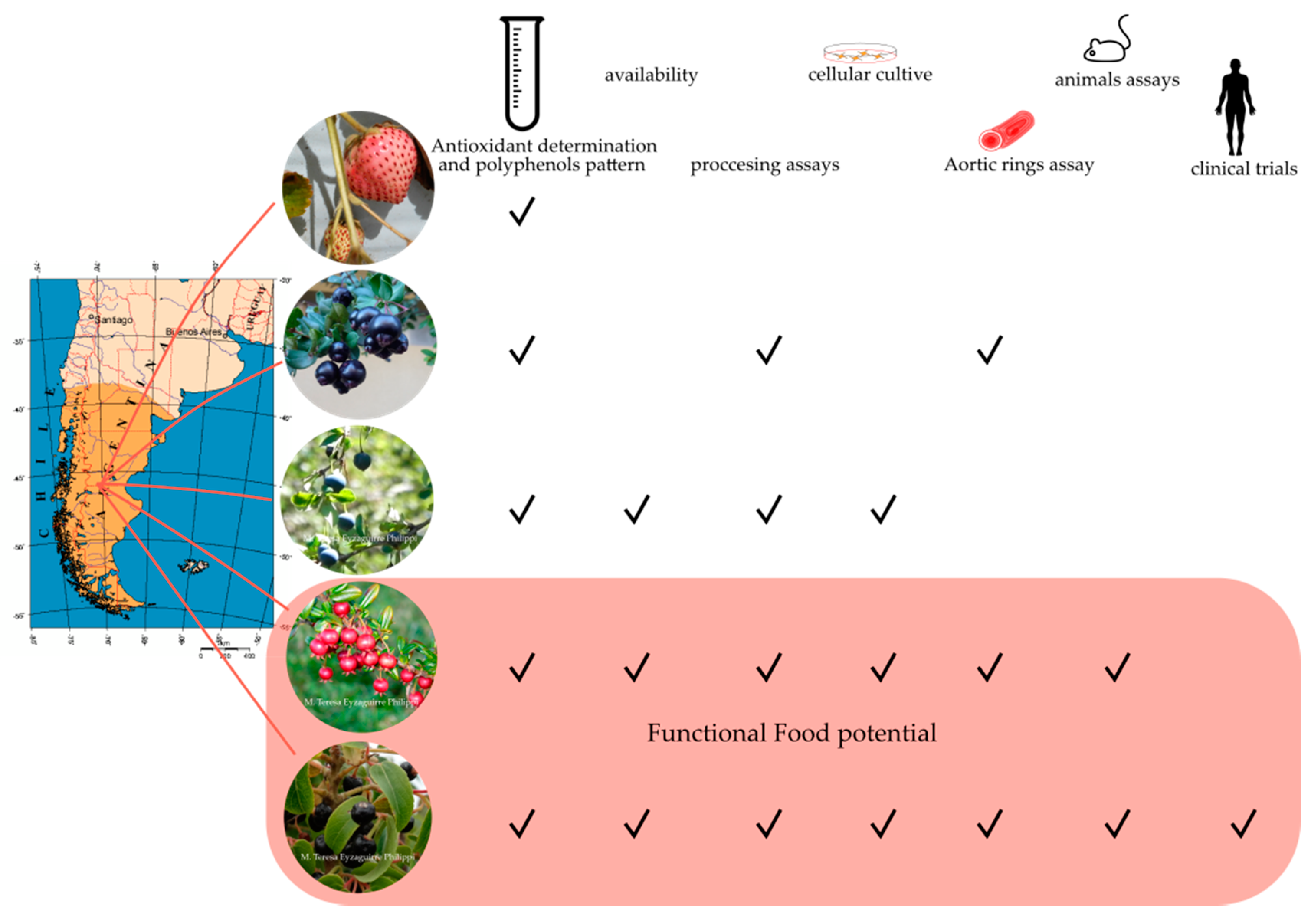
Patagonian Berries: Healthy Potential and the Path to Becoming Functional Foods



Abstract
1. Introduction
2. Quality Aspects and Bioactive Compounds of Patagonian Berries
2.1. Fruit Quality
2.1.1. Color and Appearance
2.1.2. Flavor
2.1.3. Texture
2.2. Antioxidant Capacity
2.2.1. Phenolic Content and Composition
2.2.2. Anthocyanins
3. Effects of Processing on Bioactive Compounds
3.1. Drying Process
3.2. Microencapsulation for Liquid Preparation
4. Healthy Potential of Patagonian berries
4.1. Polyphenols and Anti-Inflammatory Effects
4.1.1. Polyphenols and Metabolic Syndrome
4.1.2. Cardiovascular Effects
5. Some Commercial Aspects
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Coronato, A.; Coronato, F.; Mazzoni, E.; Vázquez, M. The physical geography of Patagonia and Tierra del Fuego. The Late Cenozoic of Patagonia and Tierra del Fuego; Rabassa, J., Ed.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 13–56. [Google Scholar]
- Hoffmann, A.; Farga, C.; Lastra, J.; Veghazi, E. Plantas Medicinales de Uso Comun en Chile; Fundación Claudio Gay: Santiago de Chile, Chile, 1992; 257p. [Google Scholar]
- Mösbach, E.W. Botánica Indígena de Chile; Museo Chileno de Arte Precolombino; Editorial Andrés Bello; Fundación Andes: Santiago de Chile, Chile, 1992; 140p. [Google Scholar]
- Ladio, A.H.; Lozada, M. Patterns of use and knowledge of wild edible plants in distinct ecological environments: A case study of a Mapuche community from Nothwestern Patagonia. Biodivers. Conserv. 2004, 13, 1153–1173. [Google Scholar] [CrossRef]
- Estomba, D.; Ladio, A.; Lozada, M. Medicinal wild plant knowledge and gathering patterns in a Mapuche community from North-western Patagonia. J. Ethnopharmacol. 2006, 103, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Forestier, J.; León-Lobos, L.; Marticorena, A.; Celis-Diez, J.L.; Giovannini, P. Native Useful Plants of Chile: A Review and Use Patterns. Econ. Bot. 2019, 73, 112–126. [Google Scholar] [CrossRef]
- Barreau, A.; Ibarra, J.T.; Wyndham, F.S.; Rojas, A.; Kozak, R.A. How can we teach our children if we cannot access the forest? Generational change in mapuche knowledge of wild edible plants in Andean temperature ecosystems of Chile. J. Ethnobiol. 2016, 36, 412–432. [Google Scholar] [CrossRef]
- Rivera, D.; Verde, A.; Fajardo, J.; Inocencio, C.; Obon, C.; Heinrich, M. Guia Etnobotanica de los Alimentos Locales Recolectados en la Provincia de Albacete; Instituto de Estudios Albacetenses; Diputación de Albacete: Albacete, Spain, 2006. [Google Scholar]
- Egea, I.; Sánchez-Bel, P.; Romojaro, F.; Pretel, M.T. Six edible wild fruits as potential antioxidant additives or nutritional supplements. Plant Foods Hum. Nutr. 2010, 65, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.C.; Steffens, R.S.; Jablonski, A.; Hertz, P.F.; Rios Ade, O.; Vizzotto, M.; Flôres, S.H. Characterization and antioxidant potential of Brazilian fruits from the Myrtaceae family. J. Agric. Food Chem. 2012, 60, 3061–3067. [Google Scholar] [CrossRef]
- Speisky, H.; López Alarcón, C.; Gómez, M.; Fuentes, J.; Sandoval Acuña, C. First web-based database on total phenolics and oxygen radical absorbance capacity (ORAC) of fruits produced and consumed within the South Andes Region of South America. J. Agric. Food Chem. 2012, 60, 8851–8859. [Google Scholar] [CrossRef]
- Ramos, A.S.; Souza, R.O.S.; Boleti, A.P.A.; Bruginski, E.R.D.; Lima, E.S.; Campos, F.R.; Machado, M.B. Chemical characterization and antioxidant capacity of the araçá-pera (Psidium acutangulum): An exotic Amazon fruit. Food Res. Int. 2015, 75, 315–327. [Google Scholar] [CrossRef]
- Kongkachuichai, R.; Charoensiri, R.; Yakoh, K.; Kringkasemsee, A.; Insung, P. Nutrients value and antioxidant content of indigenous vegetables from Southern Thailand. Food Chem. 2015, 173, 838–846. [Google Scholar] [CrossRef]
- Barros, R.G.C.; Andrade, J.K.S.; Denadai, M.; Nunes, M.L.; Narain, N. Evaluation of bioactive compounds potential and antioxidant activity in some Brazilian exotic fruit residues. Food Res. Int. 2017, 102, 84–92. [Google Scholar] [CrossRef]
- Ulloa-Inostroza, E.M.; Ulloa-Inostroza, E.G.; Alberdi, M.; Peña-Sanhueza, D.; González-Villagra, J.; Jaakola, L.; Reyes-Díaz, M. Native Chilean Fruits and the Effects of their Functional Compounds on Human Health, Superfood and Functional Food Viduranga Waisundara. Available online: https://www.intechopen.com/books/superfood-and-functional-food-an-overview-of-their-processing-and-utilization/native-chilean-fruits-and-the-effects-of-their-functional-compounds-on-human-health (accessed on 29 June 2019).
- Schmeda-Hirschmann, G.; Jiménez-Aspee, F.; Theoduloz, C.; Ladio, A. Patagonian berries as native food and medicine. J. Ethnopharmacol. 2019, 241, 111979. [Google Scholar] [CrossRef] [PubMed]
- Simirgiotis, M.J.; Theoduloz, C.; Caligari, P.D.S.; Schmeda-Hirschmann, G. Comparison of phenolic composition and antioxidant properties of two native Chilean and one domestic strawberry genotypes. Food Chem. 2009, 113, 377–385. [Google Scholar] [CrossRef]
- Simirgiotis, M.J.; Bórquez, J.; Schmeda-Hirschmann, G. Antioxidant capacity, polyphenol content and tandem HPLCDAD- ESI/MS profiling of phenolic compounds from the South American berries Luma apiculata and L. chequen. Food Chem. 2013, 139, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Brito, A.; Areche, C.; Sepúlveda, B.; Kennelly, E.; Simirgiotis, M. Anthocyanin characterization, total phenolic quantification and antioxidant features of some Chilean edible berry extracts. Molecules 2014, 19, 10936–10955. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, A.; Hermosín, I.; Mardones, C.; Vergara, C.; Herlitz, C.; Vega, M.; Dorau, C.; Winterhalter, P.; Von Baer, D. Polyphenols and antioxidant activity of calafate (Berberis microphylla) fruits and other native berries from southern Chile. J. Agric. Food Chem. 2010, 58, 6081–6089. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, L.; Valdenegro, M.; Gómez, M.G.; Ayala-Raso, A.; Quiroga, E.; Martínez, J.P.; Vinet, R.; Caballero, E.; Figueroa, C.R. Characterization of fruit development and potential health benefits of arrayan (Luma apiculata), a native berry of South America. Food Chem. 2016, 196, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Rozzi, R.; Massardo, F. Las Plantas Medicinales Chilenas II y III. Informe Nueva Medicina 1994, 1, 16–17. [Google Scholar]
- Massardo, F.; Rozzi, R. Usos medicinales de la flora nativa chilena. Ambiente y Desarrollo 1996, 12, 76–81. [Google Scholar]
- Alvarado, J.; Schoenlau, F.; Leschot, A.; Salgad, A.M.; Vigil Portales, P. Delphinol® standardized maqui berry extract significantly lowers blood glucose and improves blood lipid profile in prediabetic individuals in three-month clinical trial. Panminerva Med. 2016, 58 (Suppl. 1), 1–6. [Google Scholar]
- Alvarado, J.; Leschot, A.; Olivera Nappa, Á.; Salgado, A.; Rioseco, H.; Lyon, C.; Vigil, P. Delphinidin-Rich Maqui Berry Extract (Delphinol (R)) Lowers Fasting and Postprandial Glycemia and Insulinemia in Prediabetic Individuals during Oral Glucose Tolerance Tests. BioMed Res. Int. 2016, 2016b, 9070537. [Google Scholar] [CrossRef]
- Watson, R.R.; Schönlau, F. Nutraceutical and antioxidant effects of a delphinidin-rich maqui berry extract Delphinol®: A review. Minerva Cardioangiol. 2015, 63 (Suppl. 1), 1–12. [Google Scholar] [PubMed]
- Reyes-Farias, M.; Vasquez, K.; Ovalle-Marin, A.; Fuentes, F.; Parra, C.; Quitral, V.; Jimenez, P.; Garcia-Diaz, D.F. Chilean native fruit extracts inhibit inflammation linked to the pathogenic interaction between adipocytes and macrophages. J. Med. Food 2015, 18, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Barrett, D.M.; Beaulieu, J.C.; Shewfelt, R. Color, flavor, texture, and nutritional quality of fresh-cut fruits and vegetables: Desirable levels, instrumental and sensory measurement, and the effects of processing. Crit. Rev. Food Sci. Nutr. 2010, 50, 369–389. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A. Evaluation of quality of fruits and vegetables. In Food Quality; Irving, G.W., Jr., Hoover, S.R., Eds.; American Association for the Advancement of Science: Washington, DC, USA, 1965; pp. 9–18. [Google Scholar]
- Fuentes, L.; Figueroa, C.R.; Valdenegro, M. Recent Advances in Hormonal Regulation and Cross-Talk during Non-Climacteric Fruit Development and Ripening. Horticulturae 2019, 5, 45. [Google Scholar] [CrossRef]
- Figueroa, C.R.; Pimentel, P.; Gaete-Eastman, C.; Moya, M.; Herrera, R.; Caligari, P.D.; Moya-León, M.A. Softening rate of the Chilean strawberry (Fragaria chiloensis) fruit reflects the expression of polygalacturonase and pectate lyase genes. Postharv. Biol. Technol. 2008, 49, 210–220. [Google Scholar] [CrossRef]
- Concha, C.M.; Figueroa, N.E.; Poblete, L.A.; Oñate, F.A.; Schwab, W.; Figueroa, C.R. Methyl jasmonate treatment induces changes in fruit ripening by modifying the expression of several ripening genes in Fragaria chiloensis fruit. Plant Physiol. Biochem. 2013, 70, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Bigotes, A.; Figueroa, P.M.; Figueroa, C.R. Jasmonate metabolism and its relationship with abscisic acid during strawberry fruit development and ripening. J. Plant Growth Regul. 2018, 37, 101–113. [Google Scholar] [CrossRef]
- Fredes, C.; Montenegro, G.; Zoffoli, J.P.; Gómez, M.; Robert, P. Polyphenol content and antioxidant activity of maqui (Aristotelia chilensis Molina Stuntz) during fruit development and maturation in Central Chile. Chil. J. Agric. Res. 2012, 72, 582–589. [Google Scholar] [CrossRef]
- Rodriguez, R.; Marticorena, C.; Alarcón, D.; Baeza, C.; Cavieres, L.; Finot, V.L.; Fuentes, N.; Kiessling, A.; Mihoc, M.; Pauchard, A.; et al. Catálogo de las plantas vasculares de Chile. Gayana Botánica 2018, 75, 1–430. [Google Scholar] [CrossRef]
- Gomes, F.C.; Lacerda, I.C.; Libkind, D.; Lopes, C.; Carvajal, E.J.; Rosa, C.A. Traditional foods and beverages from South America: Microbial communities and production strategies. Ind. Ferment. Food Process. Nutr. Sour. Prod. Strateg. 2009, 3, 79–114. [Google Scholar]
- Hoffmann, A.E. Flora Silvestre de Chile, 5th ed.; Zona Araucana Edicione; Fundación Claudio Gay: Santiago de Chile, Chile, 2005; 257p. [Google Scholar]
- Quispe-Fuentes, I.; Vega-Gálvez, A.; Aranda, M. Evaluation of phenolic profiles and antioxidant capacity of maqui (Aristotelia chilensis) berries and their relationships to drying methods. J. Sci. Food Agric. 2018, 98, 4168–4176. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, K.; Ah-Hen, K.S.; Vega-Gálvez, A.; Vásquez, V.; Quispe-Fuentes, I.; Rojas, P.; Lemus-Mondaca, R. Changes in bioactive components and antioxidant capacity of maqui, Aristotelia chilensis [Mol] Stuntz, berries during drying. LWT Food Sci. Technol. 2016, 65, 537–542. [Google Scholar] [CrossRef]
- Romo, R.; Bastías, J.M. Estudio de Mercado del Maqui. PYT-0215-0219. Perspectiva del Mercado Internacional para el Desarrollo de la Industria del Maqui: Un Análisis de las Empresas en Chile. Available online: http://bibliotecadigital.fia.cl/bitstream/handle/20.500.11944/146064/Informe%20Final%20Estudio%20de%20Mercado%20MAQUI.pdf?sequence=1&isAllowed=y (accessed on 28 May 2019).
- Estudio Preparación de Expedientes Técnicos para la Presentación y Solicitud de Autorización de Alimentos Nuevos o Tradicionales de Terceros Países para Exportar a la Unión Europea. Available online: https://www.odepa.gob.cl/wp-content/uploads/2017/12/Informe-Estudio-Novel-Foods-y-anexos-1.pdf (accessed on 28 May 2019).
- Puente-Díaz, L.K.; Ah-Hen, A.; Vega-Gálvez, R.; Lemus-Mondaca; Di Scala, K. Combined infrared-convective drying of murta (Ugni molinae Turcz.) berries: Kinetic modeling and quality assessment. Dry Technol. 2013, 31, 329–338. [Google Scholar]
- Figueroa, C.R.; Concha, C.M.; Figueroa, N.E.; Tapia, G. Frutilla Chilena Nativa Fragaria chiloensis. Available online: http://www.procisur.org.uy/adjuntos/13d4953eae2f_Chilena_PROCISUR.pdf (accessed on 28 May 2019).
- Lavin, A.; Vega, A. Caracterizacion de frutos de murtilla (Ugni molinae Turcz.) en el area de Cauquenes. Agricultura Tecnica 1996, 56, 64–67. [Google Scholar]
- Arena, M.E.; Curvetto, N. Berberis buxifolia fruiting: Kinetic growth behavior and evolution of chemical properties during the fruiting period and different growing seasons. Scientia Horticulturae 2008, 118, 120–127. [Google Scholar] [CrossRef]
- Stevens, M.A.; Kader, A.A.; Albright, M. Potential for increasing tomato flavor via increased sugar and acid content. J. Am. Soc. Horticult. Sci. 1979, 104, 40–42. [Google Scholar]
- Batista-Silva, W.; Nascimento, V.L.; Medeiros, D.B.; Nunes-Nesi, A.; Ribeiro, D.M.; Zsögön, A.; Araújo, W.L. Modifications in Organic Acid Profiles During Fruit Development and Ripening: Correlation or Causation? Front Plant. Sci. 2018, 9, 1689. [Google Scholar] [CrossRef] [PubMed]
- Cherian, S.; Figueroa, C.R.; Nair, H. ‘Movers and shakers’ in the regulation of fruit ripening: A cross-dissection of climacteric versus non-climacteric fruit. J. Exp. Bot. 2014, 65, 4705–4722. [Google Scholar] [CrossRef] [PubMed]
- Arena, M.E.; Postemsky, P.; Curvetto, N.R. Accumulation patterns of phenolic compounds during fruit growth and ripening of Berberis buxifolia, a native Patagonian species. N. Z. J. Bot. 2012, 50, 1528. [Google Scholar] [CrossRef]
- Arena, M.E.; Zuleta ADyner, L.; Constenla DCeci, L.; Curvetto, N.R. Berberis buxifolia fruit growth and ripening: Evolution in carbohydrate and organic acid contents. Scientia Horticulturae 2013, 158, 52–58. [Google Scholar] [CrossRef]
- Gonzalez, M.; Gaete-Eastman, C.; Valdenegro, M.; Figueroa, C.R.; Fuentes, L.; Herrera, R.; Moya-León, M.A. Aroma development during ripening of Fragaria chiloensis fruit and participation of an alcohol acyltransferase (FcAAT1) gene. J. Agric. Food Chem. 2009, 57, 9123–9132. [Google Scholar] [CrossRef] [PubMed]
- Scheuermann, E.; Seguel, I.; Montenegro, A.; Bustos ROHormazabal, E.; Quiroz, A. Evolution of aroma compounds of murtilla fruits (Ugni molinae Turcz) during storage. J. Sci. Food Agric. 2008, 88, 485–492. [Google Scholar] [CrossRef]
- Hummer, K.E.; Hancock, J. Strawberry Genomics: Botanical History, Cultivation, Traditional Breeding, and New Technologies. In Genetics and Genomics of Rosaceae. Plant Genetics and Genomics: Crops and Models; Folta, K.M., Gardiner, S.E., Eds.; Springer: New York, NY, USA, 2009; Volume 6. [Google Scholar]
- Chen, L.; Opara, U.L. Texture measurement approaches in fresh and processed foods—A review. Food Res. Int. 2013, 51, 823–835. [Google Scholar] [CrossRef]
- Figueroa, C.R.; Rosli, H.G.; Civello, P.M.; Martínez, G.A.; Herrera, R.; Moya-León, M.A. Changes in cell wall polysaccharides and cell wall degrading enzymes during ripening of Fragaria chiloensis and Fragaria× ananassa fruits. Scientia Horticulturae 2010, 124, 454–462. [Google Scholar] [CrossRef]
- Valdenegro, M.; Fuentes, L.; Urtubia, A.; Herrera, R.; Moya-Leon, M.A. Modified atmosphere packaging preserves quality and antioxidant properties in Fragaria chiloensis. J. Biotechnol. 2010, 150 (Suppl. 1), S335. [Google Scholar]
- Vicente, A.R.; Ortugno, C.; Rosli, H.; Powell, A.L.; Greve, L.C.; Labavitch, J.M. Temporal sequence of cell wall disassembly events in developing fruits. 2. Analysis of blueberry (Vaccinium species). J. Agric. Food Chem. 2007, 55, 4125–4130. [Google Scholar] [CrossRef]
- Defilippi, B.; Robledo, P.; Becerra, C.; Seguel, I. Comportamiento en postcosecha de dos variedades de murtilla: South Pearl INIA y Red Pearl INIA. Available online: http://www.inia.cl/postcosecha/files/2015/02/Defilippi_murtilla_Iquique.pdf (accessed on 26 June 2019).
- Robledo, P.; Soto-Alvear, S.; Seguel, I.; Defilippi, B. Evaluación de Tecnologías de Postcosecha en Murtilla. Available online: http://www.inia.cl/postcosecha/files/2015/02/Poster_murtilla2012.pdf (accessed on 25 May 2019).
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085. [Google Scholar] [CrossRef] [PubMed]
- Landrum, L.; Donoso, C. Ugni molinae (Myrtaceae), a potential fruit crop for regions of Mediterranean, maritime and subtropical climates. Econ. Bot. 1990, 44, 536–539. [Google Scholar]
- Rice-Evans, C.; Miller, N.J.; Paganga, G. Structure antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Rice-Evans, C.; Miller, N.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Soto-Vaca, A.; Gutierrez, A.; Losso, J.N.; Xu, Z.; Finley, J.W. Evolution of phenolic compounds from color and flavor problems to health benefits. J. Agric. Food Chem. 2012, 60, 6658–6677. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Manach, C.; Morand, C.; Remesy, C. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Russo, B.; Picconi, F.; Malandrucco, I.; Frontoni, S. Flavonoids and Insulin-Resistance: From Molecular Evidences to Clinical Trials. Int. J. Mol. Sci. 2019, 20, 2061. [Google Scholar] [CrossRef] [PubMed]
- Wikimedia Commons. Available online: https://commons.wikimedia.org/wiki/Main_Page (accessed on 26 May 2019).
- Portal Antioxidantes. Available online: http://www.portalantioxidantes.com/orac-base-de-datos-actividad-antioxidante-y-contenido-de-polifenoles-totales-en-frutas/ (accessed on 28 May 2019).
- Genskowsky, E.; Puente, L.A.; Pérez-Álvarez, J.A.; Fernández-López, J.; Muñoz, L.A.; Viuda-Martos, M. Determination of polyphenolic profile, antioxidant activity and antibacterial properties of maqui [Aristotelia chilensis (Molina) Stuntz] a Chilean blackberry. J. Sci. Food Agric. 2016, 96, 4235–4242. [Google Scholar] [CrossRef] [PubMed]
- Escribano-Bailón, M.; Alcalde-Eon, C.; Muñoz, O.; Rivas-Gonzalo, J.; Santos-Buelga, C. Anthocyanins in berries of maqui (Aristotelia chilensis (Mol.) Stuntz). Phytochem. Anal. 2006, 17, 8–14. [Google Scholar] [CrossRef]
- Junqueira-Gonçalves, M.P.; Yáñez, L.; Morales, C.; Navarro, M.; AContreras, R.; Zúñiga, G.E. Isolation and characterization of phenolic compounds and anthocyanins from Murta (Ugni molinae Turcz.) fruits. Assessment of antioxidant and antibacterial activity. Molecules 2015, 20, 5698–5713. [Google Scholar] [CrossRef]
- Dávalos, A.; Gómez-Cordovés, C.; Bartolomé, B. Extending applicability of the oxygen radical absorbance capacity (ORAC—fluorescein) assay. J. Agric. Food Chem. 2004, 52, 48–54. [Google Scholar] [CrossRef]
- Fredes, C.; Montenegro, G.; Zoffoli, J.P.; Santander, F.; Robert, P. Comparison of the total phenolic content, total anthocyanin content and antioxidant activity of polyphenol-rich fruits grown in Chile. Cienc. Investig. Agrar. 2014, 41, 49–60. [Google Scholar] [CrossRef]
- Céspedes, C.; El-Hafidi, M.; Pavon, N.; Alarcon, J. Antioxidant and cardioprotective activities of phenolic extracts from fruits of Chilean blackberry Aristotelia chilenesis (Elaeocarpaceae), Maqui. Food Chem. 2008, 108, 820–829. [Google Scholar] [CrossRef]
- Mariangel, E.; Reyes-Diaz, M.; Lobos, W.; Bensch, E.; Schalchli, H.; Ibarra, P. The antioxidant properties of calafate (Berberis microphylla) fruits from four different locations in southern Chile. Ciencia e Investigación Agraria 2013, 40, 161–170. [Google Scholar] [CrossRef]
- González, B.; Vogel, H.; Razmilic, I.; Wolfram, E. Polyphenol, anthocyanin and antioxidant content in different parts of maqui fruits (Aristotelia chilensis) during ripening and conservation treatments after harvest. Ind. Crops Prod. 2015, 76, 158–165. [Google Scholar] [CrossRef]
- Miranda-Rottmann, S.; Aspillaga, A.A.; Pérez, D.D.; Vasquez, L.; Martinez, A.L.F.; Leighton, F. Juice and phenolic fractions of the berry Aristotelia chilensis inhibit LDL oxidation in vitro and protect human endothelial cells against oxidative stress. J. Agric. Food Chem. 2002, 50, 7542–7547. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Valcic, S.; Timmermann, B.N.; Montenegro, G. Indole alkaloids from Aristotelia chilensis (Mol.) Stuntz. Int. J. Pharm. 1997, 35, 215–217. [Google Scholar] [CrossRef]
- Ruiz, A.; Hermosín-Gutiérrez, I.; Vergara, C.; Von Baer, D.; Zapata, M.; Hitschfeld, A.; Obando, L.; Mardones, C. Anthocyanin profiles in south Patagonian wild berries by HPLC-DADESI- MS/MS. Food Res. Int. 2013, 51, 706–713. [Google Scholar] [CrossRef]
- Fredes, C.; Yousef, G.G.; Robert, P.; Grace, M.H.; Lila, M.A.; Gómez, M.; Gebauer, M.; Montenegro, G. Anthocyanin profiling of wild maqui berries (Aristotelia chilensis [Mol.] Stuntz) from different geographical regions in Chile. J. Sci. Food Agric. 2014, 94, 2639–2648. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.E.; Zambrano, R.; Sepúlveda, B.; Kennelly, E.J.; Simirgiotis, M.J. Anthocyanins and antioxidant capacities of six Chilean berries by HPLC-HR-ESI-ToF-MS. Food Chem. 2015, 176, 106–114. [Google Scholar] [CrossRef]
- Mahdavi, S.A.; Jafari, S.M.; Ghorbani, M.; Assadpoor, E. Spray-drying microencapsulation of anthocyanins by natural biopolymers: A review. Drying Technol. 2014, 32, 509–518. [Google Scholar] [CrossRef]
- Gómez-Guillén, M.C.; Ihl, M.; Bifani, V.; Silva, A.; Montero, P. Edible films made from tuna-fish gelatin with antioxidant extracts of two different murta ecotypes leaves (Ugni molinae Turcz.). Food Hydrocoll. 2007, 21, 1133–1143. [Google Scholar] [CrossRef]
- Barbosa-Cánovas, G.V.; Vega-Mercado, H. Dehydration of Foods; Chapman & Hall: New York, NY, USA, 1996. [Google Scholar]
- Doymaz, I. Effect of citric acid and blanching pre-treatments on drying and rehydration of Amasya red apples. Food Bioprod. Process. 2010, 88, 124–132. [Google Scholar] [CrossRef]
- Galaz, P.; Valdenegro, M.; Ramírez, C.; Nuñez, H.; Almonacid, S.; Simpson, R. Effect of drum drying temperature on drying kinetic and polyphenol contents in pomegranate peel. J. Food Eng. 2017, 208, 19–27. [Google Scholar] [CrossRef]
- Valdenegro, M.; Almonacid, S.; Henríquez, C.; Lutz, M.; Fuentes, L.; Simpson, R. The Effects of Drying Processes on Organoleptic Characteristics and the Health Quality of Food Ingredients Obtained from Goldenberry Fruits (Physalis peruviana). Available online: https://www.omicsonline.org/scientific-reports/srep642.php (accessed on 28 May 2019).
- Dev, S.R.S.; Padmini, T.; Adedeji, A.; Garie´py, Y.; Raghavan, G.S.V. A comparative study on the effect of chemical, microwave, and pulsed electric pretreatments on convective drying and quality of raisins. Drying Technol. 2008, 26, 1238–1243. [Google Scholar] [CrossRef]
- Álvarez, C.A.; Aguerre, R.; Gomez, S.; Vidales, S.; Alzamora, S.M.; Gerschenson, L.N. Effect of blanching and glucose dipping pre-treatment on air-drying behavior of strawberries. In Symposium on the Properties of Water. Proceedings of the Poster Session, Universidad de las Américas, México; Argaiz, A., Lopez-Malo, A., Palou, E., Corte, P., Eds.; Universidad de las Americas: Puebla, Mexico, 1994; pp. 11–14. [Google Scholar]
- Alzamora, S.M.; Greschenson, L.M.; Vidales, S.L.; Nieto, A. Structural changes in the minimal processing of fruits; some effects of blanching and sugar impregnation. In Food Engineering 2000; Fito, P., Ortega-Rodriguez, E., Barbosa-Cánovas, G.V., Eds.; Chapman & Hall: New York, NY, USA, 1996; pp. 117–139. [Google Scholar]
- Carranza-Concha, J.; Benlloch, M.; Camacho, M.M.; Martínez-Navarrete, N. Effects of drying and pretreatment on the nutritional and functional quality of raisins. Food Bioprod. Process. 2012, 90, 243–248. [Google Scholar] [CrossRef]
- Cinquanta, L.; Di Matteo, M.; Esti, M. Physical pretreatment of plums (Prunus domestica) Part 2. Effect on the quality characteristics of different prune cultivars. Food Chem. 2002, 79, 233–238. [Google Scholar] [CrossRef]
- Di Matteo, M.; Cinquanta, L.; Galiero, G.; Crescitelli, S. Effect of a novel physical pretreatment process on the drying kinetics of seedless grapes. J. Food Eng. 2000, 46, 83–89. [Google Scholar] [CrossRef]
- Doymaz, I. Influence of pretreatment solution on the drying of sour cherry. J. Food Eng. 2007, 78, 591–596. [Google Scholar] [CrossRef]
- Azoubel, P.M.; Murr, F.E.X. Effect of pretreatment on the drying kinetics of cherry tomato (Lycopersicon esculentum var. cerasiforme). In Transport Phenomena in Food Processing; Welti-Chanes, J., Velez-Ruiz, F., Barbosa-Cánovas, G.V., Eds.; CRC Press: New York, NY, USA, 2003; pp. 137–151. [Google Scholar]
- Feng, H.; Tang, J.; Mattinson, D.S.; Fellman, J.K. Microwave and spouted bed drying of frozen blueberries: The effect of drying and pre-treatment methods on physical properties and retention of flavour volatiles. J. Food Process. Preserv. 1999, 23, 463–479. [Google Scholar] [CrossRef]
- Grabowski, S.; Marcotte, M. Pretreatment efficiency in osmotic dehydration of cranberries. In Transport Phenomena in Food Processing; Welti-Chanes, J., Velez-Ruiz, F., Barbosa-Cánovas, G.V., Eds.; CRC Press: New York, NY, USA, 2003; pp. 83–94. [Google Scholar]
- Grabowski, S.; Marcotte, M.; Poirier, M.; Kudra, T. Drying Characteristics of Osmotically Pretreated Cranberries: Energy and Quality Aspects. 2002. Available online: http://cetcvarennes.nrcan.gc.ca/eng/publication/r2002083e.html (accessed on 12 June 2003).
- Yang, P.A.P.; Wills, C.; Yang, C.S.T. Use of a combination process of osmotic dehydration and freeze drying to produce a raisin-type lowbush blueberry product. J. Food Sci. 1987, 52, 1651–1653. [Google Scholar] [CrossRef]
- George, S.T.; Cenkowski, S.; Muir, W.E. A review of drying technologies for the preservation of nutritional compounds in waxy skinned fruit. In Proceedings of the 2004 North Central ASAE/CSAE Conference, Winnipeg, MB, Canada, 24–25 September 2004. [Google Scholar]
- Desai, K.; Park, H. Recent developments in microencapsulation of food ingredients. Drying Technol. 2005, 23, 1361–1394. [Google Scholar] [CrossRef]
- Robert, P.; García, P.; Fredes, C. Drying and preservation of polyphenols. In Advances in Technologies for Producing Food-Relevant Polyphenols; Cuevas-Valenzuela, J., Vergara Salinas, J.R., Pérez-Correa, J.R., Eds.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2017; pp. 281–302. [Google Scholar]
- Fredes, C.; Becerra, C.; Parada, J.; Robert, P. The Microencapsulation of Maqui (Aristotelia chilensis (Mol.) Stuntz) Juice by Spray-Drying and Freeze-Drying Produces Powders with Similar Anthocyanin Stability and Bioaccessibility. Molecules 2018, 23, 1227. [Google Scholar] [CrossRef]
- Parra, H. Revision: Food microencapsulation. Mag. Natl. Fac. Agron. 2010, 63, 5669–5684. [Google Scholar]
- Laokuldilok, T.; Kanha, N. Effects of processing conditions on powder properties of black glutinous rice (Oryza sativa L.) bran anthocyanins produced by spray drying and freeze drying. LWT Food Sci. Technol. 2015, 64, 405–411. [Google Scholar] [CrossRef]
- Ah-Hen, K.S.; Zambra, C.E.; Aguëro, J.E.; Vega-Gálvez, A.; Lemus-Mondaca, R. Moisture diffusivity coefficient and convective drying modelling of murta (Ugni molinae Turcz.): Influence of temperature and vacuum on drying kinetics. Food Bioprocess. Technol. 2013, 6, 919–930. [Google Scholar] [CrossRef]
- Rodríguez, K.; Ah-Hen, K.; Vega-Gálvez, A.; López, J.; Quispe-Fuentes, I.; Lemus-Mondaca, R.; Galvez-Ranilla, L. Changes in bioactive compounds and antioxidant activity during convective drying of murta (Ugni molinae T.) berries. Int. J. Food Sci. Technol. 2014, 49, 990–1000. [Google Scholar] [CrossRef]
- Alfaro, S.; Mutis, A.; Quiroz, A.; Seguel, I.; Scheuermann, E. Effects of drying techniques on murtilla fruit polyphenols and antioxidant activity. J. Food Res. 2014, 3, 73–82. [Google Scholar] [CrossRef]
- Bastias-Montes, J.M.; Alarcón-Enos, J.; Quevedo-León, R.; Muñoz-Fariña, O.; Vidal-San-Martín, C. Effect of spray drying at 150, 160, and 170 °C on the physical and chemical properties of maqui extract (Aristotelia chilensis (Molina) Stuntz). Chil. J. Agric. Res. 2019, 79, 144–152. [Google Scholar] [CrossRef]
- Pasin, B.; González, C.; Maestro, A. Microencapsulación con alginato en alimentos. Técnicas y aplicaciones. Cienc Tecnol Alimentos 2012, 3, 130–151. [Google Scholar]
- Dubey, R.; Shami, T.C.; Bhasker Rao, K.U. Microencapsulation Technology and Applications. Def. Sci. J. 2009, 59, 82–95. [Google Scholar]
- Saénz, C.; Tapia, S.; Chávez, J.; Robert, P. Microencapsulation by spray drying of bioactive compounds from cactus pear (Opuntia ficus-indica). Food Chem. 2009, 114, 616–622. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Pasli, A.A.; Ozcelik, B.; Capanoglu, E. Evaluating the in vitro bioaccessibility of phenolics and antioxidant activity during consumption of dried fruits with nuts. LWT Food Sci. Technol. 2014, 56, 284–289. [Google Scholar] [CrossRef]
- Mandalari, G.; Bisignano, C.; Filocamo, A.; Chessa, S.; Sarò, M.; Torre, G.; Dugo, P. Bioaccessibility of pistachio (Pistacia vera L.) polyphenols, xanthophylls and tocopherols during simulated human digestion. Nutrition 2013, 29, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Fredes, C.; Osorio, M.J.; Parada, J.; Robert, P. Stability and bioaccessibility of anthocyanins from maqui (Aristotelia chilensis [Mol.] Stuntz) juice microparticles. LWT Food Sci. Technol. 2018, 91, 549–556. [Google Scholar] [CrossRef]
- Ah-Hen, K.S.; Mathias-Rettig, K.; Gómez-Pérez, L.S.; Riquelme-Asenjo, G.; Lemus-Mondaca, R.; Muñoz-Fariña, O. Bioaccessibility of bioactive compounds and antioxidant activity in murta (Ugni molinae T.) berries juices. Food Meas. 2018, 12, 602. [Google Scholar] [CrossRef]
- Stevenson, D.E.; Hurst, R.D. Polyphenolic phytochemicals—Just antioxidants or much more? Cell Mol. Life Sci. 2007, 64, 2900–2916. [Google Scholar] [CrossRef] [PubMed]
- Amiot, M.J.; Riva, C.A.V. Effects of dietary polyphenols on metabolic syndrome features in humans: A systematic review. Obes. Rev. 2016, 17, 573–586. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef]
- D’Archivio, M.; Filesi, C.; Varì, R.; Scazzocchio, B.; Masella, R. Bioavailability of the polyphenols: Status and controversies. Int. J. Mol. Sci. 2010, 11, 1321–1342. [Google Scholar] [CrossRef]
- Rubió, L.; Macià, A.; Motilva, M.J. Impact of various factors on pharmacokinetics of bioactive polyphenols: An overview. Curr. Drug Metab. 2014, 15, 62–76. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, D.; Sun, J.; Liu, X.; Jiang, L.; Guo, H.; Ren, F. Interaction of plant phenols with food macronutrients: Characterisation and nutritional-physiological consequences. Nutr. Res. Rev. 2014, 27, 1–15. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81 (Suppl. 1), 230S–242S. [Google Scholar] [CrossRef]
- Salzanoa, S.; Checconia, P.; Hanschmannc, E.M.; Lillig, C.H.; Bowler, L.D.; Chan, P.; Vaudry, D.; Mengozzi, M.; Coppo, L.; Sacre, S.; et al. Linkage of inflammation and oxidative stress via release of glutathionylated peroxiredoxin-2, which acts as a danger signal. Proc. Natl. Acad. Sci. USA 2014, 111, 12157–12162. [Google Scholar] [CrossRef]
- Kim, H.Y.; Nam, S.Y.; Hong, S.W.; Kim, M.J.; Jeong, H.J.; Kim, H.M. Protective effects of rutin through regulation of vascular endothelial growth factor in allergic rhinitis. Am. J. Alle. 2015, 29, 87–94. [Google Scholar] [CrossRef]
- Chen, W.Y.; Huang, Y.C.; Yang, M.L.; Lee, C.Y.; Chen, C.J.; Yeh, C.H.; Pan, P.H.; Horng, C.T.; Kuo, W.H.; Kuan, Y.H. Protective effect of rutin on LPS-induced acute lung injury via down-regulation of MIP-2 expression and MMP-9 activation through inhibition of Akt phosphorylation. Int. Immunopharmacol. 2014, 22, 409–413. [Google Scholar] [CrossRef]
- Qin, M.; Chen, W.; Cui, J.; Li, W.; Liu, D.; Zhang, W. Protective efficacy of inhaled quercetin for radiation pneumonitis. Exp. Ther. Med. 2017, 14, 5773–5778. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schreckinger, M.E.; Wang, J.; Yousef, G.; Lila, M.A.; Gonzalez de Mejia, E. Antioxidant capacity and in vitro inhibition of adipogenesis and inflammation by phenolic extracts of Vaccinium floribundum and Aristotelia chilensis. J. Agric. Food Chem. 2010, 58, 8966–8976. [Google Scholar] [CrossRef] [PubMed]
- Paredes-López, O.; Cervantes-Ceja, M.L.; Vigna-Pérez, M.; Hernández-Pérez, T. Berries: Improving human health and healthy aging, and promoting quality life A review. Plant Foods Hum. Nutr. 2010, 65, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Cespedes, C.L.; Pavon, N.; Dominguez, M.; Alarcon, J.; Balbontin, C.; Kubo, I.; El-Hafidi, M.; Avila, J.G. The chilean superfruit black-berry Aristotelia chilensis (Elaeocarpaceae), Maqui as mediator in inflammation-associated disorders. Food Chem. Toxicol. 2017, 108, 438–450. [Google Scholar] [CrossRef]
- Moon, H.D.; Kim, B.H. Inhibitory effects of Aristotelia chilensis water extract on 2,4-Dinitrochlorobenzene induced atopic-like dermatitis in BALB/c Mice. Asian Pac. J. Allr. Immunol. 2019. [Google Scholar] [CrossRef]
- Vergara, D.; Ávila, D.; Escobar, E.; Carrasco-Pozo, C.; Sánchez, A.; Gotteland, M. The intake of maqui (Aristotelia chilensis) berry extract normalizes H2O2 and IL-6 concentrations in exhaled breath condensate from healthy smokers—an explorative study. Nutr. J. 2015, 14, 27. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, M.C.; Delporte, C.; Backhouse, N.; Erazo, S.; Letelier, M.E.; Cassels, B.K.; Silva, X.; Alegría, S.; Negrete, R. Topical anti-inflammatory activity of 2a-hydroxy pentacyclic triterpene acids from the leaves of Ugni molinae. Bioorg. Med. Chem. 2006, 14, 5673–5677. [Google Scholar] [CrossRef]
- Arancibia-Radich, J.; Peña-Cerda, M.; Jara, D.; Valenzuela-Bustamante, P.; Goity, L.; Valenzuela-Barra, G.; Silva, X.; Garrido, G.; Delporte, C.; Seguel, I. Comparative study of anti-inflammatory activity and qualitative-quantitative composition of triterpenoids from ten genotypes of Ugni molinae. Bol. Latinoam. Caribe Plantas Med. Aromát. 2016, 15, 274–287. [Google Scholar]
- López, J.; Vega-Gálvez, A.; Rodríguez, A.; Uribe, E.; Bilbao-Sainz, C. Murta (Ugni molinae turcz.): A review on chemical composition, functional components and biological activities of leaves and fruits. Chil. J. Agric. Anim. Sci. 2018, 34, 1–14. [Google Scholar]
- Rubilar, M.; Jara, C.; Poo, Y.; Acevedo, F.; Gutierrez, C.; Sineiro, J.; Shene, C. Extracts of maqui (Aristotelia chilensis) and murta (Ugni molinae Turcz.): Sources of antioxidant compounds and r-glucosidase/r-amylase inhibitors. J. Agric. Food Chem. 2011, 59, 1630–1637. [Google Scholar] [CrossRef] [PubMed]
- Rojo, L.E.; Ribnicky, D.; Logendra, S.; Poulev, A.; Rojas-Silva, P.; Kuhn, P.; Dorn, R.; Grace, M.H.; Lila, M.A.; Raskin, I. In Vitro and in Vivo Anti-Diabetic Effects of Anthocyanins from Maqui Berry (Aristotelia chilensis). Food Chem. 2012, 131, 387–396. [Google Scholar] [CrossRef]
- Schön, C.; Wacker, R.; Micka, A.; Steudle, J.; Lang, S.; Bonnländer, B. Bioavailability Study of Maqui Berry Extract in Healthy Subjects. Nutrients 2018, 10, 1720. [Google Scholar] [CrossRef] [PubMed]
- Vegt, D.F.; Dekker, M.J.; Ruhe, G.H.; Stehouwer, D.C.; Nijpels, G.; Bouter, M.L.; Heine, J.R. Hyperglycemia is associated with all-cause and cardiovascular mortality in the Hoorn population: The Hoorn study. Diabetologia 1999, 42, 926–931. [Google Scholar] [CrossRef]
- Durand, M.J.; Gutterman, D. Diversity in mechanisms of endothelium-dependent vasodilation in health and disease. Microcirculation 2013, 20, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Jofré, I.; Pezoa, C.; Cuevas, M.; Scheuermann, E.; Freires, I.A.; Rosalen, P.L.; de Alencar, S.M.; Romero, F. Antioxidant and Vasodilator Activity of Ugni molinae Turcz. (Murtilla) and Its Modulatory Mechanism in Hypotensive Response. Oxid. Med. Cell. Longev. 2016, 2016, 6513416. [Google Scholar] [CrossRef]
- O’Keefe, J.H.; Bell, D.S. Postprandial hyperglycemia/hyperlipidemia (postprandial dysmetabolism) is a cardiovascular risk factor. Am. J. Cardiol. 2007, 100, 899–904. [Google Scholar] [CrossRef] [PubMed]
- Cifuentes, F.; Palacios, J.; Paredes, A.; Nwokocha, C.R.; Paz, C. 8-Oxo-9-Dihydromakomakine Isolated from Aristotelia chilensis Induces Vasodilation in Rat Aorta: Role of the Extracellular Calcium Influx. Molecules 2018, 23, 3050. [Google Scholar] [CrossRef] [PubMed]
- Davinelli, S.; Bertoglio, J.C.; Zarrelli, A.; Pina, R.; Scapagnini, G. A Randomized Clinical Trial Evaluating the Efficacy of an Anthocyanin-Maqui Berry Extract (Delphinol®) on Oxidative Stress Biomarkers. J. Am. Coll. Nutr. 2015, 34 (Suppl. 1), 28–33. [Google Scholar] [CrossRef] [PubMed]
- Cenativ. Available online: http://cenativ.utalca.cl/noticias/nuevas-variedades-de-maqui-abren-opcion-para-cultivo-del-fruto/ (accessed on 28 May 2019).
- ODEPA—Oficina de Estudios y Políticas Agrarias. Boletín de Frutas y Hortalizas Procesadas. Available online: https://www.odepa.gob.cl/wp-content/uploads/2019/03/BFrutaprocesada0319.pdf (accessed on 13 May 2019).
- Infor. Available online: http://www.infor.gob.cl/ (accessed on 26 May 2019).
- Seguel, I.; Peñaloza, E.; Gaete, N. Colecta y caracterización molecular de germoplasma de murta (Ugni molinae Turcz.). en Chile. Agro Sur 2000, 28, 32–41. [Google Scholar] [CrossRef]
- Alfaro, S.; Mutis, A.; Palma, R.; Quiroz, A.; Seguel, I.; Scheuermann, E. Influence of genotype and harvest year on polyphenol content and antioxidant activity in murtilla (Ugni molinae Turcz.) fruit. J. Soil Sci. Plant Nutr. 2013, 13, 67–78. [Google Scholar] [CrossRef]
- EU. Available online: http://ec.europa.eu/food/safety/novel_food/catalogue/search/public/index.cfm# (accessed on 16 May 2019).
- Slow Food Foundation for Biodiversity. Ark of Taste (Chile): “Purén White Strawberry”. 2015. Available online: http://www.fondazioneslowfood.com/en/slow-food-presidia/puren-white-strawberry/ (accessed on 25 July 2015).
- FIA. Available online: http://www.fia.cl/inia-y-fia-lanzan-polo-territorial-para-fortalecer-industria-de-colorantes-naturales/ (accessed on 26 May 2019).




{kind=link}
{kind=link}
{kind=link}
| Species | Common Name | Family | Geographic Distribution [16,35] | Traditional Products and Uses | Functional Products |
|---|---|---|---|---|---|
| Aristotelia chilensis (Mol.) Stuntz. | Maqui | Elaeocarpaceae | Chile: from the Coquimbo to Aysén regions, including Juan Fernández Island (Latitude 31°–40°). Argentina: from Jujuy to Chubut provinces. | Fresh and dried fruit, use to make textile pigment, cake, jam, juice, alcoholic beverages [36,37] | Freeze-dried maqui (powder and capsules), honey mix, functional drinks, drugs [24,25,26,38,39,40,41] |
| Ugni molinae Turcz. | Murta | Myrtaceae | Chile: From the O’Higgins to Aysén regions, including Juan Fernández Island (Lat. 34°–40°). Argentina: Neuquén, Rio Negro, and Chubut provinces. | Fresh and dried fruit, textile pigment, bakery, jam, alcoholic beverages [37] | Freeze-dried murta (powder and capsules), honey mix [41,42] |
| Berberis microphylla G. Forst. | Calafate | Berberidaceae | Chile: From the Metropolitan to Magallanes regions (Lat. 33°–55°). Argentina: From Neuquén to Tierra del Fuego provinces. | Fresh fruit, used to make jam, juice, beer [36,37] | Natural colorants [37] |
| Luma apiculata (DC.) Burret. | Arrayán | Myrtaceae | Chile: From the Coquimbo to Aysén regions (Lat. 31°–40°). Argentina: From Neuquén to Chubut provinces. | Fresh fruit, textile pigment, bakery, jam, aromatic wine [22,23] | N.D. |
| Fragaria chiloensis (L.) Mill. | Chilean strawberry | Rosaceae | Chile: From the O’Higgins to Magallanes regions (Lat. 34°–55°). Argentina: Neuquén and Rio Negro provinces. | Fresh fruit, used to make alcoholic beverages, cake [36,43] | N.D. |
| Species Name | Average Antioxidant Capacity Determined by ORAC (µmol·100 g DW−1) a | Average Range of Total Polyphenols Compounds Content (mg GAE g−1 DW−1) a | Number of Non-Anthocyanin Polyphenol Compounds Reported | Principal Non-Anthocyanin Polyphenol Compounds | Number of Anthocyanin Compound Reported | Principal Anthocyanin Compounds |
|---|---|---|---|---|---|---|
| Maqui. | 37,174 [11,69] | 49.7 [70] | 13 [15] | Quercetin, dimethoxy-quercetin, quercetin-3-rutinoside, quercetin-3-galactoside, myricetin and its derivatives (dimethoxy-quercetin) and ellagic acid [70] | 8 [15] | 3-glucosides, 3,5-diglucosides, 3-sambubiosides and 3-sambubioside-5-glucosides of cyanidin and delphinidin (delphinidin 3-sambubioside-5-glucoside) [20,71] |
| Murta | 43,574 [11,69] | 9.2 [19] 34.9 [69] | 16 [15] | caffeic acid-3-glucoside, quercetin-3-glucoside, quercetin, gallic acid, quercetin-3-rutinoside, quercitrin, luteolin, luteolin-3-glucoside, kaempferol, kaempferol-3-glucoside, myricetin and p-coumaric acid [72] | 11 [15] | delphinidin-3-, malvidin-3- and peonidin-3-arabinoside; peonidin-3- and malvidin-3-glucoside [20,72] |
| Calafate | 72,425 [11,69] | 33.9 [69] 65.5 [19] | 36 [15] | quercetin-3-rutinoside, gallic- and chlorogenic acid, caffeic and the presence of coumaric- and ferulic acid, quercetin, myricetin, and kaempferol [19] | 30 [15] | delphinidin-3-glucoside, delphinidin-3-rutinoside, delphinidin-3,5-dihexoside, cyanidin-3-glucoside, petunidin-3-glucoside, petunidin-3-rutinoside, petunidin-3,5-dihexoside, malvidin-3-glucoside and malvidin-3-rutinoside [19,20] |
| Arrayán | 62,500 [21] | 27.6 [19] | 13 [15] | quercetin 3-rutinoside and their derivatives, tannins and their monomers [18,21] | 8 [15] | peonidin-3-galactoside, petunidin-3-arabinoside, malvidin-3-arabinoside, peonidin-3-arabinoside delphinidin-3-arabinoside, cyanidin-3-glucoside, peonidin-3-glucoside and malvidin-3-glucoside [18,19,21] |
| Chilean strawberry | N.R. | N.R | 16*20** [17] | ellagic acid and their pentoside- and rhamnoside derivatives. quercetin glucuronide, ellagitannin, quercetin pentoside, kaempferol glucuronide. Catechin *, quercetin pentosid *, and quercetin hexoside * procyanidin tetramers ** and ellagitannin ** [17] | 4 [17] | cyanidin 3-O-glucoside, pelargonidin 3-O-glucoside, cyanidin-malonyl-glucoside and pelargonidin-malonyl- glucoside [17] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuentes, L.; Figueroa, C.R.; Valdenegro, M.; Vinet, R. Patagonian Berries: Healthy Potential and the Path to Becoming Functional Foods. Foods 2019, 8, 289. https://doi.org/10.3390/foods8080289
Fuentes L, Figueroa CR, Valdenegro M, Vinet R. Patagonian Berries: Healthy Potential and the Path to Becoming Functional Foods. Foods. 2019; 8(8):289. https://doi.org/10.3390/foods8080289
Chicago/Turabian StyleFuentes, Lida, Carlos R. Figueroa, Monika Valdenegro, and Raúl Vinet. 2019. "Patagonian Berries: Healthy Potential and the Path to Becoming Functional Foods" Foods 8, no. 8: 289. https://doi.org/10.3390/foods8080289
APA StyleFuentes, L., Figueroa, C. R., Valdenegro, M., & Vinet, R. (2019). Patagonian Berries: Healthy Potential and the Path to Becoming Functional Foods. Foods, 8(8), 289. https://doi.org/10.3390/foods8080289