes, J.A.; Salas, D.; Carreno, A.; Aldea, P.; Fernandez, F.; Trombert, A.N. Inhibitory effect of biofilm-forming Lactobacillus kunkeei strains against virulent Pseudomonas aeruginosa in vitro and in honeycomb moth (Galleria mellonella) infection model. Benef. Microbes 2018, 9, 257–268. [Google Scholar] [CrossRef]
- Olofsson, T.C.; Vasquez, A. Bacteria isolated from fresh honey or the honey producing tract of honey bees. U.S. Patent No. 9,282,761, 15 March 2016. [Google Scholar]
- Matsuura, D.; Asama, T.; Motoki, H.; Tatefuji, T.; Hashimoto, K. Lactic acid bacterium having IgA production promoting activity, and use thereof. U.S. Patent No. 9,856,451, 2 January 2018. [Google Scholar]
- EFSA. Guidance on the assessment of bacterial antimicrobial susceptibility. EFSA J. 2012, 10, 2740. [Google Scholar]
- Testa, B.; Lombardi, S.J.; Macciola, E.; Succi, M.; Tremonte, P.; Iorizzo, M. Efficacy of olive leaf extract (Olea europaea L. cv Gentile di Larino) in marinated anchovies (Engraulis encrasicolus, L.) process. Heliyon 2019, 5, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Tremonte, P.; Pannella, G.; Succi, M.; Tipaldi, L.; Sturchio, M.; Coppola, R.; Luongo, D.; Sorrentino, E. Antimicrobial activity of Lactobacillus plantarum strains isolated from different environments: A preliminary study. Int. J. Food Res. 2017, 24, 852–859. [Google Scholar]
- Tremonte, P.; Sorrentino, E.; Succi, M.; Tipaldi, L.; Pannella, G.; Ibañez, E.; Mendiola, J.A.; Di Renzo, T.; Reale, A.; Coppola, R. Antimicrobial effect of Malpighia punicifolia and extension of water buffalo steak shelf-life. J. Food Sci. 2016, 81, M97–M105. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, E.; Tremonte, P.; Succi, M.; Iorizzo, M.; Pannella, G.; Lombardi, S.J.; Sturchio, M.; Coppola, R. Detection of antilisterial activity of 3-Phenyllactic acid using Listeria innocua as a model. Front. Microbiol. 2018, 9, 1373. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Lombardi, S.J.; Ganassi, S.; Testa, B.; Ianiro, M.; Letizia, F.; Sorrentino, E. Antagonistic Activity against Ascosphaera apis and Functional Properties of Lactobacillus kunkeei Strains. Antibiotics 2020, 9, 262. [Google Scholar] [CrossRef]
- Succi, M.; Sorrentino, E.; Di Renzo, T.; Tremonte, P.; Reale, A.; Tipaldi, L.; Coppola, R. Lactic acid bacteria in pharmaceutical formulations: Presence and viability of “healthy microorganisms”. J. Pharm. Nutr. Sci. 2014, 4, 66–75. [Google Scholar]
- Succi, M.; Tremonte, P.; Pannella, G.; Tipaldi, L.; Cozzolino, A.; Coppola, R.; Sorrentino, E. Survival of commercial probiotic strains in dark chocolate with high cocoa and phenols content during the storage and in a static in vitro digestion model. J. Funct. Foods 2017, 35, 60–67. [Google Scholar] [CrossRef]
- Succi, M.; Tremonte, P.; Pannella, G.; Tipaldi, L.; Cozzolino, A.; Romaniello, R.; Sorrentino, E.; Coppola, R. Pre-cultivation with selected prebiotics enhances the survival and the stress response of Lactobacillus rhamnosus strains in simulated gastrointestinal transit. Front. Microbial. 2017, 8, 1067. [Google Scholar] [CrossRef]
- Kesen, M.A.; Aiyegoro, O.A. Beneficial Characteristics and Evaluation Criteria of Probiotics. Int. J. Food Biosci. 2018, 1, 25–33. [Google Scholar]
- Liong, M.T.; Shah, N.P. Acid and bile tolerance and cholesterol removal ability of lactobacilli strains. Int. J. Dairy Sci. 2005, 88, 55–66. [Google Scholar] [CrossRef]
- Arredondo, D.; Castelli, L.; Porrini, M.P.; Garrido, P.M.; Eguaras, M.J.; Zunino, P.; Antúnez, K. Lactobacillus kunkeei strains decreased the infection by honey bee pathogens Paenibacillus larvae and Nosema ceranae. Benef. Microbes 2018, 9, 279–290. [Google Scholar] [CrossRef]
- Asama, T.; Kimura, Y.; Kono, T.; Tatefuji, T.; Hashimoto, K.; Benno, Y. Effects of heat-killed Lactobacillus kunkeei YB38 on human intestinal environment and bowel movement: A pilot study. Benef. Microbes 2016, 7, 337–344. [Google Scholar] [CrossRef]
- Piqué, N.; Berlanga, M.; Miñana-Galbis, D. Health benefits of heat-killed (tyndallized) probiotics: An overview. Int. J. Mol. Sci. 2019, 20, 2534. [Google Scholar] [CrossRef] [PubMed]
- Bisson, L.F.; Walker, G.; Ramakrishnan, V.; Luo, Y.; Fan, Q.; Wiemer, E.; Joseph, L. The two faces of Lactobacillus kunkeei: Wine spoilage agent and bee probiotic. Catal. Discov. Pract. 2017, 1, 1–11. [Google Scholar] [CrossRef]
- Filannino, P.; Di Cagno, R.; Addante, R.; Pontonio, E.; Gobbetti, M. Metabolism of fructophilic lactic acid bacteria isolated from the Apis mellifera L. bee gut: Phenolic acids as external electron acceptors. Appl. Environ. Microbiol. 2016, 82, 6899–6911. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Pannella, G.; Lombardi, S.J.; Ganassi, S.; Testa, B.; Succi, M.; Sorrentino, E.; Petrarca, S.; De Cristofaro, A.; Coppola, R.; et al. Inter- and Intra-Species Diversity of Lactic Acid Bacteria in Apis mellifera ligustica Colonies. Microorganisms 2020, 8, 1578. [Google Scholar] [CrossRef]
- Endo, A.; Futagawa-Endo, Y.; Dicks, L.M.T. Isolation and characterization of fructophilic lactic acid bacteria from fructose-rich niches. Syst. Appl. Microbiol. 2009, 32, 593–600. [Google Scholar] [CrossRef]
- Endo, A.; Irisawa, T.; Futagawa-Endo, Y.; Takano, K.; du Toit, M.; Okada, S.; Dicks, L.M. Characterization and emended description of Lactobacillus kunkeei as a fructophilic lactic acid bacterium. Int. J. Syst. Evol. Microbiol. 2012, 62, 500–504. [Google Scholar] [CrossRef]
- Son, S.H.; Yang, S.J.; Jeon, H.L.; Yu, H.S.; Lee, N.K.; Park, Y.S.; Paik, H.D. Antioxidant and immunostimulatory effect of potential probiotic Lactobacillus paraplantarum SC61 isolated from Korean traditional fermented food, jangajji. Microb. Pathog. 2018, 125, 486–492. [Google Scholar] [CrossRef]
- Rodríguez-Díaz, J.; Monedero, V.; Yebra, M.J. Utilization of natural fucosylated oligosaccharides by three novel α-l-fucosidases from a probiotic Lactobacillus casei strain. Appl. Environ. Microbiol. 2011, 77, 703–705. [Google Scholar] [CrossRef]
- Cheon, M.J.; Lim, S.M.; Lee, N.K.; Paik, H.D. Probiotic properties and neuroprotective effects of Lactobacillus buchneri KU200793 isolated from korean fermented foods. Int. J. Mol. Sci. 2020, 21, 1227. [Google Scholar] [CrossRef]
- Van Reenen, C.A.; Dicks, L.M.T. Horizontal gene transfer amongst probiotic lactic acid bacteria and other intestinal microbiota: What are the possibilities? A review. Arch. Microbiol. 2010, 193, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Olmos, A.; Henríquez-Piskulich, P.; Sanchez, C.; Rojas-Herrera, M.; Moreno-Pino, M.; Gómez, M.; Da Rodríguez Silva, R.; Maracaja-Coutinho, V.; Aldea, P.; Trombert, A.N. Draft genome of Chilean honeybee (Apis mellifera) gut strain Lactobacillus kunkeei MP2. Genome Announc. 2014, 2, 1013–1014. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Sadeghi, A.; Rahimi, D.; Purabdolah, H.; Shahryari, S. Postbiotic and Anti-aflatoxigenic Capabilities of Lactobacillus kunkeei as the Potential Probiotic LAB Isolated from the Natural Honey. Probiotics Antimicrob. Proteins 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Coppola, R.; Succi, M.; Tremonte, P.; Reale, A.; Salzano, G.; Sorrentino, E. Antibiotic susceptibility of Lactobacillus rhamnosus strains isolated from Parmigiano Reggiano cheese. Le Lait 2005, 85, 193–204. [Google Scholar] [CrossRef]
- Mathur, S.; Singh, R. Antibiotic resistance in food lactic acid bacteria a review. Int. J. Food Microbiol. 2005, 105, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Salyers, A.; Shoemaker, N.B. Reservoirs of antibiotic resistance genes. Anim. Biotechnol. 2006, 17, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Asenjo, F.; Olmos, A.; Henríquez-Piskulich, P.; Polanco, V.; Aldea, P.; Ugalde, J.A.; Trombert, A.N. Genome sequencing and analysis of the first complete genome of Lactobacillus kunkeei strain MP2, an Apis mellifera gut isolate. PeerJ 2016, 4, e1950. [Google Scholar] [CrossRef][Green Version]
- Butler, È.; Alsterfjord, M.; Olofsson, T.C.; Karlsson, C.; Malmström, J.; Vásquez, A. Proteins of novel lactic acid bacteria from Apis mellifera mellifera: An insight into the production of known extra-cellular proteins during microbial stress. BMC Microbiol. 2013, 13, 235. [Google Scholar] [CrossRef]
- Liu, F.; Jin, P.; Gong, H.; Sun, Z.; Du, L.; Wang, D. Antibacterial and antibiofilm activities of thyme oil against foodborne multiple antibiotics-resistant Enterococcus faecalis. Poult. Sci. J. 2020, 90, 5127–5135. [Google Scholar] [CrossRef]
- Tan, C.A.Z.; Antypas, H.; Kline, K.A. Overcoming the challenge of establishing biofilms in vivo: A roadmap for Enterococci. Curr. Opin. Microbiol. 2020, 53, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Guo, M.J.; Gao, Q.; Yang, J.F.; Yang, L.; Pang, X.L.; Jiang, X.J. The effects of probiotics on total cholesterol: A meta-analysis of randomized controlled trials. Medicine 2018, 97. [Google Scholar] [CrossRef]
- Dixon, A.; Robertson, K.; Yung, A.; Que, M.; Randall, H.; Wellalagodage, D.; Sun, J. Efficacy of Probiotics in Patients of Cardiovascular Disease Risk: A Systematic Review and Meta-analysis. Curr. Hypertens. Rep. 2020, 22, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Li, C.; Qin, Y.; Yin, R.; Du, S.; Ye, F.; Liu, C.; Liu, H.; Wang, M.; Li, Y.; et al. In vitro evaluation of the probiotic and functional potential of Lactobacillus strains isolated from fermented food and human intestine. Anaerobe 2014, 30, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tok, E.; Aslim, B. Cholesterol removal by some lactic acid bacteria that can be used as probiotic. Microbiol. Immunol. 2010, 54, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Lye, H.S.; Rahmat Ali, G.R.; Liong, M.T. Mechanisms of cholesterol removal by lactobacilli under conditions that mimic the human gastrointestinal tract. Int. Dairy J. 2010, 20, 169–175. [Google Scholar] [CrossRef]
- Pereira, D.I.A.; Gibson, G.R. Effects of Consumption of Probiotics and Prebiotics on Serum Lipid Levels in Humans. Crit. Rev. Biochem. Critical. 2002, 37, 259–281. [Google Scholar] [CrossRef]
- Tomaro-Duchesneau, C.; Jones, M.L.; Shah, D.; Jain, P.; Saha, S.; Prakash, S. Cholesterol Assimilation by Lactobacillus Probiotic Bacteria: An In Vitro Investigation. Biomed Res. Int. 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Kumar, M.; Nagpal, R.; Kumar, R.; Hemalatha, R.; Verma, V.; Kumar, A.; Chakraborty, C.; Singh, B.; Marotta, F.; Jain, S.; et al. Cholesterol-lowering probiotics as potential biotherapeutics for metabolic diseases. Exp. Diabetes Res. 2012. [Google Scholar] [CrossRef]
- Perricone, M.; Bevilacqua, A.; Altieri, C.; Sinigaglia, M.; Corbo, M. Challenges for the Production of Probiotic Fruit Juices. Beverages 2015, 1, 95–103. [Google Scholar] [CrossRef]
- Samedi, L.; Charles, A.L. Functional Activity of Four Autochthonous Strains L. paraplantarum AB362736.1, L. plantarum MF369875.1, W. paramesenteroides CP023501.1, and E. faecalis HQ802261.1 in a Probiotic Grape Marmalade during Storage. Antioxidants 2019, 8, 165. [Google Scholar] [CrossRef] [PubMed]
- Randazzo, C.; Pitino, I.; Licciardello, F.; Muratore, G.; Caggia, C. Survival of Lactobacillus rhamnosus probiotic strains in peach jam during storage at different temperatures. Ciência Tecnol. Aliment. 2013, 30, 1–8. [Google Scholar] [CrossRef]


{kind=link}
| K18 | K34 | K45 | DSM 12361 | ATCC 53103 | |
|---|---|---|---|---|---|
| Acid phosphatase | + | + | + | + | + |
| Alkaline phosphatase | + | + | + | + | + |
| Cystine arylamidase | + | + | + | + | + |
| Esterase (C4) | + | + | + | + | + |
| Esterase lipase (C8) | + | + | + | + | + |
| Leucine arylamidase | + | + | + | + | + |
| Lipase (C14) | - | - | - | - | + |
| N-Acetyl-β-glucosaminidase | + | + | + | + | - |
| Naphthol-AS-BI-phosphohydrolase | + | + | + | + | + |
| Trypsin | - | - | - | - | + |
| Valine arylamidase | + | + | + | + | + |
| α-Chymotrypsin | - | - | - | - | + |
| α-Fucosidase | + | + | + | - | + |
| α-Galactosidase | - | - | - | - | - |
| α-Glucosidase | + | + | + | + | + |
| α-Mannosidase | - | - | - | - | + |
| β-Galactosidase | + | + | + | - | + |
| β-Glucosidase | + | + | - | + | + |
| β-Glucuronidase | - | + | - | - | - |
| K18 | K34 | K45 | DSM 12361 | ATCC 53103 | |
|---|---|---|---|---|---|
| Amidon | - | - | - | - | - |
| Amygdaline | + | + | - | - | + |
| Arbutine | + | + | + | - | + |
| D-Adonitol | - | - | - | - | - |
| D-Arabinose | - | - | - | - | + |
| D-Arabitol | - | - | - | - | - |
| D-Cellobiose | + | + | + | - | + |
| D-Fructose | + | + | + | + | + |
| D-Fucose | - | - | - | - | - |
| D-Galactose | + | + | + | + | + |
| D-Glucose | + | + | + | + | + |
| D-Lactose | + | + | + | - | - |
| D-Lyxose | - | - | - | - | - |
| D-Maltose | + | + | + | - | - |
| D-Mannitol | + | + | + | + | + |
| D-Mannose | + | + | + | - | + |
| D-Melezitose | - | - | + | - | + |
| D-Melibiose | - | + | + | + | - |
| D-Raffinose | - | - | + | - | - |
| D-Ribose | + | + | + | - | + |
| D-Saccharose | + | + | + | + | - |
| D-Sorbitol | - | + | + | - | + |
| D-Tagatose | - | - | + | - | + |
| D-Trehalose | + | + | + | + | + |
| D-Turanose | + | + | + | - | - |
| Dulcitol | - | - | + | - | + |
| D-Xylose | - | - | + | - | - |
| Erythritol | - | - | - | - | - |
| Esculine | + | + | + | - | + |
| Gentiobiose | + | + | + | - | + |
| Glycerol | - | - | - | - | - |
| Glycogene | - | - | - | - | - |
| Inositol | - | - | - | - | + |
| Inuline | - | - | - | - | - |
| L-arabinose | + | + | + | - | - |
| L-Arabitol | - | - | - | - | - |
| L-Fucose | - | - | - | - | + |
| L-Rhamnose | - | - | - | - | - |
| L-Sorbose | - | - | + | - | - |
| L-Xylose | - | - | + | - | - |
| Methyl-a-D-Glucopyranoside | - | - | - | - | - |
| Methyl-a-D-Mannopyranoside | - | + | + | - | - |
| Methyl-b-D-Xylopyranoside | - | + | - | - | - |
| N-Acetyl-Glucopyranoside | + | + | + | - | + |
| Potassium 2-Cetogluconate | - | - | - | - | - |
| Potassium 5-Cetogluconate | + | + | - | - | - |
| Potassium Gluconate | - | - | + | - | + |
| Salicine | + | + | - | - | + |
| Xylitol | - | - | - | - | - |
| Antibiotics | Strains | ||||||
|---|---|---|---|---|---|---|---|
| Cut-Off L. rhamnosus * | Cut-Off Lactobacillus Obligate Heterofermentative * | K18 | K34 | K45 | DSM 12361 | ATCC 53103 | |
| Ampicillin | 4 | 2 | R | R | S | R | S |
| Chloramphenicol | 4 | 4 | S | R | S | R | S |
| Clindamycin | 1 | 1 | S | S | S | S | S |
| Erythromycin | 1 | 1 | S | S | S | S | S |
| Gentamycin | 16 | 16 | S | S | S | S | S |
| Kanamycin | 64 | 32 | R | R | S | S | R |
| Streptomycin | 32 | 64 | S | S | S | S | S |
| Tetracycline | 8 | 8 | S | S | S | S | S |
| Indicator Strains | Zone Diameter Breakpoints (R<) * | Producer Strains (Broth Cultures, BC; Cell-Free Supernatants, CFS) | ||||||
|---|---|---|---|---|---|---|---|---|
| K18 | K34 | K45 | DSM 12361 | ATCC 53103 | MRS pH 3.4 | |||
| S. aureus ATCC 29213 | 17.0–26.0 | BC | - | - | - | - | - | - |
| CFS | - | - | - | - | - | - | ||
| E. faecalis ATCC 29212 | 8.0–22.0 | BC | - | 15.4 ± 0.2 | - | - | - | - |
| CFS | - | 14.2 ± 0.3 | - | - | - | - | ||
| E. coli ATCC 11775 | 12.0–25.0 | BC | - | - | - | - | - | - |
| CFS | - | - | - | - | - | - | ||
| E. coli ATCC 25404 | 12.0–25.0 | BC | - | - | - | - | - | - |
| CFS | - | - | - | - | - | - | ||
| P. aeruginosa ATCC 27853 | 15.0–26.0 | BC | 12.4 ± 0.2 | 14.0 ± 0.1 | 12.0 ± 0.2 | - | - | - |
| CFS | 11.0 ± 0.1 | 12.5 ± 0.2 | 11.0 ± 0.1 | - | - | - | ||
| Storage Time (days) | Sugar Syrup (pH 4.2) | Viability (log CFU/mL) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 4 °C | 20 °C | ||||||||||
| K18 | K34 | K45 | DSM 12361 | ATCC 53103 | K18 | K34 | K45 | DSM 12361 | ATCC 53103 | ||
| T0 | A 40% glucose 20% fructose | 8.31 Aa ± 0.04 | 8.09 Ab ± 0.05 | 8.32 Ac ± 0.07 | 8.24 Ab ± 0.06 | 8.12 Ac ± 0.05 | 8.65 Cc ± 0.03 | 8.20 Ac ± 0.05 | 8.42 Bc ± 0.02 | 8.57 Cc ± 0.01 | 8.34 Bb ± 0.05 |
| T7 | 8.23 Ba ± 0.05 | 8.00 Bb ± 0.03 | 8.11 Bb ± 0.07 | 8.19 Bb ± 0.04 | 7.12 Ab ± 0.06 | 5.23 Eb ± 0.05 | 4.30 Db ± 0.02 | 3.21 Cb ± 0.03 | 2.60 Bb ± 0.02 | <1 Aa | |
| T30 | 8.18 Da ± 0.07 | 7.72 Ca ± 0.06 | 7.73 Ca ± 0.03 | 7.44 Ba ± 0.04 | 3.43 Aa ± 0.03 | <1 Aa | <1 Aa | <1 Aa | <1 Aa | <1 Aa | |
| T0 | B 40% glucose 30% fructose | 8.63 Bc ± 0.04 | 8.19 Ac ± 0.02 | 8.22 Ac ± 0.03 | 8.14 Ac ± 0.04 | 8.18 Ac ± 0.03 | 8.72 Cc ± 0.06 | 8.35 Ab ±0.03 | 8.43 Bb ± 0.05 | 8.35 Ab ± 0.04 | 8.32 Ab ± 0.07 |
| T7 | 7.87 Db ± 0.06 | 7.30 Cb ± 0.05 | 7.11 Bb ± 0.04 | 7.09 Bb ± 0.03 | 5.34 Ab ± 0.04 | 1.68 Bb ± 0.01 | <1 Aa | <1 Aa | <1 Aa | <1 Aa | |
| T30 | 5.98 Da ± 0.04 | 5.72 Ca ± 0.03 | 5.73 Ca ± 0.04 | 4.44 Ba ± 0.03 | 1.20 Aa ± 0.03 | <1 Aa | <1 Aa | <1 Aa | <1 Aa | <1 Aa | |
| T0 | C 50% sucrose | 8.23 Ac ± 0.01 | 8.15 Ac ± 0.03 | 8.28 Ac ± 0.04 | 8.20 Ac ± 0.02 | 8.23 Ac ± 0.07 | 8.71 Dc ± 0.03 | 8.25 Ac ± 0.02 | 8.62 Cc ± 0.05 | 8.44 Bc ± 0.02 | 8.23 Ac ± 0.03 |
| T7 | 8.12 Bb ± 0.06 | 8.06 Bb ± 0.01 | 8.00 Bb ± 0.03 | 8.10 Bb ± 0.02 | 7.12 Ab ± 0.05 | 7.15 Db ± 0.05 | 6.95 Cb ± 0.04 | 6.80 Bb ± 0.03 | 6.89 Cb ± 0.02 | 5.50 Ab ± 0.03 | |
| T30 | 7.90 Da ± 0.06 | 7.55 Ba ± 0.03 | 7.50 Ba ± 0.05 | 7.65 Ca ± 0.04 | 4.23 Aa ± 0.04 | 4.63 Ea ± 0.02 | 4.30 Da ± 0.03 | 4.21 Ca ± 0.02 | 3.60 Ba ± 0.03 | <1 Aa | |
| Control | K18 | K34 | K45 | DSM 12361 | ATCC 53103 | |
|---|---|---|---|---|---|---|
| Cholesterol (μg/mL) | 10.03 ± 0.35 | 4.12 ± 0.50 | 10.02 ± 0.60 | 8.24 ± 0.44 | 7.90 ± 0.40 | 4.02 ± 0.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vergalito, F.; Testa, B.; Cozzolino, A.; Letizia, F.; Succi, M.; Lombardi, S.J.; Tremonte, P.; Pannella, G.; Di Marco, R.; Sorrentino, E.; et al. Potential Application of Apilactobacillus kunkeei for Human Use: Evaluation of Probiotic and Functional Properties. Foods 2020, 9, 1535. https://doi.org/10.3390/foods9111535
Vergalito F, Testa B, Cozzolino A, Letizia F, Succi M, Lombardi SJ, Tremonte P, Pannella G, Di Marco R, Sorrentino E, et al. Potential Application of Apilactobacillus kunkeei for Human Use: Evaluation of Probiotic and Functional Properties. Foods. 2020; 9(11):1535. https://doi.org/10.3390/foods9111535
Chicago/Turabian StyleVergalito, Franca, Bruno Testa, Autilia Cozzolino, Francesco Letizia, Mariantonietta Succi, Silvia Jane Lombardi, Patrizio Tremonte, Gianfranco Pannella, Roberto Di Marco, Elena Sorrentino, and et al. 2020. "Potential Application of Apilactobacillus kunkeei for Human Use: Evaluation of Probiotic and Functional Properties" Foods 9, no. 11: 1535. https://doi.org/10.3390/foods9111535
APA StyleVergalito, F., Testa, B., Cozzolino, A., Letizia, F., Succi, M., Lombardi, S. J., Tremonte, P., Pannella, G., Di Marco, R., Sorrentino, E., Coppola, R., & Iorizzo, M. (2020). Potential Application of Apilactobacillus kunkeei for Human Use: Evaluation of Probiotic and Functional Properties. Foods, 9(11), 1535. https://doi.org/10.3390/foods9111535