Zymomonas mobilis in Bread Dough: Characterization of Dough Leavening Performance in Presence of Sucrose




Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Samples
2.2. Flour Characterization and Dough Production
2.3. Microbial Population Counts
2.4. Sugars and Ethanol Determination
2.5. Dough Leavening Properties
2.6. Dough pH Evolution during Leavening
2.7. Statistical Analysis
3. Results and Discussion
3.1. Flour Characterization
3.2. Sugar, Ethanol, pH, and Microbial Evolution during Dough Leavening
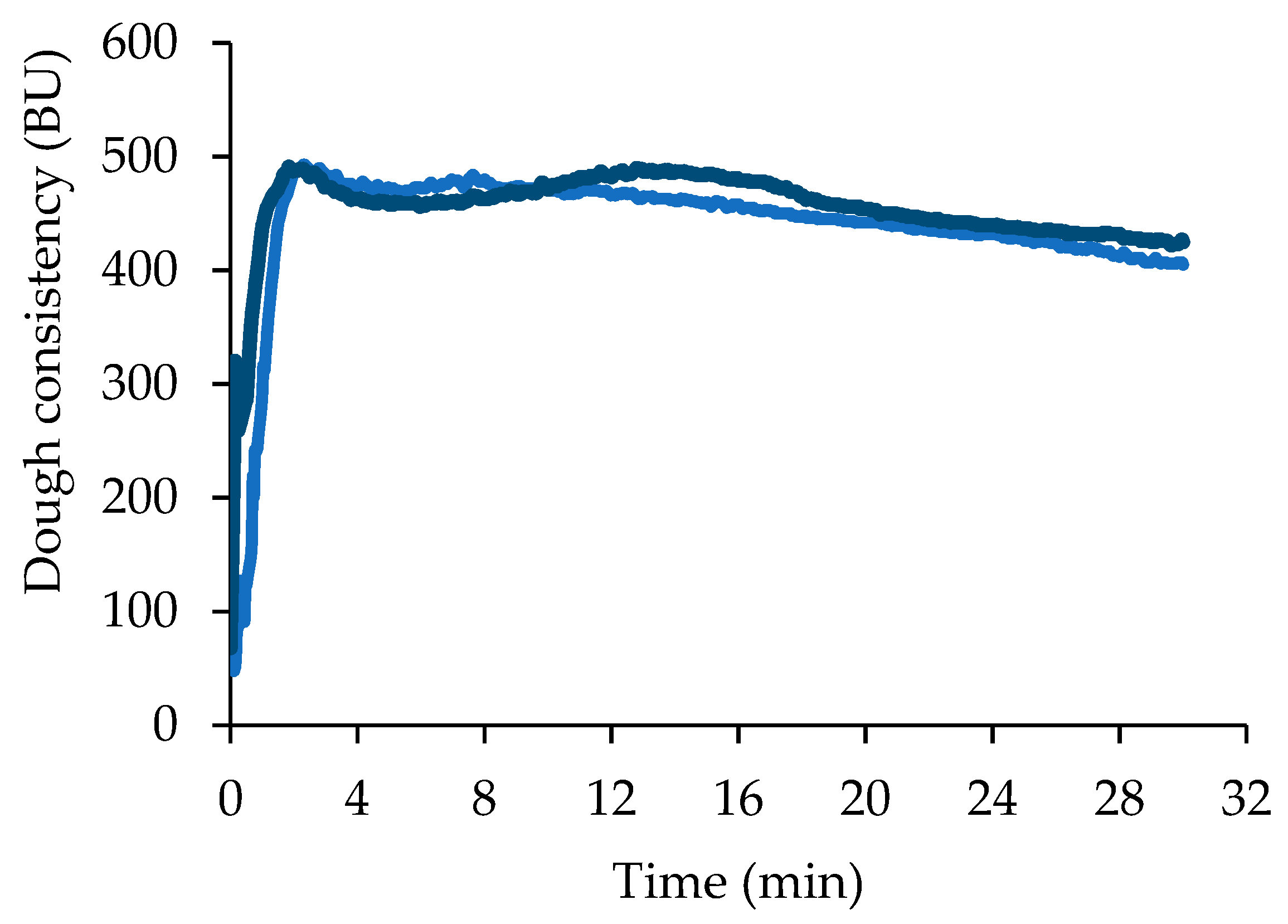
3.3. Dough Leavening Properties
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Salamati, S.; Martins, C.; Kulseng, B. Baker’s yeast (Saccharomyces cerevisiae) antigen in obese and normal weight subjects. Clin. Obes. 2015, 5, 42–47. [Google Scholar] [CrossRef]
- Rinaldi, M.; Perricone, R.; Blank, M.; Perricone, C.; Shoenfeld, Y. Anti-Saccharomyces cerevisiae autoantibodies in autoimmune diseases: From bread baking to autoimmunity. Clin. Rev. Allergy Immunol. 2013, 45, 152–161. [Google Scholar] [CrossRef]
- Caselli, M.; Lo Cascio, N.; Rabitti, S.; Eusebi, L.H.; Zeni, E.; Soavi, C.; Cassol, F.; Zuliani, G.; Zagari, R.M. Pattern of food intolerance in patients with gastro-esophageal reflux symptoms. Minerva Med. 2017, 108, 496–501. [Google Scholar] [CrossRef]
- Zar, S.; Kumar, D.; Benson, M.J. Food hypersensitivity and irritable bowel syndrome. Aliment. Pharmacol. Ther. 2001, 15, 439–449. [Google Scholar] [CrossRef]
- Mansueto, P.; Montalto, G.; Pacor, M.L.; Esposito-Pellitteri, M.; Ditta, V.; Lo Bianco, C.; Leto-Barone, S.M.; Di Lorenzo, G. Food allergy in gastroenterologic diseases: Review of literature. World J. Gastroenterol. 2006, 12, 7744–7752. [Google Scholar] [CrossRef]
- Boyce, J.A.; Assa’ad, A.; Burks, A.W.; Jones, S.M.; Sampson, H.A.; Wood, R.A.; Fenton, M.J. Guidelines for the diagnosis and management of food allergy in the United States: Report of the NIAID-sponsored expert panel. J. Allergy Clin. Immunol. 2010, 126, S1–S58. [Google Scholar] [CrossRef] [PubMed]
- Brunner, B.; Scheurer, U.; Seibold, F. Differences in yeast intolerance between patients with Crohn’s disease and ulcerative colitis. Dis. Colon Rectum 2007, 50, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Desplat-Jégo, S.; Johanet, C.; Escande, A.; Goetz, J.; Fabien, N.; Olsson, N.; Humbel, R.L. Update on anti-Saccharomyces cerevisiae antibodies, anti-nuclear associated anti-neutrophil antibodies and antibodies to exocrine pancreas detected by indirect immunofluorescence as biomarkers in chronic inflammatory bowel diseases: Results of a multicenter study. World J. Gastroenterol. 2007, 13, 2312–2318. [Google Scholar] [CrossRef] [PubMed]
- Forcione, D.G.; Rosen, M.J.; Kisiel, J.B.; Sands, B.E. Anti-Saccharomyces cerevisiae antibody (ASCA) positivity is associated with increased risk for early surgery in Crohn’s disease. Gut 2004, 53, 1117–1122. [Google Scholar] [CrossRef]
- Bansal, R.A.; Tadros, S.; Bansal, A.S. Beer, Cider, and Wine Allergy. Case Rep. Immunol. 2017, 2017, 7958924. [Google Scholar] [CrossRef]
- De Bellis, P.; Rizzello, C.G.; Sisto, A.; Valerio, F.; Lonigro, S.L.; Conte, A.; Lorusso, V.; Lavermicocca, P. Use of a selected Leuconostoc citreum strain as a starter for making a “yeast-free” bread. Foods 2019, 8, 70. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Tonomura, K. Dough-leavening ability by Zymomonas mobilis and its application to breadmaking. J. Food Sci. 1994, 59, 171–174. [Google Scholar] [CrossRef]
- Krishnan, M.S.; Taylor, F.; Davison, B.H.; Nghiem, N.P. Economic analysis of fuel ethanol production from corn starch using fluidized-bed reactor. Bioresour. Technol. 2000, 75, 99–105. [Google Scholar] [CrossRef]
- Sprenger, G.A. Carbohydrate metabolism in Zymomonas mobilis: A catabolic highway with some scenic routes. FEMS Microbiol. Lett. 1996, 145, 301–307. [Google Scholar] [CrossRef]
- Stear, C.A. Handbook of Breadmaking Technology; Elsevier Science Publishers Ltd.: Essex, UK, 1990; Chapter 1.8; pp. 306–372. [Google Scholar]
- Martínez-Anaya, M.A.; Pitarch, B.; Bayarri, P.; Benedito de Barber, C. Microflora of the sourdoughs of wheat flour bread. X. Interactions between yeasts and lactic acid bacteria in wheat doughs and their effects on bread quality. Cereal Chem. 1990, 67, 85–91. [Google Scholar]
- Musatti, A.; Mapelli, C.; Foschino, R.; Picozzi, C.; Rollini, M. Unconventional bacterial association for dough leavening. Int. J. Food Microbiol. 2016, 237, 28–34. [Google Scholar] [CrossRef]
- Gobbetti, M.; Corsetti, A. Lactobacillus sanfrancisco a key sourdough lactic acid bacterium: A review. Food Microbiol. 1997, 14, 175–187. [Google Scholar] [CrossRef]
- Musatti, A.; Mapelli, C.; Rollini, M.; Foschino, R.; Picozzi, C. Can Zymomonas mobilis substitute Saccharomyces cerevisiae in cereal dough leavening? Foods 2018, 7, 61. [Google Scholar] [CrossRef]
- Musatti, A.; Rollini, M.; Sambusiti, C.; Manzoni, M. Zymomonas mobilis: Biomass production and use as dough leavening agent. Ann. Microbiol. 2015, 65, 1583–1589. [Google Scholar] [CrossRef]
- Picozzi, C.; Mariotti, M.; Cappa, C.; Tedesco, B.; Vigentini, I.; Foschino, R.; Lucisano, M. Development of a Type I gluten-free sourdough. Lett. Appl. Microbiol. 2016, 62, 119–125. [Google Scholar] [CrossRef]
- Cappa, C.; Lucisano, M.; Mariotti, M. Influence of Psyllium, sugar beet fibre and water on gluten-free dough properties and bread quality. Carbohydr. Polym. 2013, 98, 1657–1666. [Google Scholar] [CrossRef] [PubMed]
- Cappa, C.; Lucisano, M.; Raineri, A.; Fongaro, L.; Foschino, R.; Mariotti, M. Gluten-free bread: Influence of sourdough and compressed yeast on proofing and baking properties. Foods 2016, 5, 69. [Google Scholar] [CrossRef] [PubMed]
- Aldovrandi, L.; Vitali, F. Criteri di valutazione del frumento tenero da parte dell’industria molitoria. Molini d’Italia 1995, 46, 16–19. [Google Scholar]
- Musatti, A.; Manzoni, M.; Rollini, M. Post-fermentative production of glutathione by baker’s yeast (S. cerevisiae) in compressed and dried forms. New Biotechnol. 2013, 30, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Verheyen, C.; Albrecht, A.; Elgeti, D.; Jekle, M.; Becker, T. Impact of gas formation kinetics on dough development and bread quality. Food Res. Int. 2015, 76, 860–866. [Google Scholar] [CrossRef]
- Struyf, N.; Van der Maelen, E.; Hemdane, S.; Verspreet, J.; Verstrepen, K.J.; Courtin, C.M. Bread dough and baker’s yeast: An uplifting synergy. Compr. Rev. Food Sci. Food Saf. 2017, 16, 850–867. [Google Scholar] [CrossRef]
- Novak, S.; Zechner-Krpan, V.; Marić, V. Regulation of maltose transport and metabolism in Saccharomyces cerevisiae. Food Technol. Biotechnol. 2004, 42, 213–218. [Google Scholar] [CrossRef][Green Version]
- Mariotti, M.; Alamprese, C. About the use of different sweeteners in baked goods. Influence on the mechanical and rheological properties of the doughs. LWT Food Sci. Technol. 2012, 48, 9–15. [Google Scholar] [CrossRef]
- Rollini, M.; Casiraghi, E.; Pagani, M.A.; Manzoni, M. Technological performances of commercial yeast strains (Saccharomyces cerevisiae) in different complex dough formulations. Eur. Food Res. Technol. 2007, 226, 19–24. [Google Scholar] [CrossRef]




{kind=link}
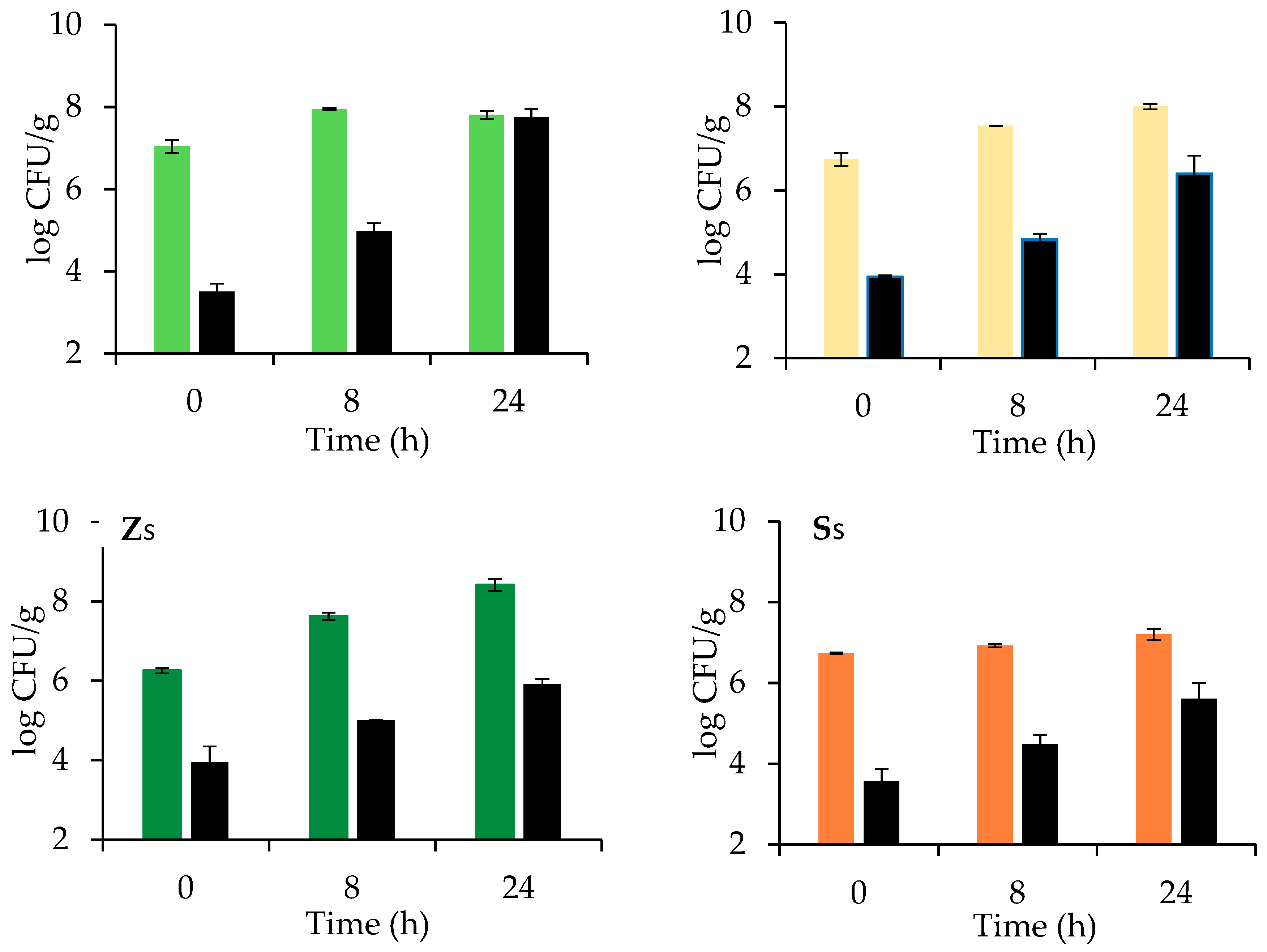{kind=link}
{kind=link}
| Sample 1 | Flour (g/100 g) | Water (g/100 g) | Sucrose (g/100 g) | Z. mobilis (log CFU/g) | S. cerevisiae (log CFU/g) |
|---|---|---|---|---|---|
| Z | 100 | 63.3 | - | 7 | - |
| Zs | 100 | 57.3 | 2.5 | 7 | - |
| S | 100 | 63.3 | - | - | 7 |
| Ss | 100 | 57.3 | 2.5 | - | 7 |
| Leavening Time (h) | Sample | ||||
|---|---|---|---|---|---|
| Z | Zs | S | Ss | ||
| Maltose | 0 | 1.84 ± 0.38 bc | 1.49 ± 0.04 ab | 2.09 ± 0.04 c | 1.27 ± 0.13 a |
| 8 | 3.08 ± 0.10 b | 2.36 ± 0.28 a | 2.13 ± 0.09 a | 2.55 ± 0.11 a | |
| 24 | 3.85 ± 0.08 b | 3.38 ± 0.37 b | n.d. | 2.21 ± 0.17 a | |
| Glucose | 0 | 0.27 ± 0.05 b | 0.03 ± 0.01 a | 0.36 ± 0.01 c | 0.62 ± 0.02 d |
| 8 | 0.02 ± 0.01 a | 0.07 ± 0.03 a | 0.09 ± 0.01 a | 0.79 ± 0.05 b | |
| 24 | n.d. | 0.07 ± 0.02 a | n.d. | 0.35 ± 0.05 b | |
| Sucrose | 0 | n.d. | 2.78 ± 0.36 a | n.d. | 2.05 ± 0.04 a |
| 8 | n.d. | 2.76 ± 0.29 b | n.d. | 0.17 ± 0.07 a | |
| 24 | n.d. | n.d. | n.d. | n.d. | |
| Ethanol | 0 | n.d. | n.d. | n.d. | n.d. |
| 8 | 0.24 ± 0.01 a | 0.10 ± 0.07 a | 1.18 ± 0.14 b | 0.81 ± 0.01 b | |
| 24 | 0.75 ± 0.01 a | 1.72 ± 0.01 b | 3.49 ± 0.29 d | 2.58 ± 0.05 c | |
| Leavening Indices 1 | Z | Zs | S | Ss |
|---|---|---|---|---|
| Hm (mm) | 14.95 ± 0.21 a | 39.7 ± 1.41 b | 45.95 ± 2.33 c | 49.45 ± 1.06 c |
| T1 (h) | 11.16 ± 0.06 c | 11.55 ± 0.21 c | 7.87 ± 0.35 b | 7.87 ± 0.35 b |
| LLT (h) | 7.12 ± 0.12 d | 6.07 ± 0.02 c | 3.19 ± 0.12 b | 1.56 ± 0.04 a |
| LR (mm/h) | 5.16 ± 0.05 a | 6.86 ± 1.54 a | 14.56 ± 0.95 c | 10.60 ± 0.10 b |
| CO2-TOT (mL) | 855 ± 136 a | 1542 ± 38 b | 2231 ± 20 c | 2086 ± 136 c |
| CO2-RET (mL) | 739 ± 3 a | 1368 ± 23 b | 1570 ± 27 c | 1574 ± 2 c |
| Tx (h) | 10.75 ± 0.78 c | 11.26 ± 0.34 c | 6.57 ± 0.07 a | 8.29 ± 0.12 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Musatti, A.; Cappa, C.; Mapelli, C.; Alamprese, C.; Rollini, M. Zymomonas mobilis in Bread Dough: Characterization of Dough Leavening Performance in Presence of Sucrose. Foods 2020, 9, 89. https://doi.org/10.3390/foods9010089
Musatti A, Cappa C, Mapelli C, Alamprese C, Rollini M. Zymomonas mobilis in Bread Dough: Characterization of Dough Leavening Performance in Presence of Sucrose. Foods. 2020; 9(1):89. https://doi.org/10.3390/foods9010089
Chicago/Turabian StyleMusatti, Alida, Carola Cappa, Chiara Mapelli, Cristina Alamprese, and Manuela Rollini. 2020. "Zymomonas mobilis in Bread Dough: Characterization of Dough Leavening Performance in Presence of Sucrose" Foods 9, no. 1: 89. https://doi.org/10.3390/foods9010089
APA StyleMusatti, A., Cappa, C., Mapelli, C., Alamprese, C., & Rollini, M. (2020). Zymomonas mobilis in Bread Dough: Characterization of Dough Leavening Performance in Presence of Sucrose. Foods, 9(1), 89. https://doi.org/10.3390/foods9010089