Evaluation of Biomass and Chitin Production of Morchella Mushrooms Grown on Starch-Based Substrates

 ,
, 


Abstract
1. Introduction
2. Materials and Methods
2.1. Raw Materials
2.2. Morchella Strains, Media, and Culture Conditions
2.3. Submerged Fermentations on Commercial Substrates
2.4. Solid-State Fermentations on Commercial Substrates
2.5. Solid-State Fermentations on Agro-Industrial Substrates
2.6. Analytical Methods
2.6.1. Mycelium Growth Rate
2.6.2. Determination of Biomass and Sugar Consumption in Submerged Fermentations
2.6.3. Determination of Biomass in Solid-State Fermentations
2.6.4. Determination of Total Polysaccharide Content
3. Results and Discussion
3.1. Submerged Fermentations
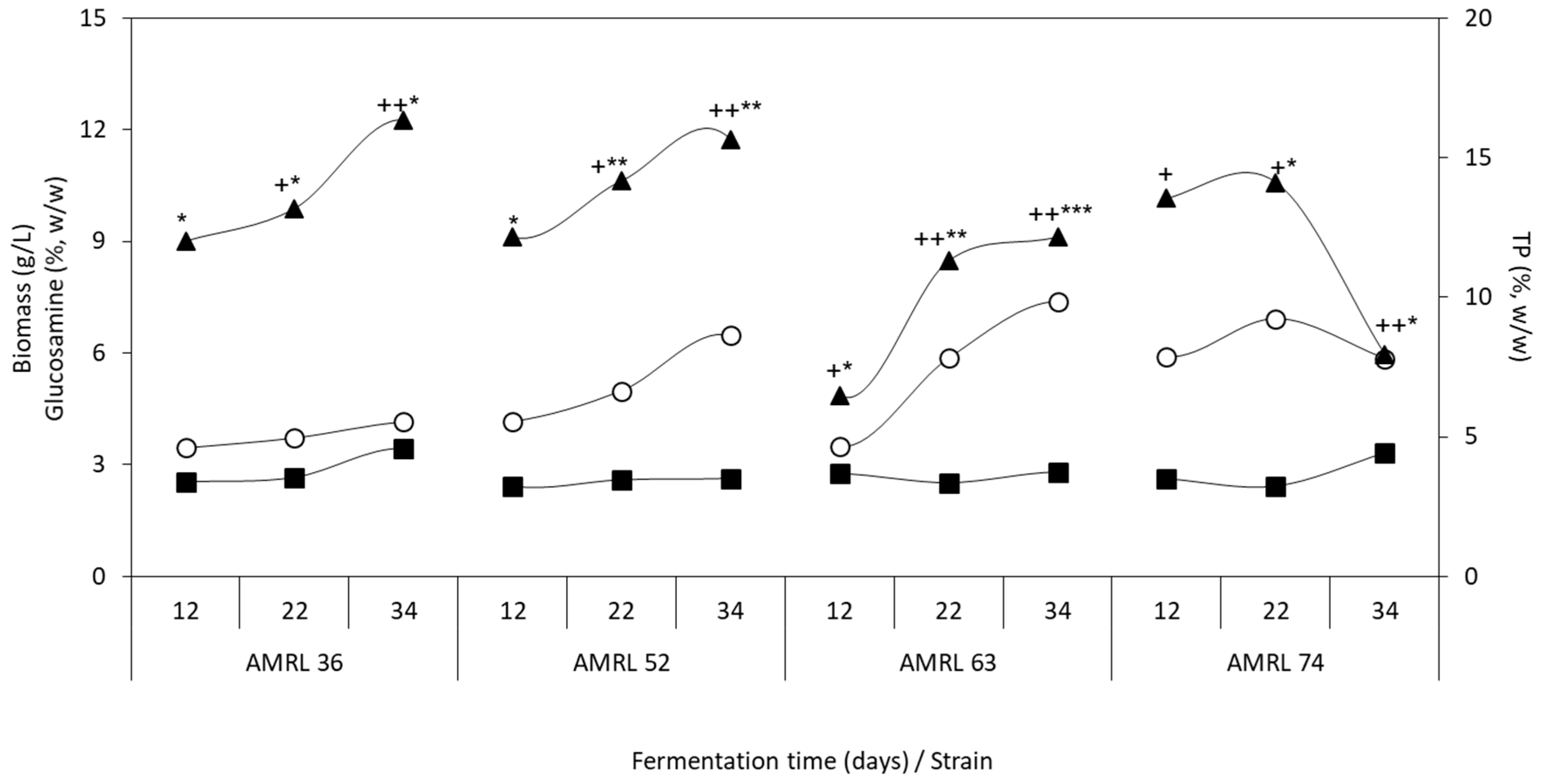
3.1.1. Biomass Production, Glucosamine, and Total Polysaccharide Content
3.2. Solid-State Fermentations on Commercial Substrates
3.2.1. Mycelial Growth Rate
3.2.2. Sclerotia Formation
3.2.3. Biomass Production, Glucosamine, and Total Polysaccharide Content
3.3. Solid-State Fermentations on Agro-Industrial Substrates
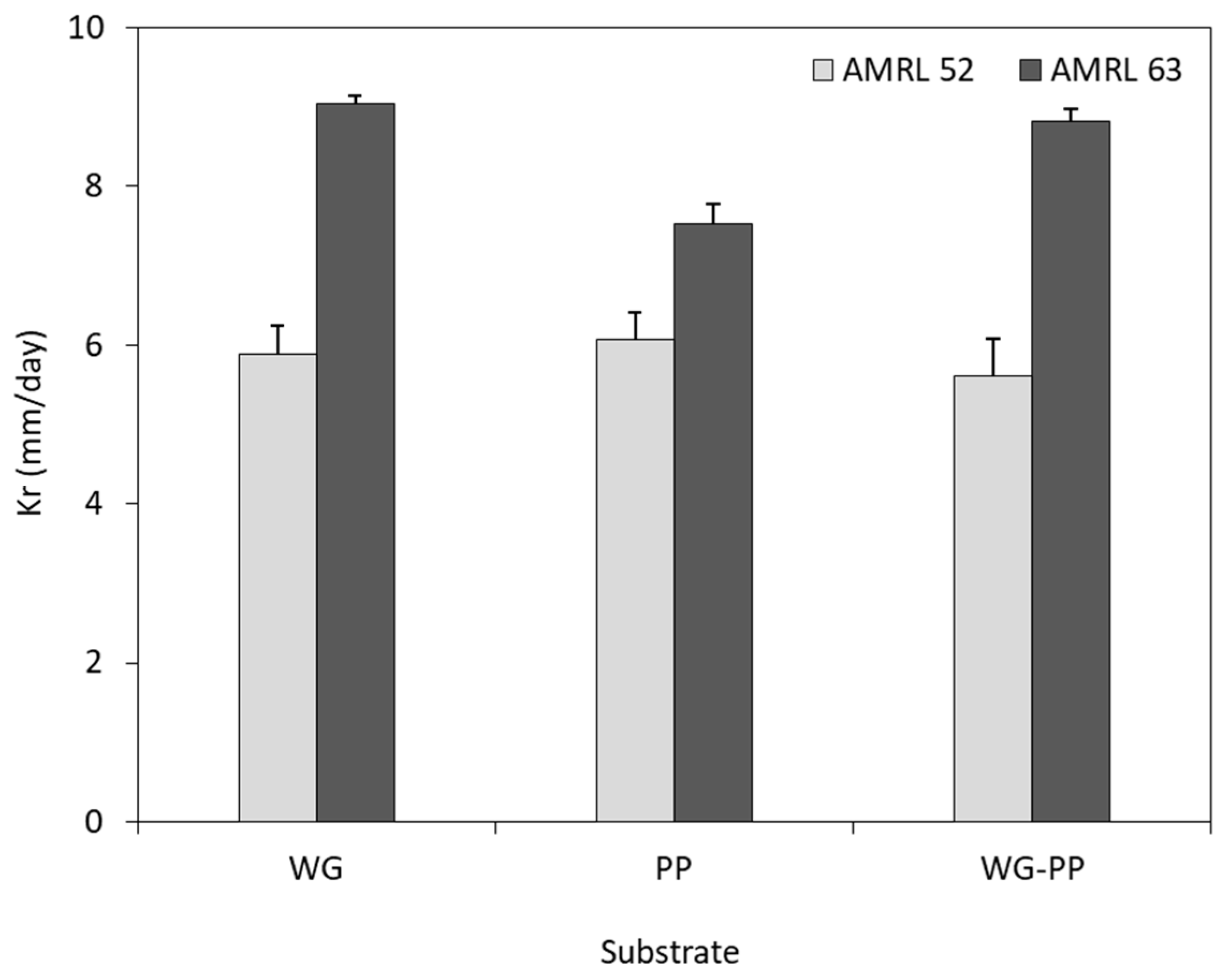
3.3.1. Mycelial Growth Rate
3.3.2. Sclerotia Formation
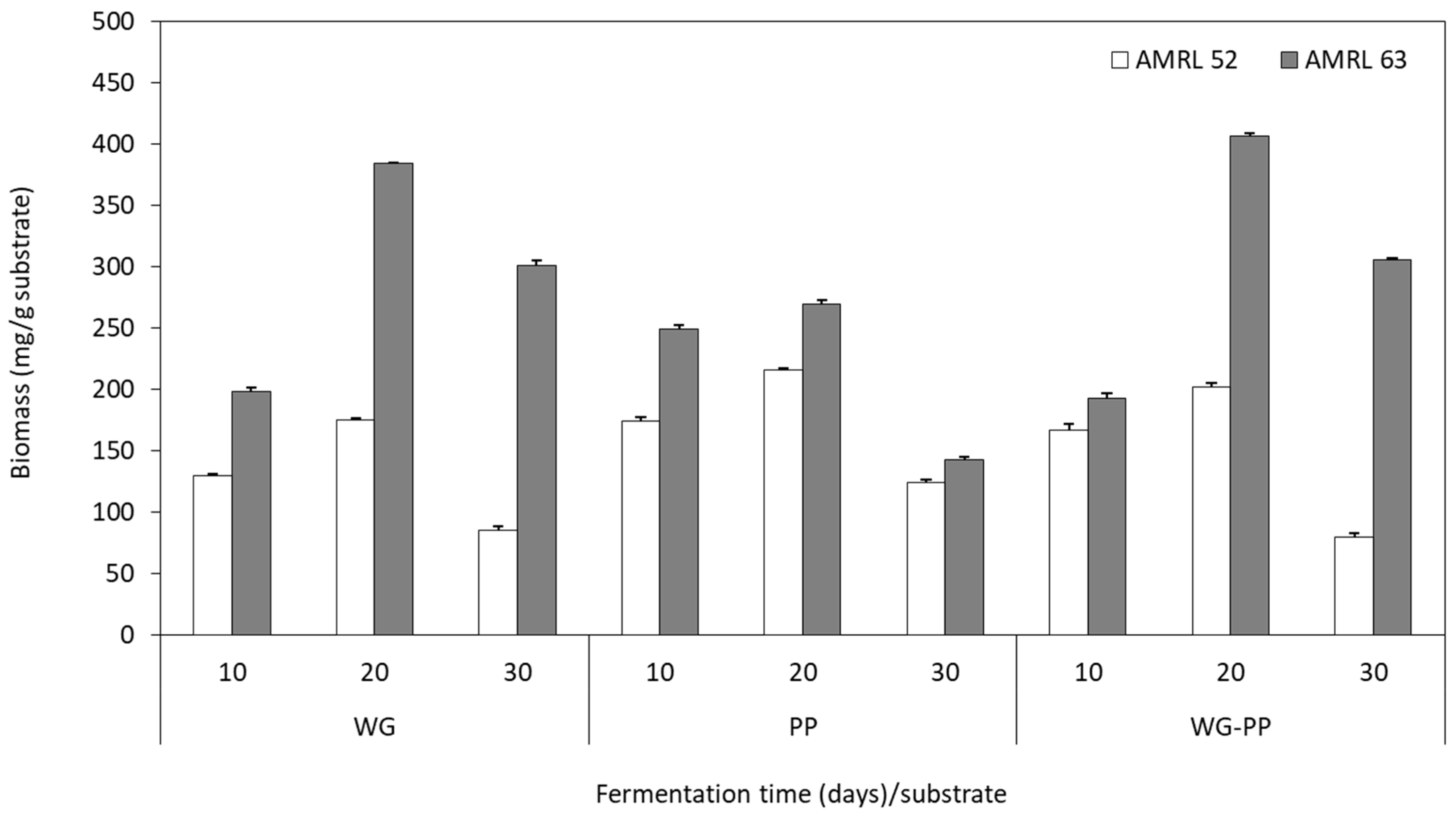
3.3.3. Biomass Production and Total Polysaccharide Content
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Diamantopoulou, P.; Philippoussis, A. Cultivated Mushrooms: Preservation and Processing. In Handbook of Vegetable Preservation and Processing, 2nd ed.; Hui, Y.H., Evranuz, E.Ö., Bingöl, G., Erten, H., Jaramillo-Flores, M.E., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 495–525. [Google Scholar]
- Carvajal, A.E.S.S.; Koehnlein, E.A.; Soares, A.A.; Eler, G.J.; Nakashima, A.T.A.; Bracht, A.; Peralta, R.M. Bioactives of fruiting bodies and submerged culture mycelia of Agaricus brasiliensis (A. blazei) and their antioxidant properties. LWT-Food Sci. Technol. 2012, 46, 493–499. [Google Scholar] [CrossRef]
- Economou, C.N.; Diamantopoulou, P.A.; Philippoussis, A.N. Valorization of spent oyster mushroom substrate and laccase recovery through successive solid state cultivation of Pleurotus, Ganoderma, and Lentinula strains. Appl. Microbiol. Biotechnol. 2017, 101, 5213–5222. [Google Scholar] [CrossRef] [PubMed]
- Philippoussis, A.; Diamantopoulou, P.; Papadopoulou, K.; Lakhtar, H.; Roussos, S.; Parissopoulos, G.; Papanikolaou, S. Biomass, laccase and endoglucanase production by Lentinula edodes during solid state fermentation of reed grass, bean stalks and wheat straw residues. World J. Microbiol. Biotechnol. 2011, 27, 285–297. [Google Scholar] [CrossRef]
- Philippoussis, A.; Zervakis, G.; Diamantopoulou, P. Bioconversion of lignocellulosic wastes through the cultivation of the edible mushrooms Agrocybe aegerita, Volvariella volvacea and Pleurotus spp. World J. Microbiol. Biotechnol. 2001, 17, 191–200. [Google Scholar] [CrossRef]
- Choong, Y.-K.; Ellan, K.; Chen, X.-D.; Mohamad, S.A. Extraction and Fractionation of Polysaccharides from a Selected Mushroom Species, Ganoderma lucidum: A Critical Review, Fractionation, Hassan Al- Haj Ibrahim. IntechOpen, 2018; pp. 39–60. Available online: https://www.intechopen.com/books/fractionation/extraction-and-fractionation-of-polysaccharides-from-a-selected-mushroom-species-ganoderma-lucidum-a (accessed on 2 June 2019). [CrossRef]
- Hamed, I.; Özogul, F.; Regenstein, J.M. Industrial applications of crustacean by-products (chitin, chitosan, and chitooligosaccharides): A review. Trend Food Sci. Technol. 2016, 48, 40–50. [Google Scholar] [CrossRef]
- Sitanggang, A.B.; Sophia, L.; Wu, H.S. Aspects of glucosamine production using microorganisms. Int. Food Res. J. 2012, 19, 393–404. [Google Scholar]
- Ride, J.P.; Drysdale, R.B. A chemical method for estimating Fusarium oxysporum f. lycopersici in infected tomato plants. Physiol. Plant Pathol. 1971, 1, 409–420. [Google Scholar] [CrossRef]
- Zdarta, J.; Meyer, A.S.; Jesionowski, T.; Pinelo, M. A General Overview of Support Materials for Enzyme Immobilization: Characteristics, Properties, Practical Utility. Catalysts 2018, 8, 92. [Google Scholar] [CrossRef]
- Tan, C.; Feng, B.; Zhang, X.; Xia, W.; Xia, S. Biopolymer-coated liposomes by electrostatic adsorption of chitosan (chitosomes) as novel delivery systems for carotenoids. Food Hydrocoll 2016, 52, 774–784. [Google Scholar] [CrossRef]
- Cheung, R.C.F.; Ng, T.B.; Wong, J.H.; Chan, W.Y. Chitosan: An Update on Potential Biomedical and Pharmaceutical Applications. Mar. Drugs 2015, 13, 5156–5186. [Google Scholar] [CrossRef]
- A Worldwide Market with a Strong Demand. Available online: http://sflyproteins.com/a-worldwide-market-with-a-strong-demand/ (accessed on 2 June 2019).
- Wu, T.; Zivanovic, S.; Draughon, F.A.; Sams, C.E. Chitin and Chitosan Value-Added Products from Mushroom Waste. J. Agric. Food Chem. 2004, 52, 7905–7910. [Google Scholar] [CrossRef] [PubMed]
- Di Lena, G.; Annibate, A.D.; Sermanni, G.G. Influence of the age and growth conditions on the mycelial chitin content of Lentinus edodes. J. Basic Microbiol. 1994, 34, 11–16. [Google Scholar] [CrossRef]
- Prasad, P.; Chauhan, K.; Kandari, L.S.; Maikhuri, R.K.; Purohit, A.; Bhatt, R.P.; Rao, K.S. Morchella esculenta (Guchhi): Need for scientific intervention for its cultivation in Central Himalaya. Curr. Sci. 2002, 82, 1098–1100. [Google Scholar]
- Volk, T.J.; Leonard, T.J. Cytology of the life-cycle of Morchella. Mycol. Res. 1990, 94, 399–406. [Google Scholar] [CrossRef]
- Winder, R.S. Cultural studies of Morchella elata. Mycol. Res. 2006, 110, 612–623. [Google Scholar] [CrossRef] [PubMed]
- Güller, P.; Arkan, O. Cultural Characteristics of Morchella esculenta Mycelium on Some Nutrients. Turk. J. Biol. 2000, 24, 783–794. [Google Scholar]
- Xu, N.; Lu, Y.; Hou, J.; Liu, C.; Sun, Y. A Polysaccharide Purified from Morchella conica Pers. Prevents Oxidative Stress Induced by H2O2 in Human Embryonic Kidney (HEK) 293T Cells. Int. J. Mol. Sci. 2018, 19, 4027. [Google Scholar] [CrossRef]
- Yang, C.; Zhou, X.; Meng, Q.; Wang, Μ.; Zhang, Y.; Fu, S. Secondary Metabolites and Antiradical Activity of Liquid Fermentation of Morchella sp. Isolated from Southwest China. Molecules 2019, 24, 1706. [Google Scholar] [CrossRef]
- Liu, C.; Sun, Y.; Mao, Q.; Guo, X.; Li, P.; Liu, Y.; Xu, N. Characteristics and Antitumor Activity of Morchella esculenta Polysaccharide Extracted by Pulsed Electric Field. Int. J. Mol. Sci. 2016, 17, 986. [Google Scholar] [CrossRef]
- Lau, B.F.; Abdullah, N. Sclerotium-Forming Mushrooms as an Emerging Source of Medicinals: Current perspectives. In Mushroom Biotechnology, Developments and Applications; Petre, M., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 111–136. [Google Scholar]
- Gilbert, F. The submerged culture of Morchella. Mycologia 1960, 52, 201–209. [Google Scholar] [CrossRef]
- Kaul, T.N. Physiological studies on Morchella spp. I. Carbon utilization. Bull. Bot. Soc. Bengal. 1978, 31, 35–42. [Google Scholar]
- Kosaric, N.; Miyata, N. Growth of morel mushroom mycelium in cheese whey. J. Dairy Res. 1981, 48, 149–162. [Google Scholar] [CrossRef]
- Philippoussis, A.; Balis, C. Studies on the morphogenesis of sclerotia and subterranean mycelial network of ascocarps in “Morchella” species. In Science and Cultivation of Edible Fungi, Proceedings of the 14th international congress on the science and cultivation of edible fungi, Oxford, England; Elliott, T.J., Ed.; A.A. Balkema: Rotterdam, the Netherlands, 1995; pp. 847–855. [Google Scholar]
- Ang, T.N.; Ngoh, G.C.; Chua, A.S.M. Development of a novel inoculum preparation method for solid-state fermentation-Cellophane film culture (CFC) technique. Ind. Crop. Prod. 2013, 43, 774–777. [Google Scholar] [CrossRef]
- Araujo, A.A.; Roussos, S. A technique for mycelial development of ectomycorrhizal fungi on agar media. Appl. Biochem. Biotechnol. 2002, 98, 311–318. [Google Scholar] [CrossRef]
- Robson, G.D.; Bell, S.D.; Kuhn, P.J.; Trinci, A.P.J. Glucose and penicillin concentrations in agar medium below fungal colonies. J. Gen. Microbiol. 1987, 133, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Dantigny, P.; Guilmart, A.; Bensoussan, M. Basis of predictive mycology. Int. J. Food Microbiol. 2005, 100, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Baranyi, J.; Roberts, T.A.; McClure, P. A non-autonomous differential equation to model bacterial growth. Food Microbiol. 1993, 10, 43–59. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugars. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Scotti, C.T.; Vergoignan, C.; Feron, G.; Durand, A. Glucosamine measurement as indirect method for biomass estimation of Cunninghamella elegans grown in solid state cultivation conditions. Biochem. Eng. J. 2001, 7, 1–5. [Google Scholar] [CrossRef]
- Bailey, R.W. The reaction of pentoses with anthrone. Biochem. J. 1958, 68, 669–672. [Google Scholar] [CrossRef] [PubMed]
- De Bruyn, J.W.; Van Keulen, H.A.; Ferguson, J.H.A. Rapid method for the simultaneous determination of glucose and fructose using anthrone reagent. J. Sci. Food Agric. 1968, 19, 597–601. [Google Scholar] [CrossRef]
- Desgranges, C.; Vergoignan, C.; Georges, M.; Durand, A. Biomass estimation in solid state fermentation I. Manual biochemical methods. Appl. Microbiol. Biotechnol. 1991, 35, 200–205. [Google Scholar] [CrossRef]
- Roche, N.; Venague, A.; Desgranges, C.; Durand, A. Use of chitin measurement to estimate fungal biomass in solid state fermentation. Biotechnol. Adv. 1993, 11, 677–683. [Google Scholar] [CrossRef]
- Brock, D.T. Studies on the Nutrition of Morchella esculenta Fries. Mycologia 1951, 43, 402–422. [Google Scholar] [CrossRef]
- Stott, K.; Mohammed, C. Specialty Mushroom Production Systems: Maitake and Morels. A report for the Rural Industries Research and Development Corporation; Rural Industries Research and Development Corporation: Barton, Australia, 2004. [Google Scholar]
- Zhang, G.P.; Zhang, F.; Ru, W.M.; Han, J.-R. Solid-state fermentation of cornmeal with the ascomycete Morchella esculenta for degrading starch and upgrading nutritional value. World J. Microbiol. Biotechnol. 2010, 26, 15. [Google Scholar] [CrossRef]
- Xing, Z.; Sun, F.; Liu, J. Studies on the submerged-cultured conditions of Morchella conica. Acta Edulis Fungi 2004, 11, 38–43. [Google Scholar]
- Buswell, A.J.; Chang, S. Biomass and extracellular hydrolytic enzyme production by six mushroom species grown on soybean waste. Biotechnol. Lett. 1994, 16, 1317–1322. [Google Scholar]
- Bensoussan, M.; Tisserand, E.; Kabbaji, W.; Roussos, S. Partial characterization of aroma produced by submerged culture of morel mushroom mycelium. Cryptog. Mycol. 1995, 16, 65–75. [Google Scholar]
- Meng, F.; Liu, X.; Jia, L.; Song, Z.; Deng, P.; Fan, K. Optimization for the production of exopolysaccharides from Morchella esculenta SO-02 in submerged culture and its antioxidant activities in vitro. Carbohydr. Polym. 2010, 79, 700–704. [Google Scholar] [CrossRef]
- Xu, H.; Sun, L.-P.; Shi, Y.-Z.; Wu, Y.-H.; Zhang, B.; Zhao, D.-Q. Optimization of cultivation conditions for extracellular polysaccharide and mycelium biomass by Morchella esculenta As51620. Biochem. Eng. J. 2008, 39, 66–73. [Google Scholar] [CrossRef]
- Sparringa, A.R.; Owens, D.J. Glucosamine content of tempe mould, Rhizopus oligosporus. Int. J. Food Microbiol. 1999, 47, 153–157. [Google Scholar] [CrossRef]
- Ruíz-Herrera, J.; Osorio, E. Isolation and chemical analysis of the cell wall of Morchella sp. Antonie van Leeuwenhoek 1974, 40, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Broecker, F.; Seeberger, P.H. Identification and Design of Synthetic B Cell Epitopes for Carbohydrate-Based Vaccines. Methods Enzymol. 2017, 597, 311–334. [Google Scholar] [PubMed]
- Tietel, Z.; Masaphy, S. True morels (Morchella)—Nutritional and phytochemical composition, health benefits and flavor: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1888–1901. [Google Scholar] [CrossRef] [PubMed]
- Zervakis, G.; Philippoussis, A.; Ioannidou, S.; Diamantopoulou, P. Mycelium growth kinetics and optimal temperature conditions for the cultivation of edible mushroom species on lignocellulosic substrates. Folia Microbiol. 2001, 46, 231. [Google Scholar] [CrossRef]
- Masaphy, S. Effect of CaCO3 on Morchella growth and sclerotia formation. Int. Soc. Mushroom Sci. 2004, 16. Available online: http://www.isms.biz/download/volume-16-part-1-article-14-effect-of-caco3-on-morchella-growth-and-sclerotia-formation/ (accessed on 2 June 2019).
- Reeslev, M.; Kjøller, A. Comparison of biomass dry weights and radial growth rates of fungal colonies on media solidified with different gelling compounds. Appl. Environ. Microbiol. 1995, 61, 4236–4239. [Google Scholar]
- Petre, V.; Petre, M.; Rusea, I.; Stǎnicǎ, F. Biotechnological recycling of fruit tree wastes by solid-state cultivation of mushrooms. In Mushroom Biotechnology, Developments and Applications; Petre, M., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 19–29. [Google Scholar]
- Cheung, P.C.K.; Lee, M.Y. Fractionation and Characterization of Mushroom Dietary Fiber (Nonstarch Polysaccharides) as Potential Nutraceuticals from Sclerotia of Pleurotus tuber-regium (Fries) Singer. J. Agric. Food Chem. 2000, 48, 3148–3151. [Google Scholar] [CrossRef]
- Alvarado-Castillo, G.; Mata, G.; Pérez-Vázquez, A.; Martínez-Carrera, D.; Tablada, M.E.N.; Gellardo-López, F.; Osorio-Acosta, F. Morchella sclerotia production through grain supplementation. Interciencia 2011, 36, 768–773. [Google Scholar]
- Papinutti, L.; Lechner, B. Influence of the carbon source on the growth and lignocellulolytic enzyme production by Morchella esculenta strains. J. Ind. Microbiol. Biotechnol. 2008, 35, 1715–1721. [Google Scholar] [CrossRef] [PubMed]
- Philippoussis, A.; Diamantopoulou, P. Exploitation of the biotechnological potential of agro-industrial by-products through mushroom cultivation. In Mushroom Biotechnology and Bioengineering; Petre, M., Berovic, M., Eds.; University of Pitesti: Bucharest, Romania, 2012; pp. 161–184. [Google Scholar]
- Lo, Y.C.; Lin, S.Y.; Ulziijargal, E.; Chen, S.Y.; Chien, R.C.; Tzou, Y.J.; Mau, J.L. Comparative study of contents of several bioactive components in fruiting bodies and mycelia of culinary-medicinal mushrooms. Int. J. Med. Mushrooms 2012, 14, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.R.; Barreto-Bergter, E.; Taborda, C.P. Glycoconjugates and polysaccharides of fungal cell wall and activation of immune system. Braz. J. Microbiol. 2008, 39, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Ochsenreither, K.; Glück, C.; Stressler, T.; Fischer, L.; Syldatk, C. Production Strategies and Applications of Microbial Single Cell Oils. Front. Microbiol. 2016, 7, 1539. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morels Group | Morchella Strain |
|---|---|
| Yellow morels | M. rotunda AMRL 14 |
| M. vulgaris AMRL 36 | |
| M. crassipes AMRL 52 | |
| Black morels | M. elata AMRL 63 |
| M. conica AMRL 74 | |
| M. angusticeps AMRL 82 |
| Morchella Strains | Medium | Time (days) | Biomass X (g/L) | Biomass Yield Yx/s (g/g) | Productivity PX (g/L/day) | Glucosamine (%, w/w) | TP (%, w/w) |
|---|---|---|---|---|---|---|---|
| AMRL 14 | PDB | 14 | 10.7 ± 0.67 | 0.42 | 0.76 | 3.0 ± 0.07 | 9.6 ± 0.60 |
| GPYB | 14 | 11.1 ± 0.80 | 0.41 | 0.79 | 3.7 ± 0.08 | 10.6 ± 0.03 | |
| AMRL 36 | PDB | 7 | 9.8 ± 0.45 | 0.41 | 1.40 | 2.5 ± 0.07 | 9.7 ± 0.02 |
| GPYB | 7 | 10.2 ± 0.07 | 0.52 | 1.46 | 2.3 ± 0.08 | 10.8 ± 0.31 | |
| AMRL 52 | PDB | 7 | 9.3 ± 0.40 | 0.43 | 1.33 | 2.5 ± 0.09 | 10.1 ± 0.05 |
| GPYB | 14 | 10.9 ± 0.35 | 0.43 | 0.78 | 3.1 ± 0.06 | 11.6 ± 0.30 | |
| AMRL 63 | PDB | 7 | 10.6 ± 0.45 | 0.43 | 1.51 | 2.3 ± 0.06 | 11.8 ± 0.12 |
| GPYB | 14 | 14.2 ± 0.10 | 0.47 | 1.01 | 3.0 ± 0.04 | 12.2 ± 0.06 | |
| AMRL 74 | PDB | 7 | 11.8 ± 0.75 | 0.47 | 1.69 | 2.4 ± 0.04 | 10.3 ± 0.14 |
| GPYB | 14 | 9.4 ± 0.97 | 0.32 | 0.67 | 3.1 ± 0.08 | 10.5 ± 0.10 | |
| AMRL 82 | PDB | 14 | 10.9 ± 0.55 | 0.42 | 0.78 | 3.5 ± 0.07 | 10.4 ± 0.30 |
| GPYB | 14 | 12.4 ± 0.05 | 0.56 | 0.89 | 2.8 ± 0.05 | 10.9 ± 0.23 |
| Morchella Strains | Medium | Biomass (y)/Glucosamine (x) | Glucosamine (y)/Biomass (x) | R2 |
|---|---|---|---|---|
| AMRL 14 | PDB | y = 0.0335x − 0.0256 | y = 29.499x + 0.8482 | 0.99 |
| GPYB | y = 0.0277x − 0.0046 | y = 34.845x + 0.4457 | 0.97 | |
| AMRL 36 | PDB | y = 0.0376x − 0.0064 | y = 26.476x + 0.1888 | 0.99 |
| GPYB | y = 0.0336x − 0.0055 | y = 29.308x + 0.2457 | 0.98 | |
| AMRL 52 | PDB | y = 0.0369x − 0.0026 | y = 26.151x + 0.2788 | 0.97 |
| GPYB | y = 0.0301x − 0.0002 | y = 32.581x + 0.1026 | 0.98 | |
| AMRL 63 | PDB | y = 0.0378x + 0.0020 | y = 25.753x + 0.0513 | 0.97 |
| GPYB | y = 0.0328x − 0.0010 | y = 30.057x + 0.0825 | 0.99 | |
| AMRL 74 | PDB | y = 0.0385x + 0.0045 | y = 25.627x - 0.0563 | 0.99 |
| GPYB | y = 0.0306x − 0.0124 | y = 32.223x + 0.4669 | 0.99 | |
| AMRL 82 | PDB | y = 0.027x − 0.0073 | y = 36.685x + 0.3512 | 0.99 |
| GPYB | y = 0.0355x − 0.0219 | y = 27.988x + 0.6530 | 0.99 |
| Medium | Time (days) | Strains/Sclerotia Number and Maturation | |||||||
|---|---|---|---|---|---|---|---|---|---|
| AMRL 36 | AMRL 52 | AMRL 63 | AMRL 74 | ||||||
| I 1 | M 2 | I | M | I | M | I | M | ||
| PDA | 8 | + | − | + | − | + | − | + | − |
| 13 | + | − | + | − | ++ | * | + | − | |
| 21 | + | * | + | * | ++ | * | + | * | |
| PDA–PET | 12 | − | * | − | * | + | * | + | − |
| 22 | + | * | + | ** | ++ | ** | + | * | |
| 34 | ++ | * | ++ | ** | ++ | *** | ++ | * | |
| Medium | Time (days) | Strains/Sclerotia Number and Maturation | |||
|---|---|---|---|---|---|
| AMRL 52 | AMRL 63 | ||||
| I 1 | M 2 | I | M | ||
| WG–PET | 20 | + | − | + | − |
| 30 | ++ | − | + | * | |
| PP–PET | 20 | − | − | − | − |
| 30 | − | − | − | − | |
| WG:PP–PET | 20 | + | * | ++ | * |
| 30 | + | * | ++ | ** | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papadaki, A.; Diamantopoulou, P.; Papanikolaou, S.; Philippoussis, A. Evaluation of Biomass and Chitin Production of Morchella Mushrooms Grown on Starch-Based Substrates. Foods 2019, 8, 239. https://doi.org/10.3390/foods8070239
Papadaki A, Diamantopoulou P, Papanikolaou S, Philippoussis A. Evaluation of Biomass and Chitin Production of Morchella Mushrooms Grown on Starch-Based Substrates. Foods. 2019; 8(7):239. https://doi.org/10.3390/foods8070239
Chicago/Turabian StylePapadaki, Aikaterini, Panagiota Diamantopoulou, Seraphim Papanikolaou, and Antonios Philippoussis. 2019. "Evaluation of Biomass and Chitin Production of Morchella Mushrooms Grown on Starch-Based Substrates" Foods 8, no. 7: 239. https://doi.org/10.3390/foods8070239
APA StylePapadaki, A., Diamantopoulou, P., Papanikolaou, S., & Philippoussis, A. (2019). Evaluation of Biomass and Chitin Production of Morchella Mushrooms Grown on Starch-Based Substrates. Foods, 8(7), 239. https://doi.org/10.3390/foods8070239