Physicochemical Characterization and Biological Activities of Black and White Garlic: In Vivo and In Vitro Assays †

 , ,
, ,  , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Samples
2.2. Measurement of Soluble Solid Content, pH, aw, and Browning Intensity
2.3. Total Polyphenol Content and Antioxidant Capacity
2.4. In Vivo Assays
2.4.1. D. melanogaster Strains
2.4.2. Toxicity and Antitoxicity Assays
2.4.3. Genotoxicity and Antigenotoxicity Assays
2.4.4. Lifespan Assays
2.5. In Vitro Assays
2.5.1. HL-60 Cell Line Culture Conditions
2.5.2. Cytotoxicity Assay
2.5.3. Determination of DNA Fragmentation
2.6. Statistical Analysis
3. Results and Discussion
3.1. Soluble Solids Content, pH, Water Activity, and Browning Intensity
3.2. Total Polyphenol Content and Antioxidant Capacity
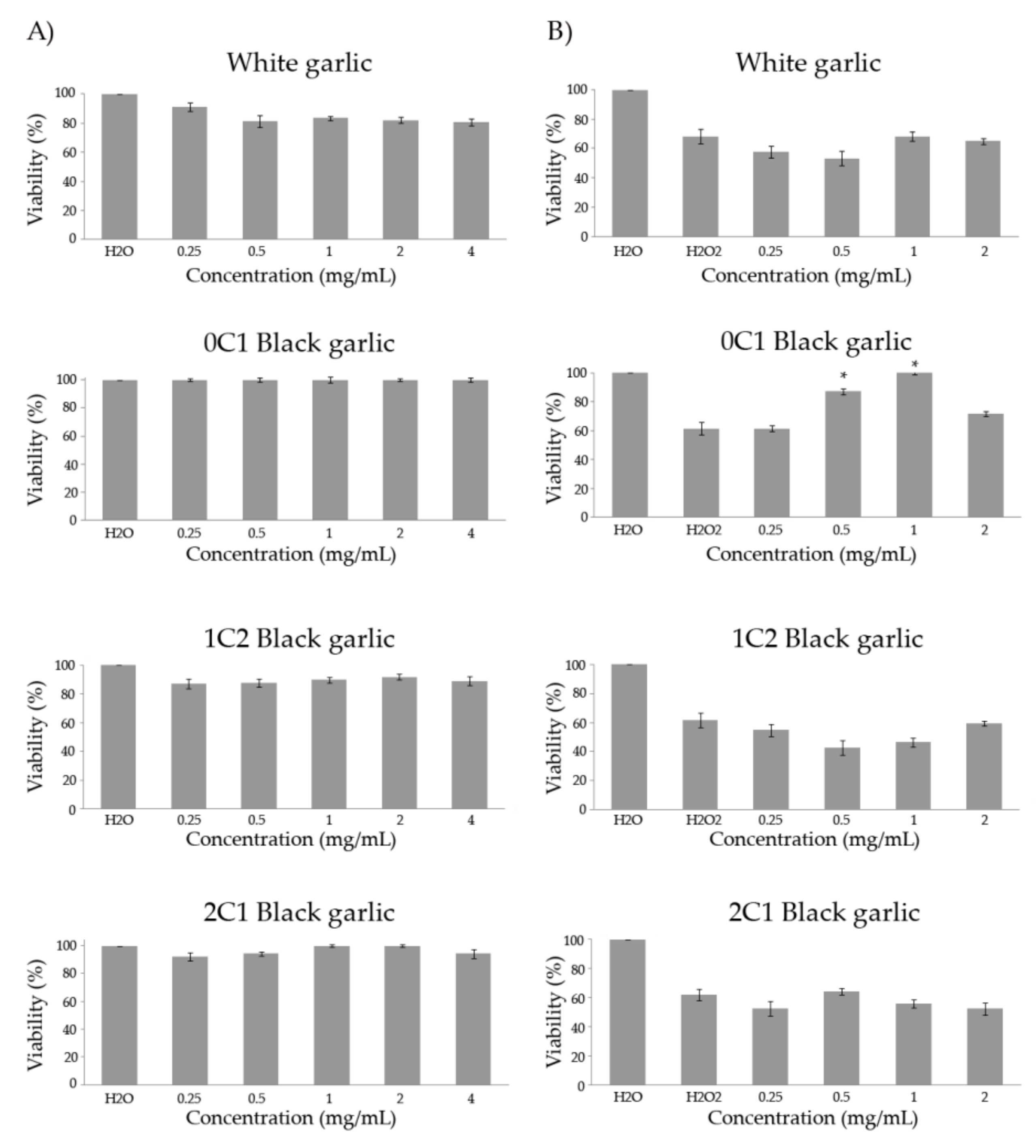
3.3. Toxicity/Antitoxicity
3.4. Genotoxicity/Antigenotoxicity
3.5. Longevity Assays
3.6. Healthspan Assays
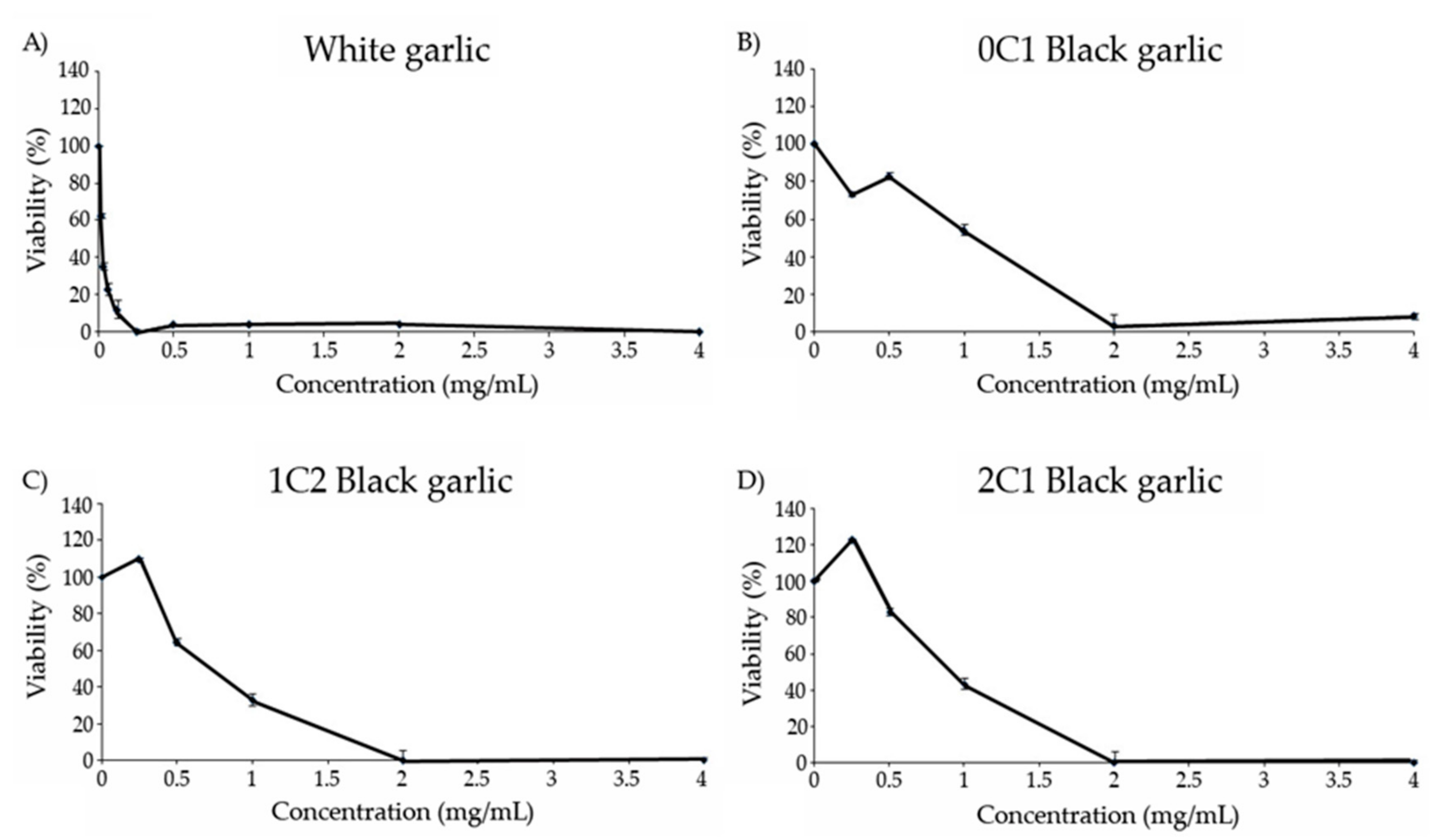
3.7. Cytotoxicity
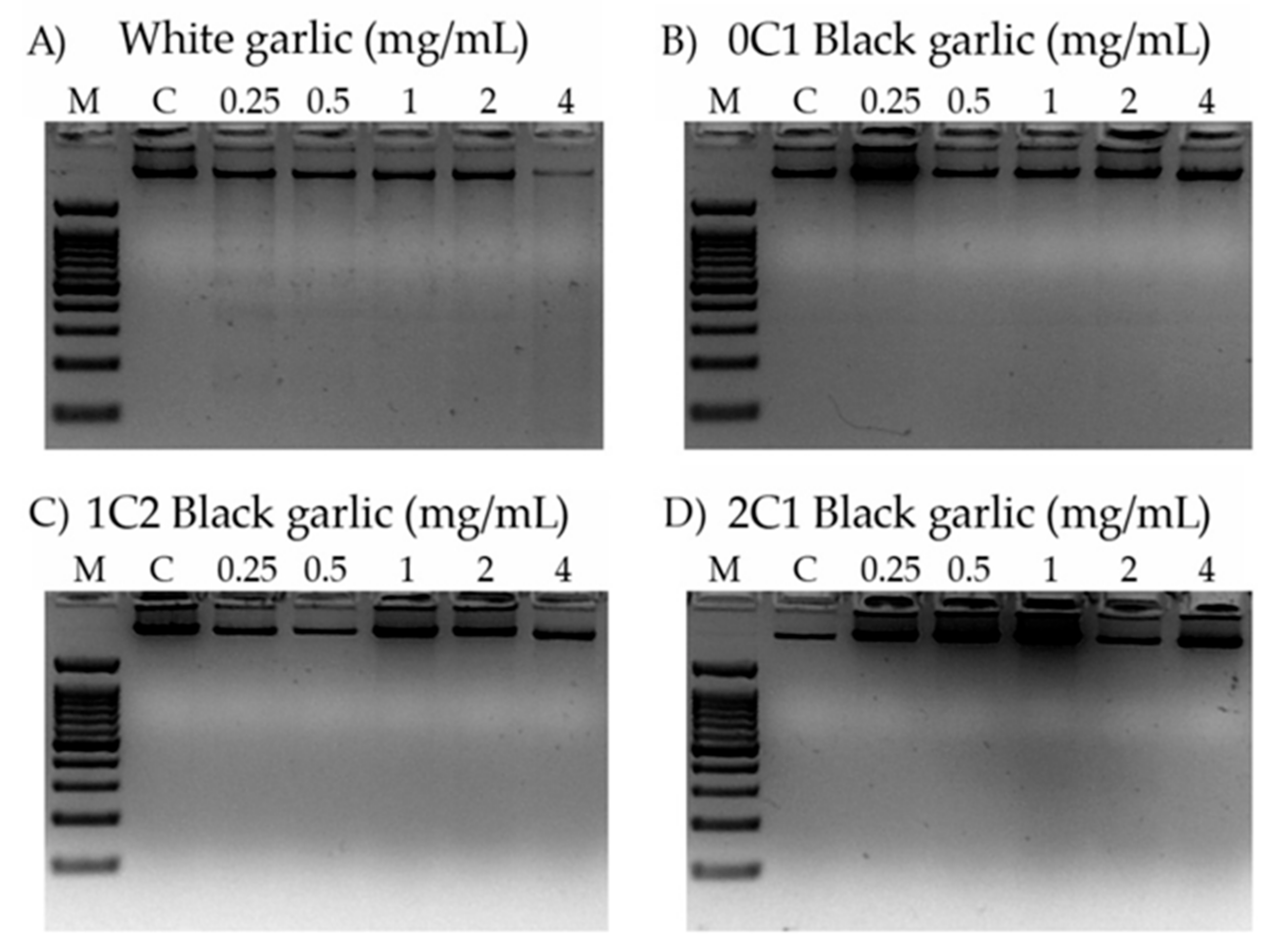
3.8. DNA Internucleosomal Fragmentation
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Block, E. The chemistry of garlic and onions. Sci. Am. 1985, 252, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Rivlin, R.S. Is garlic alternative medicine? J. Nutr. 2006, 136, 713S–715S. [Google Scholar] [CrossRef] [PubMed]
- Tattelman, E. Health effects of garlic. Am. Fam. Physician 2005, 72, 103–106. [Google Scholar] [PubMed]
- Amagase, H. Clarifying the real bioactive constituents of garlic. J. Nutr. 2006, 136, 716S–725S. [Google Scholar] [CrossRef] [PubMed]
- Cerella, C.; Dicato, M.; Jacob, C.; Diederich, M. Chemical properties and mechanisms determining the anti-cancer action of garlic-derived organic sulfur compounds. Anti-Cancer Agents Med. Chem. 2011, 11, 267–271. [Google Scholar] [CrossRef]
- Corzo-Martínez, M.; Corzo, N.; Villamiel, M. Biological properties of onions and garlic. Trends Food Sci. Technol. 2007, 18, 609–625. [Google Scholar] [CrossRef]
- Chowdhury, R.; Dutta, A.; Chaudhuri, S.R.; Sharma, N.; Giri, A.K.; Chaudhuri, K. In vitro and in vivo reduction of sodium arsenite induced toxicity by aqueous garlic extract. Food Chem. Toxicol. 2008, 46, 740–751. [Google Scholar] [CrossRef] [PubMed]
- Brodnitz, M.H.; Pascale, J.V. Thiopropanal s-oxide: A lachrymatory factor in onions. J. Agric. Food Chem. 1971, 19, 269–272. [Google Scholar] [CrossRef]
- Rana, S.; Pal, R.; Vaiphei, K.; Singh, K. Garlic hepatotoxicity: Safe dose of garlic. Trop. Gastroenterol. 2006, 27, 26–30. [Google Scholar]
- Ryu, J.H.; Kang, D. Physicochemical properties, biological activity, health benefits, and general limitations of aged black garlic: A review. Molecules 2017, 22, 919. [Google Scholar] [CrossRef] [PubMed]
- Toledano-Medina, M.A.; Pérez-Aparicio, J.; Moreno-Rojas, R.; Merinas-Amo, T. Evolution of some physicochemical and antioxidant properties of black garlic whole bulbs and peeled cloves. Food Chem. 2016, 199, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Capuano, E.; Fogliano, V. Acrylamide and 5-hydroxymethylfurfural (HMF): A review on metabolism, toxicity, occurrence in food and mitigation strategies. LWT—Food Sci. Technol. 2011, 44, 793–810. [Google Scholar] [CrossRef]
- Shinkawa, H.; Takemura, S.; Minamiyama, Y.; Kodai, S.; Tsukioka, T.; Osada-Oka, M.; Kubo, S.; Okada, S.; Suehiro, S. S-allylcysteine is effective as a chemopreventive agent against porcine serum-induced hepatic fibrosis in rats. Osaka City Med. J. 2009, 55, 61–69. [Google Scholar] [PubMed]
- Imai, J.; Ide, N.; Nagae, S.; Moriguchi, T.; Matsuura, H.; Itakura, Y. Antioxidant and radical scavenging effects of aged garlic extract and its constituents. Planta Med. 1994, 60, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-M.; Gweon, O.-C.; Seo, Y.-J.; Im, J.; Kang, M.-J.; Kim, M.-J.; Kim, J.-I. Antioxidant effect of garlic and aged black garlic in animal model of type 2 diabetes mellitus. Nutr. Res. Pract. 2009, 3, 156–161. [Google Scholar] [CrossRef]
- Sasaki, J.-I.; Lu, C.; Machiya, E.; Tanahashi, M.; Hamada, K. Processed black garlic (allium sativum) extracts enhance anti-tumor potency against mouse tumors. Med. Aromat. Plant Sci. Biotechnol. 2007, 227, 138. [Google Scholar]
- Nai-feng, Y.G.-m.X.; Bao-cui, Q.D.-w.Z.; Ya-yu, Z. Study on the technology of black garlic drink fermented by immobilized lactobacillus. Shandong Food Ferment. 2012, 4, 005. [Google Scholar]
- William, W.J.; Carvalho, G.B.; Mak, E.M.; Noelle, N.; Fang, A.Y.; Liong, J.C.; Brummel, T.; Benzer, S. Prandiology of drosophila and the cafe assay. Proc. Natl. Acad. Sci. USA 2007, 104, 8253–8256. [Google Scholar]
- Klein, S.; Rister, R.; Riggins, C. The Complete German Commission E Monographs: Therapeutic Guide to Herbal Medicines; American Botanical Council: Austin, TX, USA, 1998; p. 356. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Elsevier Ltd.: Amsterdam, The Netherlands, 1999; pp. 152–178. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved abts radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Yan, J.; Huen, D.; Morely, T.; Johnson, G.; Gubb, D.; Roote, J.; Adler, P.N. The multiple-wing-hairs gene encodes a novel GBD-FH3 domain-containing protein that functions both prior to and after wing hair initiation. Genetics 2008, 180, 219–228. [Google Scholar] [CrossRef]
- Ren, N.; Charlton, J.; Adler, P.N. The flare gene, which encodes the aip1 protein of drosophila, functions to regulate f-actin disassembly in pupal epidermal cells. Genetics 2007, 176, 2223–2234. [Google Scholar] [CrossRef] [PubMed]
- Lindsley, D.L.; Zimm, G.G. The Genome of Drosophila Melanogaster; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Fernández-Bedmar, Z.; Anter, J.; Alonso Moraga, Á. Anti/genotoxic, longevity inductive, cytotoxic, and clastogenic-related bioactivities of tomato andlycopene. Environ. Mol. Mutagen. 2018, 59, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Graf, U.; Würgler, F.; Katz, A.; Frei, H.; Juon, H.; Hall, C.; Kale, P. Somatic mutation and recombination test in drosophila melanogaster. Environ. Mutagen. 1984, 6, 153–188. [Google Scholar] [CrossRef] [PubMed]
- Graf, U.; Abraham, S.K.; Guzmán-Rincón, J.; Würgler, F.E. Antigenotoxicity studies in drosophila melanogaster. Mutat. Res. 1998, 402, 203–209. [Google Scholar] [CrossRef]
- Moraga, A.A.; Graf, U. Genotoxicity testing of antiparasitic nitrofurans in the drosophila wing somatic mutation and recombination test. Mutagenesis 1989, 4, 105–110. [Google Scholar] [CrossRef]
- Frei, H.; Würgler, F. Statistical methods to decide whether mutagenicity test data from drosophila assays indicate a positive, negative, or inconclusive result. Mutat. Res. 1988, 203, 297–308. [Google Scholar] [CrossRef]
- Frei, H.; Würgler, F.E. Optimal experimental design and sample size for the statistical evaluation of data from somatic mutation and recombination tests (smart) in drosophila. Mutat. Res. 1995, 334, 247–258. [Google Scholar] [CrossRef]
- Abraham, S.K. Antigenotoxicity of coffee in the drosophila assay for somatic mutation and recombination. Mutagenesis 1994, 9, 383–386. [Google Scholar] [CrossRef]
- Gallagher, R.; Collins, S.; Trujillo, J.; McCredie, K.; Ahearn, M.; Tsai, S.; Metzgar, R.; Aulakh, G.; Ting, R.; Ruscetti, F. Characterization of the continuous, differentiating myeloid cell line (hl-60) from a patient with acute promyelocytic leukemia. Blood 1979, 54, 713–733. [Google Scholar]
- Merinas-Amo, T.; Tasset-Cuevas, I.; Díaz-Carretero, A.M.; Alonso-Moraga, Á.; Calahorro, F. In vivo and in vitro studies of the role of lyophilised blond lager beer and some bioactive components in the modulation of degenerative processes. J. Funct. Foods 2016, 27, 274–294. [Google Scholar] [CrossRef]
- Jones, M.; Collin, H.; Tregova, A.; Trueman, L.; Brown, L.; Cosstick, R.; Hughes, J.; Milne, J.; Wilkinson, M.; Tomsett, A. The biochemical and physiological genesis of alliin in garlic. Med. Aromat. Plant Sci. Biotechnol. 2007, 1, 21–24. [Google Scholar]
- Choi, D.-J.; Lee, S.-J.; Kang, M.-J.; Cho, H.-S.; Sung, N.-J.; Shin, J.-H. Physicochemical characteristics of black garlic (Allium sativum L.). J. Korean Soc. Food Sci. Nutr. 2008, 37, 465–471. [Google Scholar] [CrossRef]
- Shin, J.-H.; Choi, D.-J.; Lee, S.-J.; Cha, J.-Y.; Kim, J.-G.; Sung, N.-J. Changes of physicochemical components and antioxidant activity of garlic during its processing. J. Life Sci. 2008, 18, 1123–1131. [Google Scholar] [CrossRef]
- Bae, S.E.; Cho, S.Y.; Won, Y.D.; Lee, S.H.; Park, H.J. Changes in s-allyl cysteine contents and physicochemical properties of black garlic during heat treatment. LWT—Food Sci. Technol. 2014, 55, 397–402. [Google Scholar] [CrossRef]
- Kaanane, A.; Labuza, T. The Maillard reaction in foods. In The Maillard Reaction in Aging, Diabetes and Nutrition; AR Liss Press, Inc.: New York, NY, USA, 1989. [Google Scholar]
- Labuza, T.P.; Saltmarch, M. The nonenzymatic browning reaction as affected by water in foods. In Water Activity: Influences on Food Quality; Elsevier: Amsterdam, The Netherlands, 1981; pp. 605–650. [Google Scholar]
- Choi, J.-H.; Kim, W.-J.; Yang, J.-W.; Sung, H.-S.; Hong, S.-K. Quality changes in red ginseng extract during high temperature storage. Appl. Biol. Chem. 1981, 24, 50–58. [Google Scholar]
- Kim, M.-H.; Kim, B.-Y. Development of optimum processing conditions in air dried garlics using response surface methodology. J. Korean Soc. Food Nutr. 1990, 19, 234–238. [Google Scholar]
- Kang, Y.-H.; Park, Y.-K.; Lee, G.-D. The nitrite scavenging and electron donating ability of phenolic compounds. Korean J. Food Sci. Technol. 1996, 28, 232–239. [Google Scholar]
- Jang, E.-K.; Seo, J.-H.; Lee, S.-P. Physiological activity and antioxidative effects of aged black garlic (Allium sativum L.) extract. Korean J. Food Sci. Technol. 2008, 40, 443–448. [Google Scholar]
- Kakimoto, M.; Suzuki, A.; Nishimoto, I. Iadditives, supplements, antioxidants and phyto-chemicals. Trends Food Sci. Technol. 2000, 11, 266–267. [Google Scholar]
- Xu, B.; Chang, S.K. Total phenolics, phenolic acids, isoflavones, and anthocyanins and antioxidant properties of yellow and black soybeans as affected by thermal processing. J. Agric. Food Chem. 2008, 56, 7165–7175. [Google Scholar] [CrossRef]
- Gorinstein, S.; Leontowicz, M.; Leontowicz, H.; Najman, K.; Namiesnik, J.; Park, Y.-S.; Jung, S.-T.; Kang, S.-G.; Trakhtenberg, S. Supplementation of garlic lowers lipids and increases antioxidant capacity in plasma of rats. Nutr. Res. 2006, 26, 362–368. [Google Scholar] [CrossRef]
- Nakagawa, S.; Masamoto, K.; Sumiyoshi, H.; Harada, H. Acute toxicity test of garlic extract. J. Toxicol. Sci. 1984, 9, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Sumiyoshi, H.; Kanezawa, A.; Masamoto, K.; Harada, H.; Nakagami, S.; Yokota, A.; Nishikawa, M.; Nakagawa, S. Chronic toxicity test of garlic extract in rats. J. Toxicol. Sci. 1984, 9, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Kanezawa, A.; Nakagawa, S.; Sumiyoshi, H.; Masamoto, K.; Harada, H.; Nakagami, S.; Date, S.; Yokota, A.; Nishikawa, M.; Fuwa, T. General toxicity tests of garlic extract preparation (kyoleopin) containing vitamins. Oyo Yakuri 1984, 27, 909–929. [Google Scholar]
- Nakagawa, S.; Masamoto, K.; Sumiyoshi, H.; Kunihiro, K.; FUWA, T. Effect of raw and extracted-aged garlic juice on growth of young rats and their organs after peroral administration. J. Toxicol. Sci. 1980, 5, 91–112. [Google Scholar] [CrossRef]
- Assayed, M.; Khalaf, A.; Salem, H. Protective effects of garlic extract and vitamin C against in vivo cypermethrin-induced teratogenic effects in rat offspring. Food Chem. Toxicol. 2010, 48, 3153–3158. [Google Scholar] [CrossRef] [PubMed]
- Steiner, M.; Khan, A.H.; Holbert, D.; Lin, R. A double-blind crossover study in moderately hypercholesterolemic men that compared the effect of aged garlic extract and placebo administration on blood lipids. Am. J. Clinic. Nutr. 1996, 64, 866–870. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.-M.; Xu, M.-Y.; Zhang, Z.-S.; Zhang, M.; Gao, Y.-F. The analysis of saccharide in black garlic and its antioxidant activity. Adv. J. Food Sci. Technol. 2014, 6, 755–760. [Google Scholar] [CrossRef]
- Fernández-Bedmar, Z.; Alonso-Moraga, A. In vivo and in vitro evaluation for nutraceutical purposes of capsaicin, capsanthin, lutein and four pepper varieties. Food Chem. Toxicol. 2016, 98, 89–99. [Google Scholar] [CrossRef]
- Rojas-Molina, M.; Campos-Sánchez, J.; Analla, M.; Muñoz-Serrano, A.; Alonso-Moraga, Á. Genotoxicity of vegetable cooking oils in the drosophila wing spot test. Environ. Mol. Mutagen. 2005, 45, 90–95. [Google Scholar] [CrossRef]
- Romero-Jimenez, M.; Campos-Sanchez, J.; Analla, M.; Muñoz-Serrano, A.; Alonso-Moraga, Á. Genotoxicity and anti-genotoxicity of some traditional medicinal herbs. Mutat. Res. 2005, 585, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.K.; Kesavan, P. Genotoxicity of garlic, turmeric and asafoetida in mice. Mutat. Res. 1984, 136, 85–88. [Google Scholar] [CrossRef]
- Shukla, Y.; Taneja, P. Antimutagenic effects of garlic extract on chromosomal aberrations. Cancer Let. 2002, 176, 31–36. [Google Scholar] [CrossRef]
- Sowjanya, B.L.; Devi, K.R.; Madhavi, D. Modulatory effects of garlic extract against the cyclophosphamide induced genotoxicity in human lymphocytes in vitro. J. Environ. Boil. 2009, 30, 663. [Google Scholar]
- Chughtai, S.R.; Dhmad, M.; Khalid, N.; Mohamed, A. Genotoxicity testing of some spices in diploid yeast. Pak. J. Bot. 1998, 30, 33–38. [Google Scholar]
- Fernández-Bedmar, Z.; Anter, J.; de La Cruz-Ares, S.; Muñoz-Serrano, A.; Alonso-Moraga, Á.; Pérez-Guisado, J. Role of citrus juices and distinctive components in the modulation of degenerative processes: Genotoxicity, antigenotoxicity, cytotoxicity, and longevity in drosophila. J. Toxicol. Environ. Health 2011, 74, 1052–1066. [Google Scholar] [CrossRef]
- Park, J.-H.; Park, Y.K.; Park, E. Antioxidative and antigenotoxic effects of garlic (Allium sativum L.) prepared by different processing methods. Plant Foods Human Nutr. 2009, 64, 244. [Google Scholar] [CrossRef]
- Benkeblia, N. Free-radical scavenging capacity and antioxidant properties of some selected onions (allium cepa l.) and garlic (Allium sativum L.) extracts. Braz. Arch. Boil. Technol. 2005, 48, 753–759. [Google Scholar] [CrossRef]
- Prakash, D.; Singh, B.N.; Upadhyay, G. Antioxidant and free radical scavenging activities of phenols from onion (allium cepa). Food Chem. 2007, 102, 1389–1393. [Google Scholar] [CrossRef]
- Kim, S.H.; Jung, E.Y.; Kang, D.H.; Chang, U.J.; Hong, Y.-H.; Suh, H.J. Physical stability, antioxidative properties, and photoprotective effects of a functionalized formulation containing black garlic extract. J. Photochem. Photobiol. B 2012, 117, 104–110. [Google Scholar] [CrossRef]
- Bingqiao, Z.; Haige, W.; Yuanyuan, L. The antioxidation of black garlic. Food Res. Dev. 2008, 29, 56–61. [Google Scholar]
- Huang, C.-H.; Hsu, F.-Y.; Wu, Y.-H.; Zhong, L.; Tseng, M.-Y.; Kuo, C.-J.; Hsu, A.-L.; Liang, S.-S.; Chiou, S.-H. Analysis of lifespan-promoting effect of garlic extract by an integrated metabolo-proteomics approach. J. Nutr. Biochem. 2015, 26, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Momoucm, T.; Takashina, P. Prolongation of life span and improved learning in the senescence accelerated mouse produced by aged garlic extract. Biol. Pharm. Bull. 1994, 7, l1589–l1594. [Google Scholar]
- Prowse, G.M.; Galloway, T.S.; Foggo, A. Insecticidal activity of garlic juice in two dipteran pests. Agric. For. Entomol. 2006, 8, 1–6. [Google Scholar] [CrossRef]
- Soh, J.W.; Hotic, S.; Arking, R. Dietary restriction in drosophila is dependent on mitochondrial efficiency and constrained by pre-existing extended longevity. Mech. Ageing Dev. 2007, 128, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.; Kusumoto, Y.; Al-Mamun, M.A. Cytotoxicity and cancer (HeLa) cell killing efficacy of aqueous garlic (Allium sativum) extract. J. Sci. Res. 2011, 3, 375–382. [Google Scholar] [CrossRef]
- Lau, B.H.; Tadi, P.P.; Tosk, J.M. Allium sativum (garlic) and cancer prevention. Nutr. Res. 1990, 10, 937–948. [Google Scholar] [CrossRef]
- Seki, T.; Tsuji, K.; Hayato, Y.; Moritomo, T.; Ariga, T. Garlic and onion oils inhibit proliferation and induce differentiation of hl-60 cells. Cancer Let. 2000, 160, 29–35. [Google Scholar] [CrossRef]
- Wang, X.; Jiao, F.; Wang, Q.-W.; Wang, J.; Yang, K.; Hu, R.-R.; Liu, H.-C.; Wang, H.-Y.; Wang, Y.-S. Aged black garlic extract induces inhibition of gastric cancer cell growth in vitro and in vivo. Mol. Med. Rep. 2012, 5, 66–72. [Google Scholar] [CrossRef]
- Katsuki, T.; Hirata, K.; Ishikawa, H.; Matsuura, N.; Sumi, S.-I.; Itoh, H. Aged garlic extract has chemopreventative effects on 1,2-dimethylhydrazine-induced colon tumors in rats. J. Nutr. 2006, 136, 847S–851S. [Google Scholar] [CrossRef]
- Wyllie, A.; Kerr, J.; Currie, A. Apoptosis: The significance of cell death. Int. Rev. Cytol. 1980, 68, 251–306. [Google Scholar] [PubMed]
- Hong, Y.-S.; Ham, Y.-A.; Choi, J.-H.; Kim, J. Effects of allyl sulfur compounds and garlic extract on the expression of Bcl-2, Bax, and p53 in non small cell lung cancer cell lines. Exp. Mol. Med. 2000, 32, 127. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Garlic | White | 0C1 Black | 1C2 Black | 2C1 Black |
|---|---|---|---|---|
| Aging process (days) | 0 | 13 | 32 | 45 |
| Weight of 10 garlic cloves (g) | 49.69 ± 0.25 a,1 | 45.83 ± 0.32 b,c | 37.57 ± 0.38 b | 19.67 ± 0.19 d |
| Soluble solid content (ºBrix) | 40.47 ± 0.29 c | 40.47 ± 0.34 c | 43.17 ± 0.48 b | 45.67 ± 0.42 a |
| pH | 5.94 ± 0.01 a | 3.69 ± 0.03 b | 3.60 ± 0.04 c | 3.49 ± 0.06 d |
| Water activity (aw) | 0.97 ± 0 a | 0.93 ± 0 c | 0.93 ± 0 c | 0.93 ± 0 c |
| Browning intensity (L) | 47.16 ± 0.15 a | 18.73 ± 0.21 c | 17.85 ± 0.24 b | 17.58 ± 0.25 b |
| Polyphenol content (g GAE/kg) | 4.30 ± 0.04 d | 10.94 ± 0.28 c | 14.67 ± 0.19 b | 16.17 ± 0.29 a |
| Antioxidant activity (TROLOX equivalents/kg) | 10.20 ± 0.27 d | 67.65 ± 1.26 b | 57.35 ± 1.74 c | 78.61 ± 2.41 a |
| Compound | Clones per Wing (n° spots) 1 | Inhibition Percentage (%) 2 | ||||
|---|---|---|---|---|---|---|
| Number of Wings | Small Single Clones (1–2 Cells) m = 2 | Large Simple Clones (More Than 2 Cells) m = 5 | Twin Clones m = 5 | Total Clones m = 2 | ||
| H2O | 41 | 0.146 (6) | 0.049 (2) | 0 | 0.195 (8) | |
| H2O2 | 40 | 0.350 (14) | 0.075 (3) | 0 | 0.425 (17) + | |
| Simple Treatment | ||||||
| White garlic (mg/mL) | ||||||
| 0.25 | 40 | 0.225 (9) | 0.025 (1) | 0.025 (1) | 0.275 (11) − | |
| 2 | 40 | 0.375 (15) | 0.050 (2) | 0.000 | 0.425 (17) + | |
| 0C1 Black garlic (mg/mL) | ||||||
| 0.25 | 40 | 0.175 (7) | 0.025 (1) | 0.000 | 0.200 (8) − | |
| 2 | 41 | 0.122 (5) | 0.000 | 0.000 | 0.122 (5) − | |
| 1C2 Black garlic (mg/mL) | ||||||
| 0.25 | 40 | 0.200 (8) | 0.025 (1) | 0.025 (1) | 0.250 (10) - | |
| 2 | 40 | 0.175 (7) | 0.025 (1) | 0.000 | 0.200 (8) - | |
| 2C1 Black garlic (mg/mL) | ||||||
| 0.25 | 40 | 0.175 (7) | 0.05 (2) | 0.000 | 0.225 (9) − | |
| 2 | 40 | 0.250 (10) | 0.000 | 0.000 | 0.250 (10) − | |
| Combined Treatment With H2O2 (0.12 M) | ||||||
| White garlic (mg/mL) | ||||||
| 0.25 | 34 | 0.235 (8) | 0.088 (3) | 0.000 | 0.323 (11) − | 24 |
| 2 | 34 | 0.265 (10) | 0.206 (7) | 0.000 | 0.500 (17) + | −17 |
| 0C1 Black garlic (mg/mL) | ||||||
| 0.25 | 30 | 0.5 (15) | 0.033 (1) | 0.000 | 0.533 (16) + | −25.4 |
| 2 | 30 | 0.233 (7) | 0.033 (1) | 0.000 | 0.266 (8) − | 37.4 |
| 1C2 Black garlic (mg/mL) | ||||||
| 0.25 | 26 | 0.307 (8) | 0.038 (1) | 0.000 | 0.346 (9) − | 18.6 |
| 2 | 38 | 0.368 (14) | 0.053 (2) | 0.000 | 0.421 (16) − | 0.17 |
| 2C1 Black garlic (mg/mL) | ||||||
| 0.25 | 28 | 0.357 (10) | 0.036 (1) | 0.000 | 0.393 (11) − | 7.5 |
| 2 | 28 | 0.357 (10) | 0.250 (7) | 0.000 | 0.607 (17) + | −42.8 |
| Compound Title | Treatment (mg/mL) | Mean Lifespan (Days) | Mean Healthspan (Days) | ||
|---|---|---|---|---|---|
| Negative Control | 0 | 60.31 | 32.46 | ||
| WG | 0.25 | 70.47 | * | 38.40 | ns |
| 0.5 | 68.72 | ns | 29.40 | ns | |
| 1 | 71.43 | ** | 40.39 | * | |
| 2 | 78.89 | *** | 40.44 | * | |
| 4 | 58.21 | ns | 31.91 | ns | |
| 0C1 | 0.25 | 70.15 | * | 37.67 | ns |
| 0.5 | 71.72 | ** | 38.18 | ns | |
| 1 | 70.73 | * | 30.85 | ns | |
| 2 | 71.80 | * | 43.90 | * | |
| 4 | 62.60 | ns | 33.10 | ns | |
| 1C2 | 0.25 | 69.73 | * | 29.36 | ns |
| 0.5 | 65.19 | ns | 25.15 | * | |
| 1 | 70.49 | ** | 31.10 | ns | |
| 2 | 70.18 | * | 28.57 | ns | |
| 4 | 60.26 | ns | 26.20 | ns | |
| 2C1 | 0.25 | 59.06 | ns | 24.07 | ns |
| 0.5 | 64.75 | ns | 41.29 | ns | |
| 1 | 59.82 | ns | 28.50 | ns | |
| 2 | 57.30 | ns | 23.46 | ** | |
| 4 | 66.38 | ns | 40.50 | ns | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toledano Medina, M.Á.; Merinas-Amo, T.; Fernández-Bedmar, Z.; Font, R.; del Río-Celestino, M.; Pérez-Aparicio, J.; Moreno-Ortega, A.; Alonso-Moraga, Á.; Moreno-Rojas, R. Physicochemical Characterization and Biological Activities of Black and White Garlic: In Vivo and In Vitro Assays. Foods 2019, 8, 220. https://doi.org/10.3390/foods8060220
Toledano Medina MÁ, Merinas-Amo T, Fernández-Bedmar Z, Font R, del Río-Celestino M, Pérez-Aparicio J, Moreno-Ortega A, Alonso-Moraga Á, Moreno-Rojas R. Physicochemical Characterization and Biological Activities of Black and White Garlic: In Vivo and In Vitro Assays. Foods. 2019; 8(6):220. https://doi.org/10.3390/foods8060220
Chicago/Turabian StyleToledano Medina, María Ángeles, Tania Merinas-Amo, Zahira Fernández-Bedmar, Rafael Font, Mercedes del Río-Celestino, Jesús Pérez-Aparicio, Alicia Moreno-Ortega, Ángeles Alonso-Moraga, and Rafael Moreno-Rojas. 2019. "Physicochemical Characterization and Biological Activities of Black and White Garlic: In Vivo and In Vitro Assays" Foods 8, no. 6: 220. https://doi.org/10.3390/foods8060220
APA StyleToledano Medina, M. Á., Merinas-Amo, T., Fernández-Bedmar, Z., Font, R., del Río-Celestino, M., Pérez-Aparicio, J., Moreno-Ortega, A., Alonso-Moraga, Á., & Moreno-Rojas, R. (2019). Physicochemical Characterization and Biological Activities of Black and White Garlic: In Vivo and In Vitro Assays. Foods, 8(6), 220. https://doi.org/10.3390/foods8060220