Inhibition of the Maillard Reaction by Phytochemicals Composing an Aqueous Coffee Silverskin Extract via a Mixed Mechanism of Action

 ,
,  ,
, 


Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Coffee Silverskin Extract (CSE) Preparation
2.3. Phytochemical Characterisation of CSE
2.3.1. Total Phenolic Compounds (TPC)
2.3.2. Total Flavonoids Content (TFC)
2.3.3. Melatonin Quantification by High-Performance Liquid Chromatography-Electrospray Ionization Tandem Mass Spectrometry (HPLC-ESI-MS/MS)
2.3.4. CGA and Caffeine Quantification by Ultra-Performance Liquid Chromatography-Electrospray Ionization Tandem Mass Spectrometry (UPLC-ESI-MS/MS)
2.3.5. In Vitro Antioxidant Capacity
2.4. Maillard Reaction In Vitro Models: Protein Glycation and Formation of Methylglyoxal-Derivate AGE
2.5. Assessment of the Degree of Progress of the Maillard Reaction
2.5.1. Free Amino Groups
2.5.2. Early Products of the Maillard Reaction
2.5.3. Fluorescent AGE
2.5.4. Total AGE
2.5.5. Protein Oxidation
2.6. Evaluation of the Formation of Protein–Phytochemical Complexes
2.6.1. UV-Vis Spectra
2.6.2. Antioxidant Capacity of the Protein–Phytochemical Complexes
2.6.3. Fluorescence Spectra
2.6.4. In Silico Prediction of Protein–Phytochemical Interaction
2.7. Statistical Analysis
3. Results
3.1. Characterisation of CSE
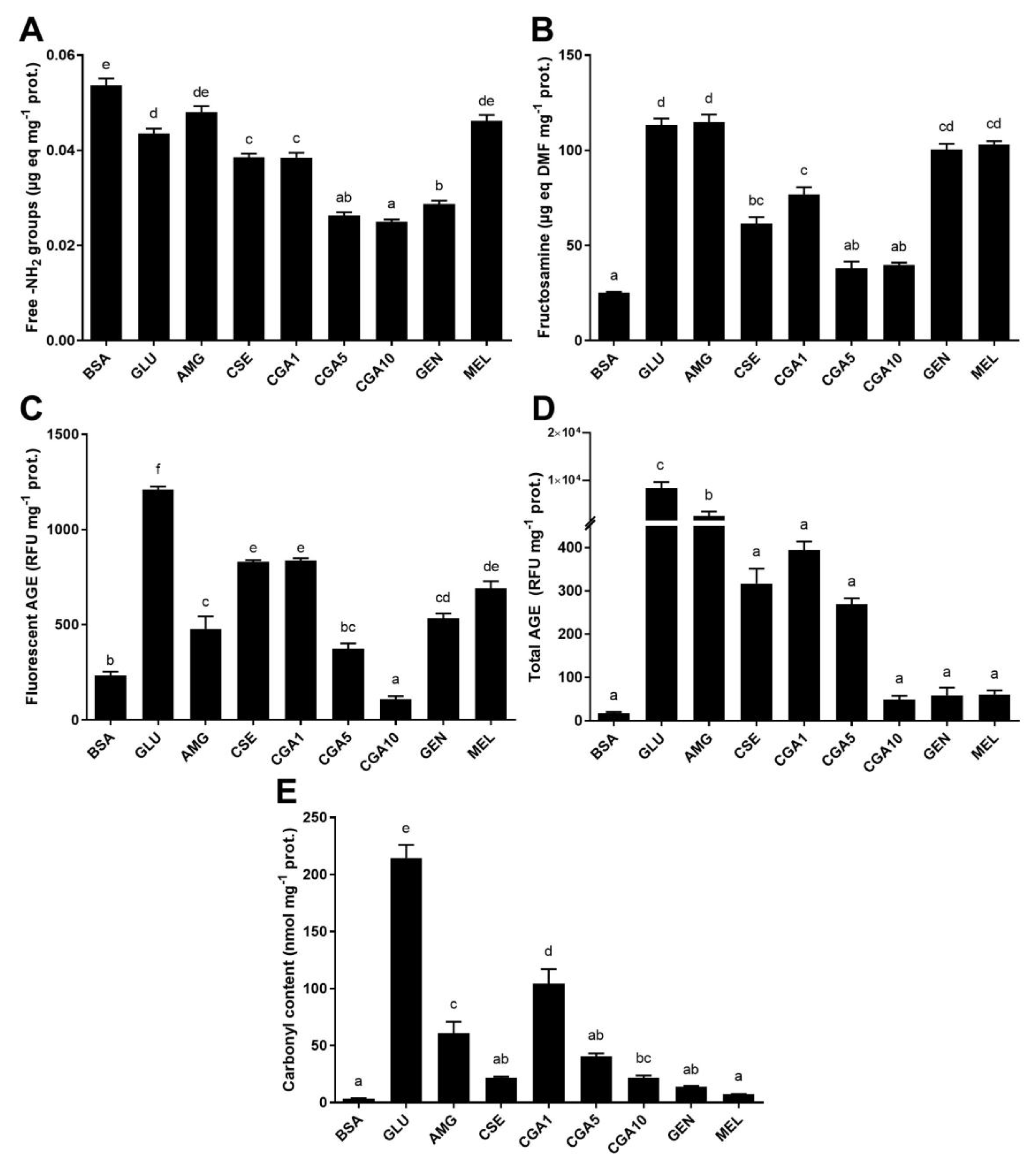
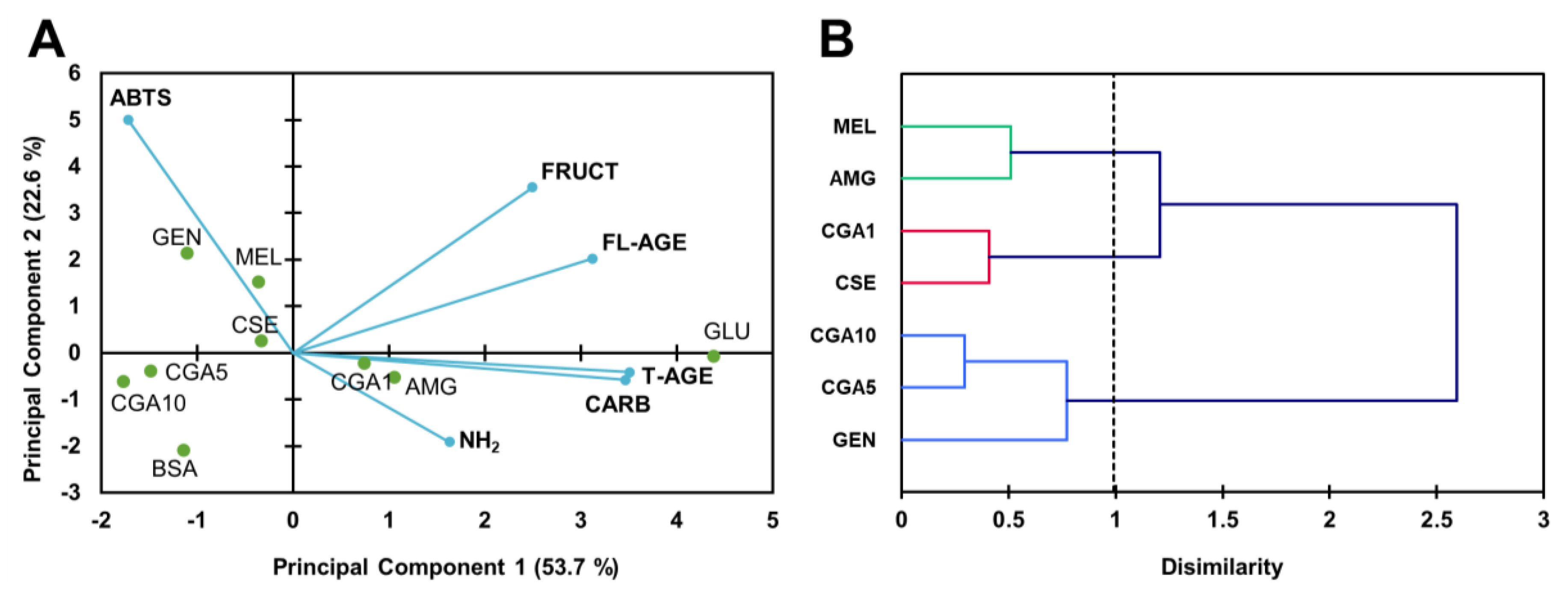
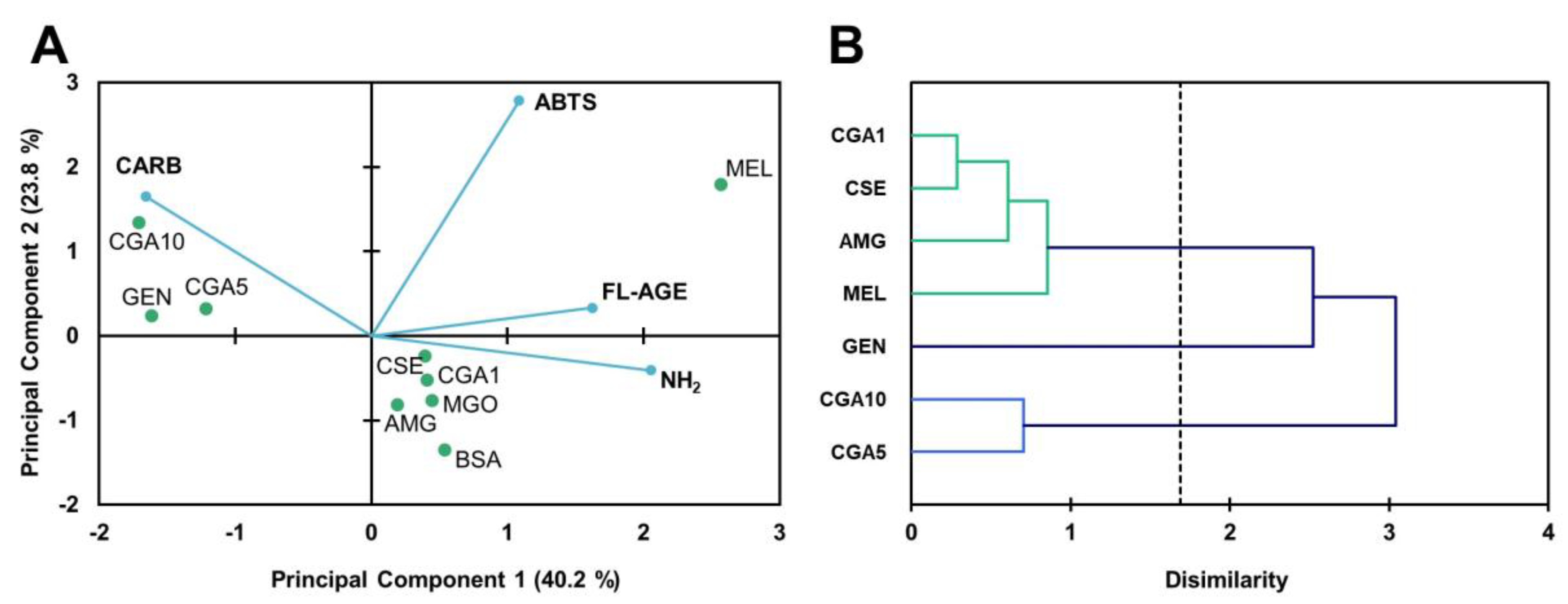
3.2. Inhibition of the Maillard Reaction
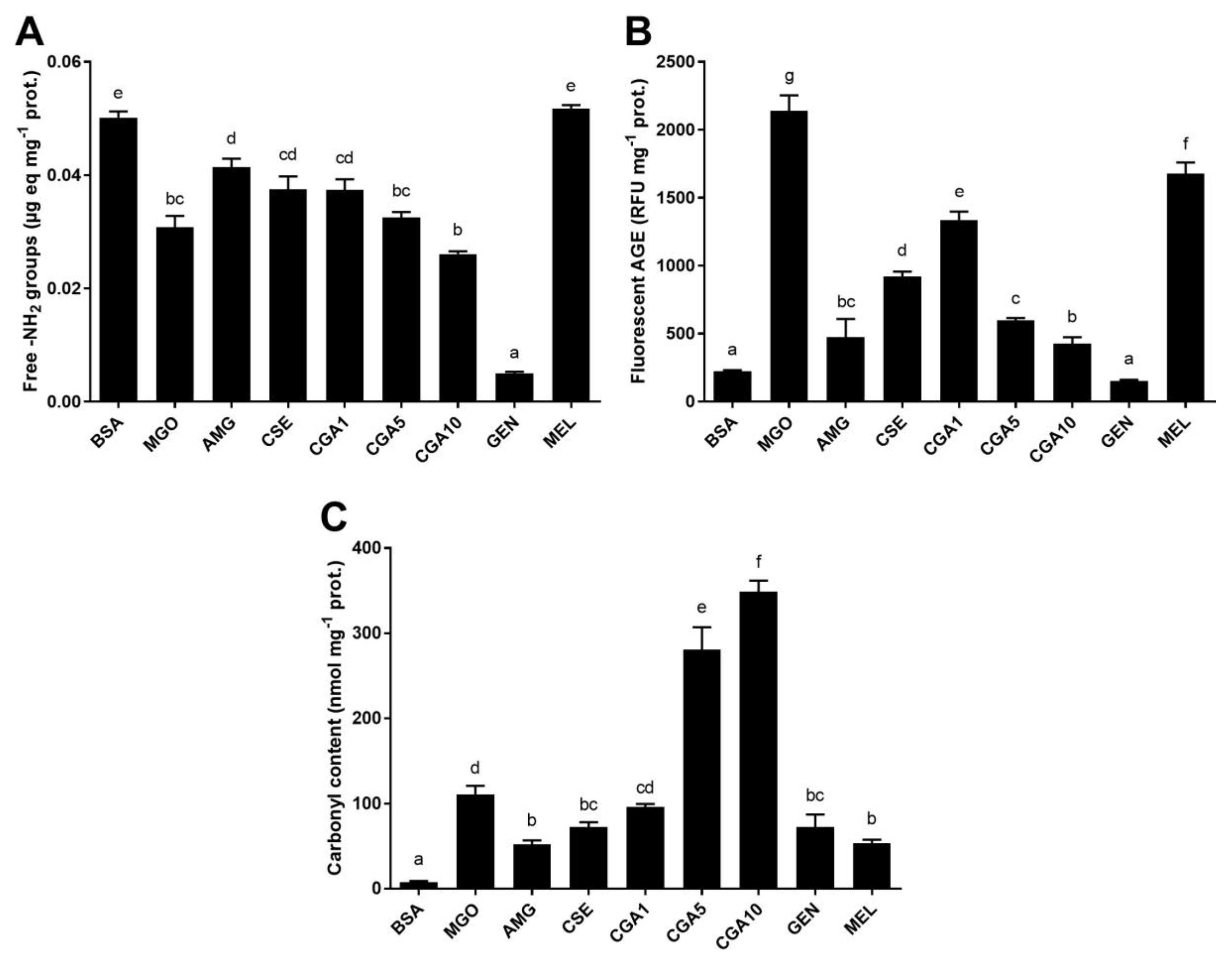
3.2.1. Protein Glycation in the BSA-GLU Model System
3.2.2. MGO-Derivative AGE Formation in the BSA-MGO Model System
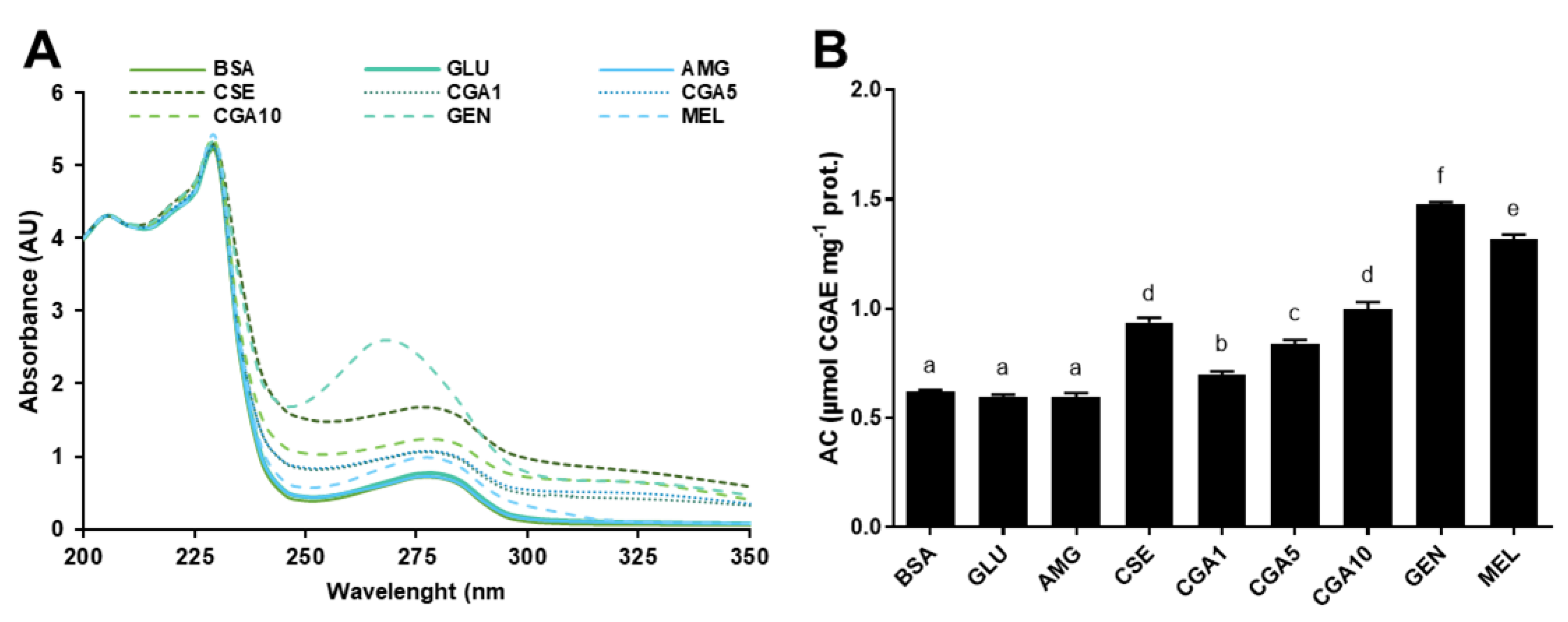
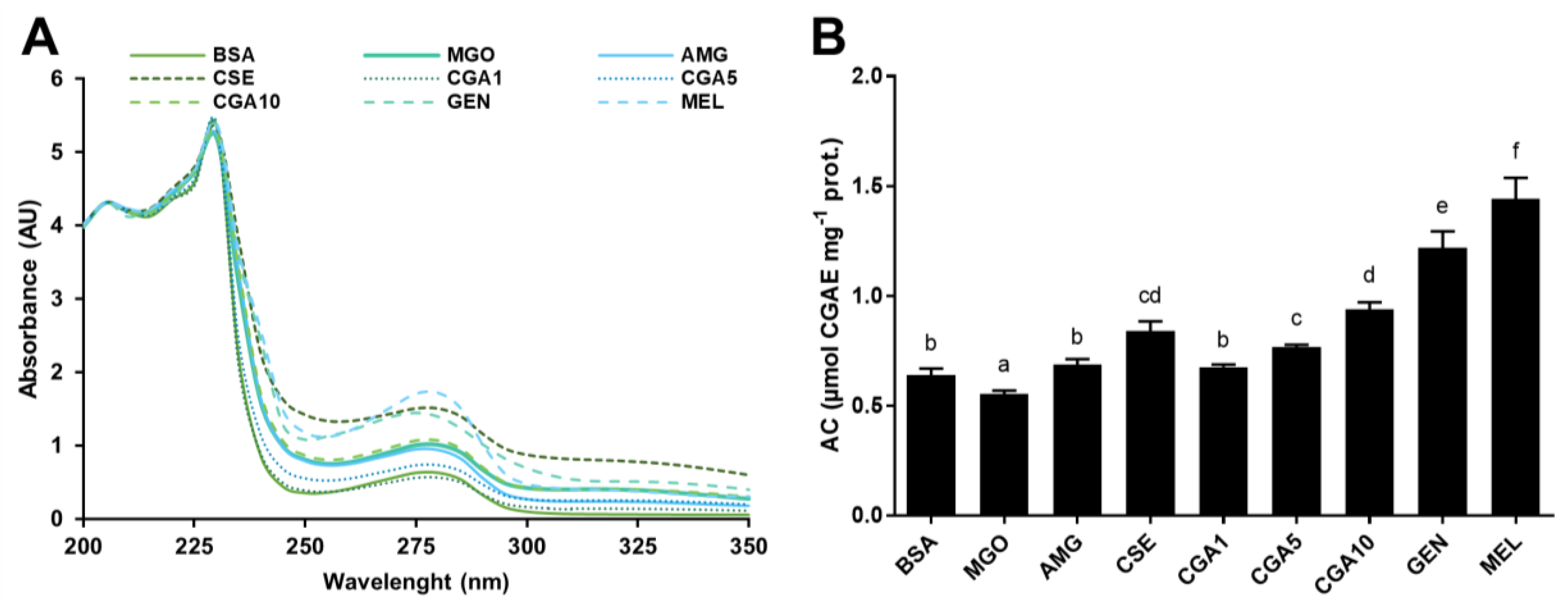
3.3. Formation of Protein–Phytochemical Complexes
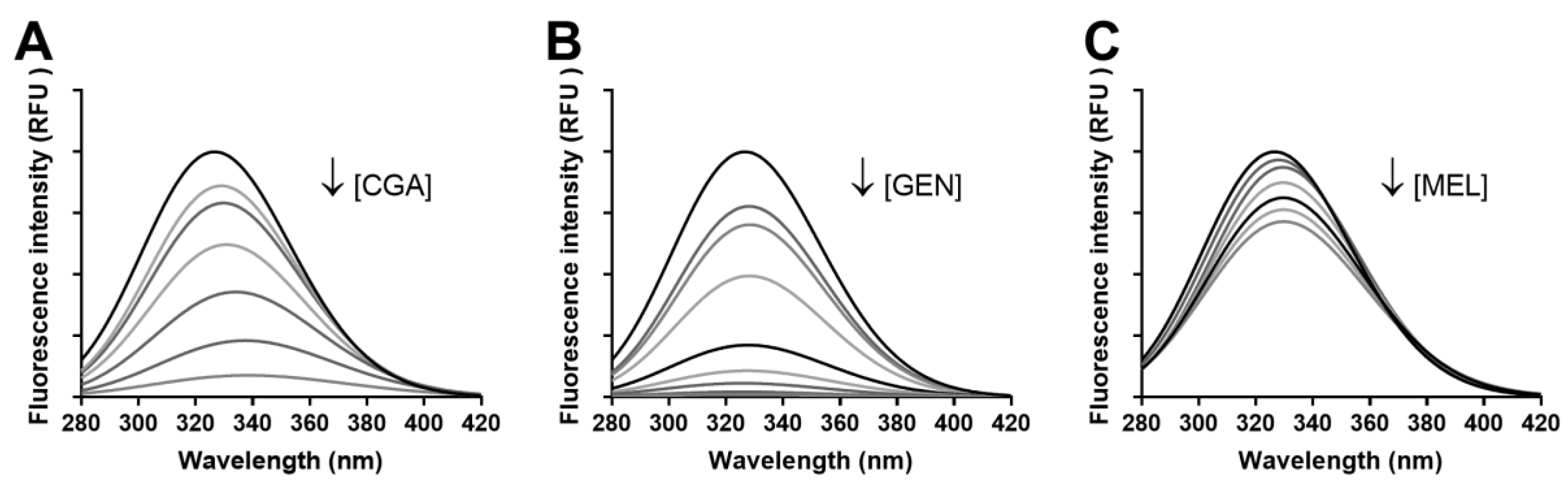
3.3.1. Fluorescence Spectra
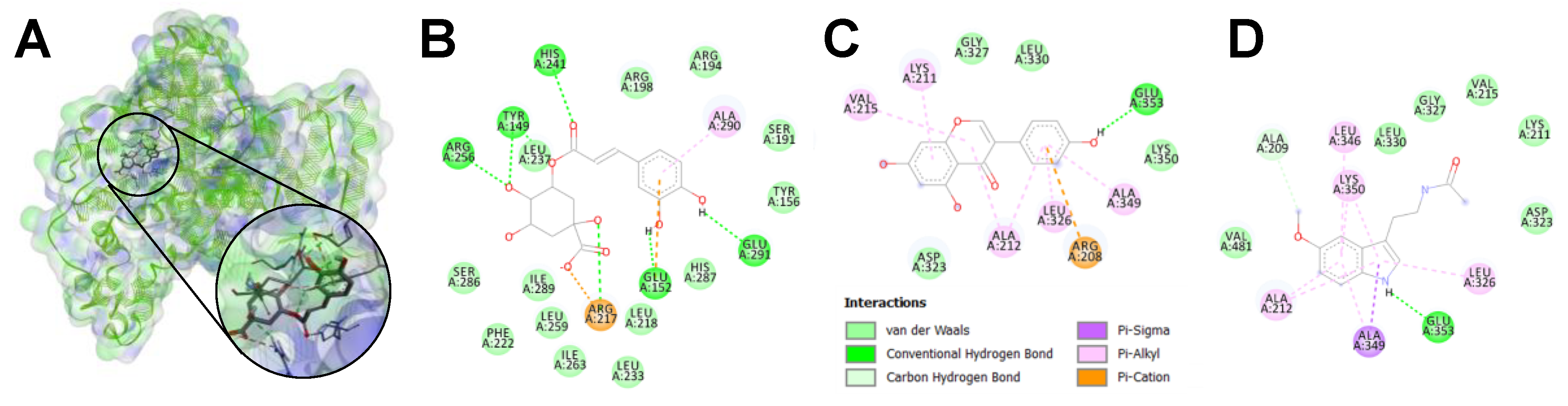
3.3.2. In Silico Prediction of Protein–Phytochemical Interactions
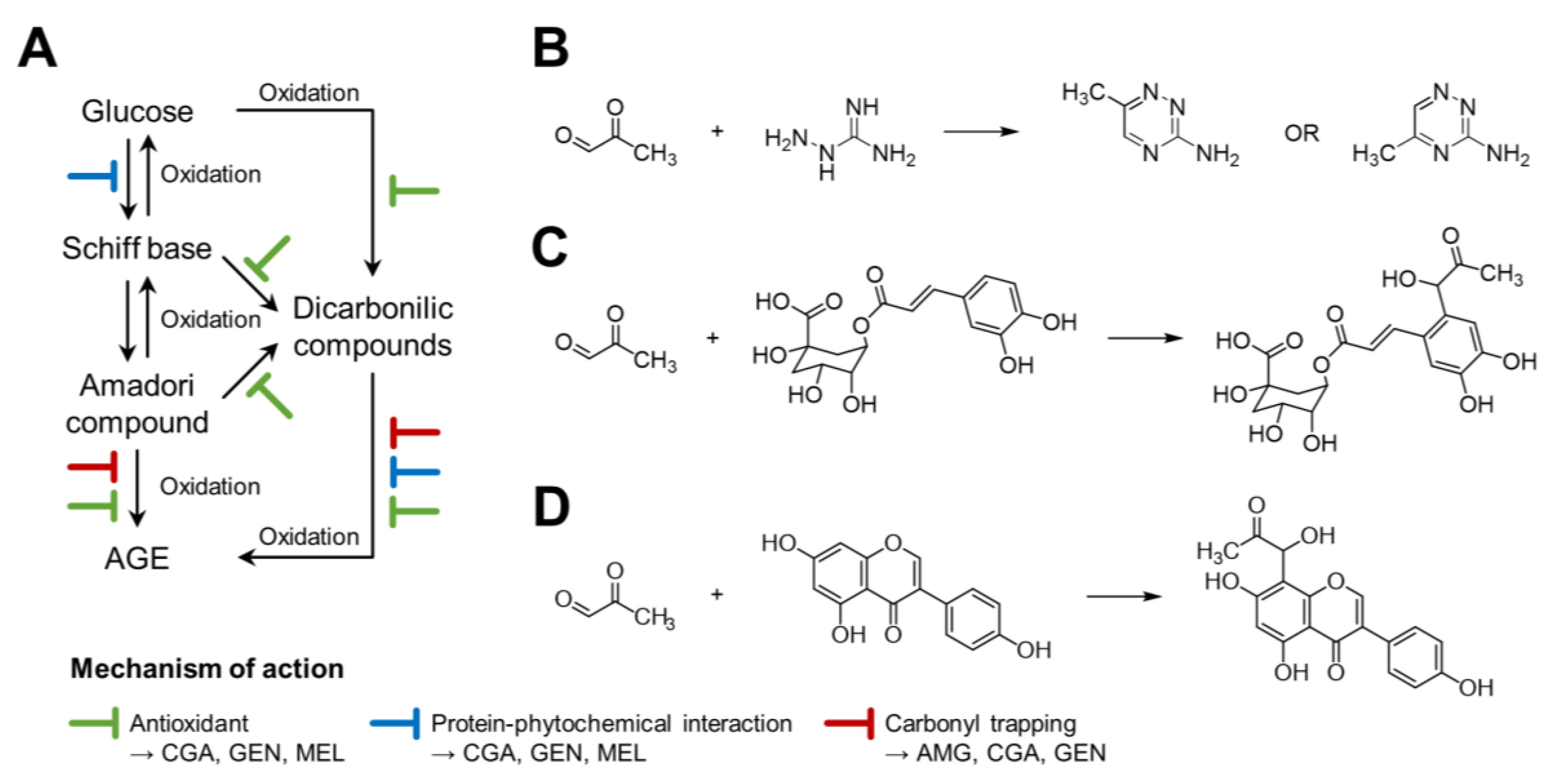
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ames, J.M. Evidence against dietary advanced glycation endproducts being a risk to human health. Mol. Nutr. Food Res. 2007, 51, 1085–1090. [Google Scholar] [CrossRef] [PubMed]
- Nowotny, K.; Jung, T.; Höhn, A.; Weber, D.; Grune, T. Advanced glycation end products and oxidative stress in type 2 diabetes mellitus. Biomolecules 2015, 5, 194–222. [Google Scholar] [CrossRef] [PubMed]
- Uribarri, J.; del Castillo, M.D.; de la Maza, M.P.; Filip, R.; Gugliucci, A.; Luevano-Contreras, C.; Macias-Cervantes, M.H.; Markowicz Bastos, D.H.; Medrano, A.; Menini, T.; et al. Dietary Advanced Glycation End Products and Their Role in Health and Disease. Adv. Nutr. An Int. Rev. J. 2015, 6, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Kellow, N.J.; Coughlan, M.T. Effect of diet-derived advanced glycation end products on inflammation. Nutr. Rev. 2015, 73, 737–759. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Saez, N.; Fernandez-Gomez, B.; Cai, W.; Uribarri, J.; del Castillo, M.D. In vitro formation of Maillard reaction products during simulated digestion of meal-resembling systems. Food Res. Int. 2019, 118, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.E.; Prasad, C.; Vijayagopal, P.; Juma, S.; Imrhan, V. Advanced Glycation End Products, Inflammation, and Chronic Metabolic Diseases: Links in a Chain? Crit. Rev. Food Sci. Nutr. 2016, 56, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Lund, M.N.; Ray, C.A. Control of Maillard Reactions in Foods: Strategies and Chemical Mechanisms. J. Agric. Food Chem. 2017, 65, 4537–4552. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Hedegaard, R.V.; Skibsted, L.H.; Andersen, M.L. Epicatechin and epigallocatechin gallate inhibit formation of intermediary radicals during heating of lysine and glucose. Food Chem. 2014, 146, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jongberg, S.; Andersen, M.L.; Davies, M.J.; Lund, M.N. Quinone-induced protein modifications: Kinetic preference for reaction of 1,2-benzoquinones with thiol groups in proteins. Free Radic. Biol. Med. 2016, 97, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-H.; Huang, S.-M.; Lin, J.-A.; Yen, G.-C. Inhibition of advanced glycation endproduct formation by foodstuffs. Food Funct. 2011, 2, 224. [Google Scholar] [CrossRef] [PubMed]
- Iriondo-DeHond, A.; Aparicio García, N.; Fernandez-Gomez, B.; Guisantes-Batan, E.; Velázquez Escobar, F.; Blanch, G.P.; San Andres, M.I.; Sanchez-Fortun, S.; del Castillo, M.D. Validation of coffee by-products as novel food ingredients. Innov. Food Sci. Emerg. Technol. 2019, 51, 194–204. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martin-Cabrejas, M.A.; Gonzalez de Mejia, E. Phenolic compounds from coffee by-products modulate adipogenesis-related inflammation, mitochondrial dysfunction, and insulin resistance in adipocytes, via insulin/PI3K/AKT signaling pathways. Food Chem. Toxicol. 2019, 132, 110672. [Google Scholar] [CrossRef] [PubMed]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martin-Cabrejas, M.A.; Gonzalez de Mejia, E. Relationship of the Phytochemicals from Coffee and Cocoa By-Products with their Potential to Modulate Biomarkers of Metabolic Syndrome In Vitro. Antioxidants 2019, 8, 279. [Google Scholar] [CrossRef] [PubMed]
- Mesías, M.; Navarro, M.; Martínez-Saez, N.; Ullate, M.; del Castillo, M.D.; Morales, F.J. Antiglycative and carbonyl trapping properties of the water soluble fraction of coffee silverskin. Food Res. Int. 2014, 62, 1120–1126. [Google Scholar] [CrossRef]
- Wang, P.; Chen, H.; Sang, S. Trapping Methylglyoxal by Genistein and Its Metabolites in Mice. Chem. Res. Toxicol. 2016, 29, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Shao, X.; Chen, H.; Ho, C.-T.; Sang, S. Genistein inhibits advanced glycation end product formation by trapping methylglyoxal. Chem. Res. Toxicol. 2011, 24, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Silvan, J.M.; Srey, C.; Ames, J.M.; Dolores del Castillo, M.; Silván, J.M.; Van de Lagemaat, J.; Olano, A.; del Castillo, M.D.; Van Nguyen, C.; Peng, X.F.; et al. Glycation is regulated by isoflavones. Food Funct. 2014, 5, 2036. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dontsov, A.E.; Vospelnikova, N.D.; Zack, P.P.; Ostrovsky, A.M.A. Antiglycation Activity of Melatonin. Dokl. Biochem. Biophys. 2017, 475, 283–286. [Google Scholar] [CrossRef]
- Del Castillo, M.D.; Ibañez, E.; Amigo-Benavent, M.; Herrero, M.; Plaza del Moral, M.; Ullate, M. Application of Products of Coffee Silverskin in Anti-ageing Cosmetics and Functional Food. WO 2013/004873, 10 January 2013. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Herrera, T.; Aguilera, Y.; Rebollo-Hernanz, M.; Bravo, E.; Benítez, V.; Martínez-Sáez, N.; Arribas, S.M.S.M.; del Castillo, M.D.M.D.; Martín-Cabrejas, M.A.M.A.M.A. Teas and herbal infusions as sources of melatonin and other bioactive non-nutrient components. LWT - Food Sci. Technol. 2018, 89, 65–73. [Google Scholar] [CrossRef]
- Aguilera, Y.; Herrera, T.; Liébana, R.; Rebollo-Hernanz, M.; Sanchez-Puelles, C.; Martín-Cabrejas, M.A. Impact of Melatonin Enrichment during Germination of Legumes on Bioactive Compounds and Antioxidant Activity. J. Agric. Food Chem. 2015, 63, 7967–7974. [Google Scholar] [CrossRef] [PubMed]
- Benitez, V.; Rebollo-Hernanz, M.; Hernanz, S.; Chantres, S.; Aguilera, Y.; Martin-Cabrejas, M.A. Coffee parchment as a new dietary fiber ingredient: Functional and physiological characterization. Food Res. Int. 2019, 122, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Saez, N.; García, A.T.A.T.; Pérez, I.D.I.D.; Rebollo-Hernanz, M.; Mesías, M.; Morales, F.J.F.J.; Martín-Cabrejas, M.A.M.A.M.A.; del Castillo, M.D.M.D. Use of spent coffee grounds as food ingredient in bakery products. Food Chem. 2017, 216, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Gomez, B.; Ramos, S.; Goya, L.; Mesa, M.D.; del Castillo, M.D.; Martín, M.Á. Coffee silverskin extract improves glucose-stimulated insulin secretion and protects against streptozotocin-induced damage in pancreatic INS-1E beta cells. Food Res. Int. 2016, 89, 1015–1022. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martin-Cabrejas, M.A.; Gonzalez de Mejia, E. Cocoa Shell Aqueous Phenolic Extract Preserves Mitochondrial Function and Insulin Sensitivity by Attenuating Inflammation Between Macrophages and Adipocytes in vitro. Mol. Nutr. Food Res. 2019, 63, 1801413. [Google Scholar] [CrossRef]
- Alonso-Salces, R.M.; Serra, F.; Reniero, F.; HÉberger, K. Botanical and Geographical Characterization of Green Coffee (Coffea arabica and Coffea canephora): Chemometric Evaluation of Phenolic and Methylxanthine Contents. J. Agric. Food Chem. 2009, 57, 4224–4235. [Google Scholar] [CrossRef]
- Farah, A.; de Paulis, T.; Trugo, L.C.; Martin, P.R. Effect of roasting on the formation of chlorogenic acid lactones in coffee. J. Agric. Food Chem. 2005, 53, 1505–1513. [Google Scholar] [CrossRef]
- Garrett, R.; Vaz, B.G.; Hovell, A.M.C.; Eberlin, M.N.; Rezende, C.M. Arabica and Robusta Coffees: Identification of Major Polar Compounds and Quantification of Blends by Direct-Infusion Electrospray Ionization–Mass Spectrometry. J. Agric. Food Chem. 2012, 60, 4253–4258. [Google Scholar] [CrossRef]
- Bertone, E.; Venturello, A.; Giraudo, A.; Pellegrino, G.; Geobaldo, F. Simultaneous determination by NIR spectroscopy of the roasting degree and Arabica/Robusta ratio in roasted and ground coffee. Food Control 2016, 59, 683–689. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Fernández-Gómez, B.; Herrero, M.; Sanchez-Puelles, C.; Aguilera, Y.; del Castillo, M.D.; Martin-Cabrejas, M.A. Effect of brewing process on the content of bioactive compounds (caffeine, chlorogenic acids and melatonin) of coffee brews. In Proceedings of the EuroFoodChem XVIII, Madrid, Spain, 13–16 October 2015; p. 192. [Google Scholar]
- Del Castillo, M.D.; Iriondo-DeHond, A.; Fernandez-Gomez, B.; Martinez-Saez, N.; Rebollo-Hernanz, M.; Martín-Cabrejas, M.A.; Farah, A. Coffee Antioxidants in Chronic Diseases. In Coffee: Consumption and Health Implications; Farah, A., Ed.; Royal Society of Chemistry: Cambridge, UK, 2019; pp. 20–56. [Google Scholar]
- Martín-Cabrejas, M.A.; Aguilera, Y.; Benítez, V.; Reiter, R.J. Melatonin Synthesis in Fermented Foods. In Fermented Foods in Health and Disease Prevention; Academic Press, Elsevier: Cambridge, MA, USA, 2017; pp. 105–129. ISBN 9780128023099. [Google Scholar]
- Fernandez-Gomez, B.; Ullate, M.; Picariello, G.; Ferranti, P.; Mesa, M.D.; del Castillo, M.D. New knowledge on the antiglycoxidative mechanism of chlorogenic acid. Food Funct. 2015, 6, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Echavarría, A.P.; Pagán, J.; Ibarz, A. Melanoidins Formed by Maillard Reaction in Food and Their Biological Activity. Food Eng. Rev. 2012, 4, 203–223. [Google Scholar] [CrossRef]
- Morales, F.J.; Somoza, V.; Fogliano, V. Physiological relevance of dietary melanoidins. Amino Acids 2012, 42, 1097–1109. [Google Scholar] [CrossRef] [PubMed]
- Sadowska-Bartosz, I.; Bartosz, G. Prevention of protein glycation by natural compounds. Molecules 2015, 20, 3309–3334. [Google Scholar] [CrossRef] [PubMed]
- Silván, J.M.; Assar, S.H.; Srey, C.; Dolores del Castillo, M.; Ames, J.M. Control of the Maillard reaction by ferulic acid. Food Chem. 2011, 128, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Chinchansure, A.A.; Korwar, A.M.; Kulkarni, M.J.; Joshi, S.P. Recent development of plant products with anti- glycation activity: a review. RSC Adv. 2015, 5, 31113–31138. [Google Scholar] [CrossRef]
- Gómez-Ojeda, A.; Jaramillo-Ortíz, S.; Wrobel, K.; Wrobel, K.; Barbosa-Sabanero, G.; Luevano-Contreras, C.; de la Maza, M.P.; Uribarri, J.; del Castillo, M.D.; Garay-Sevilla, M.E. Comparative evaluation of three different ELISA assays and HPLC-ESI-ITMS/MS for the analysis of Nε-carboxymethyl lysine in food samples. Food Chem. 2018, 243, 11–18. [Google Scholar] [CrossRef]
- Adisakwattana, S.; Sompong, W.; Meeprom, A.; Ngamukote, S.; Yibchok-Anun, S. Cinnamic acid and its derivatives inhibit fructose-mediated protein glycation. Int. J. Mol. Sci. 2012, 13, 1778–1789. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Lee, J.-A.; Kim, M.; Kum, H.; Jung, E.; Park, D. Anti-Glycation Activities of Phenolic Constituents from Silybum marianum (Milk Thistle) Flower in Vitro and on Human Explants. Molecules 2015, 20, 3549–3564. [Google Scholar] [CrossRef]
- Budryn, G.; Rachwal-Rosiak, D. Interactions of Hydroxycinnamic Acids with Proteins and Their Technological and Nutritional Implications. Food Rev. Int. 2013, 29, 217–230. [Google Scholar] [CrossRef]
- Kalinowska, M.; Bajko, E.; Matejczyk, M.; Kaczyński, P.; Łozowicka, B.; Lewandowski, W. The Study of Anti-/Pro-Oxidant, Lipophilic, Microbial and Spectroscopic Properties of New Alkali Metal Salts of 5-O-Caffeoylquinic Acid. Int. J. Mol. Sci. 2018, 19, 463. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Huang, Y.; Ma, X.; Liao, X.; Wang, Q.; Xiong, X.; Li, H. Multispectroscopic and docking studies on the binding of chlorogenic acid isomers to human serum albumin: Effects of esteryl position on affinity. Food Chem. 2016, 212, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Long, X.; Zeng, Y.-F.; Liu, Y.; Liu, Y.; Li, T.; Liao, L.; Guo, Y. Synthesis of novel genistein amino acid derivatives and investigation on their interactions with bovine serum albumin by spectroscopy and molecular docking. RSC Adv. 2018, 8, 31201–31212. [Google Scholar] [CrossRef]
- Prigent, S.V.; Voragen, A.G.; Visser, A.J.; van Koningsveld, G.A.; Gruppen, H. Covalent interactions between proteins and oxidation products of caffeoylquinic acid (chlorogenic acid). J. Sci. Food Agric. 2007, 87, 2502–2510. [Google Scholar] [CrossRef]
- Tang, D.; Li, H.-J.; Li, P.; Wen, X.-D.; Qian, Z.-M. Interaction of bioactive components caffeoylquinic Acid derivatives in Chinese medicines with bovine serum albumin. Chem. Pharm. Bull. (Tokyo) 2008, 56, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Gomez, B.; Nitride, C.; Ullate, M.; Mamone, G.; Ferranti, P.; del Castillo, M.D. Inhibitors of advanced glycation end products from coffee bean roasting by-product. Eur. Food Res. Technol. 2018, 244, 1101–1110. [Google Scholar] [CrossRef]
- Murthy, P.S.; Naidu, M.M. Recovery of Phenolic Antioxidants and Functional Compounds from Coffee Industry By-Products. Food Bioprocess Technol. 2012, 5, 897–903. [Google Scholar] [CrossRef]
- Gugliucci, A.; Bastos, D.H.M.; Schulze, J.; Souza, M.F.F. Caffeic and chlorogenic acids in Ilex paraguariensis extracts are the main inhibitors of AGE generation by methylglyoxal in model proteins. Fitoterapia 2009, 80, 339–344. [Google Scholar] [CrossRef]
- Kim, J.; Jeong, I.-H.; Kim, C.-S.; Lee, Y.M.; Kim, J.M.; Kim, J.S. Chlorogenic acid inhibits the formation of advanced glycation end products and associated protein cross-linking. Arch. Pharm. Res. 2011, 34, 495–500. [Google Scholar] [CrossRef]
- Edelstein, D.; Brownlee, M. Mechanistic studies of advanced glycosylation end product inhibition by aminoguanidine. Diabetes 1992, 41, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Nagai, R.; Murray, D.B.; Metz, T.O.; Baynes, J.W. Chelation: a fundamental mechanism of action of AGE inhibitors, AGE breakers, and other inhibitors of diabetes complications. Diabetes 2012, 61, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Fishman, S.L.; Sonmez, H.; Basman, C.; Singh, V.; Poretsky, L. The role of advanced glycation end-products in the development of coronary artery disease in patients with and without diabetes mellitus: A review. Mol. Med. 2018, 24, 59. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemicals | Concentration (mean ± SD) |
|---|---|
| TPC (μg CGAE mg−1 extract) | 70.4 ± 4.2 |
| CGA (μg mg−1 extract) | 35.1± 0.7 |
| TFC (μg RE mg−1 extract) | 83.2 ± 4.3 |
| Melatonin (ng mg−1 extract) | 2.0 ± 0.1 |
| Caffeine (μg mg−1 extract) | 47.0 ± 0.5 |
| ABTS•+ (μg CGAE mg−1 extract) | 166.5 ± 1.8 |
| Compound | KSV (104 mol L−1) | kq (mol L−1 s−1) | Ka (mol L−1) | n |
|---|---|---|---|---|
| Chlorogenic acid | 4.42 ± 0.14 | (7.64 ± 0.24) × 1012 | 4.78 ± 0.19 | 1.05 ± 0.04 |
| Genistein | 9.52 ± 0.10 | (1.65 ± 0.18) × 1013 | 5.36 ± 0.12 | 1.12 ± 0.03 |
| Melatonin | 0.70 ± 0.02 | (1.21 ± 0.34) × 1012 | 3.11 ± 0.18 | 0.82 ± 0.04 |
| BSA Domain | GLU | MGO | AMG | CGA | GEN | MEL | Interacting Amino Acids * |
|---|---|---|---|---|---|---|---|
| IA | ‒4.6 | ‒3.7 | ‒3.6 | ‒6.1 | ‒6.6 | ‒5.8 | Lys4, Arg10, Lys20, Lys41, Lys131, Lys132 |
| IB | ‒5.9 | ‒3.8 | ‒4.0 | ‒8.2 | ‒8.7 | ‒6.7 | Arg144, Arg194, Arg196, Arg198, Arg256, Lys431, Arg458 |
| IIA | ‒5.9 | ‒3.8 | ‒4.1 | ‒9.0 | ‒8.4 | ‒6.8 | Arg194, Arg198, Arg208, Lys211 Arg256, Lys350, Arg347, Arg484 |
| IIB | ‒5.4 | ‒3.3 | ‒4.0 | ‒7.9 | ‒8.4 | ‒6.9 | Arg208, Lys211, Lys350, Arg347, Arg483, Arg484 |
| IIIA | ‒5.4 | ‒3.5 | ‒4.1 | ‒8.1 | ‒8.0 | ‒7.5 | Arg217, Arg347, Arg409, Lys413, Lys431, Arg458, Arg484 |
| IIIB | ‒4.8 | ‒3.4 | ‒3.6 | ‒6.2 | ‒6.2 | ‒5.5 | Arg427, Arg431, Lys499, Lys524, Lys533 Lys556, Lys573 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rebollo-Hernanz, M.; Fernández-Gómez, B.; Herrero, M.; Aguilera, Y.; Martín-Cabrejas, M.A.; Uribarri, J.; del Castillo, M.D. Inhibition of the Maillard Reaction by Phytochemicals Composing an Aqueous Coffee Silverskin Extract via a Mixed Mechanism of Action. Foods 2019, 8, 438. https://doi.org/10.3390/foods8100438
Rebollo-Hernanz M, Fernández-Gómez B, Herrero M, Aguilera Y, Martín-Cabrejas MA, Uribarri J, del Castillo MD. Inhibition of the Maillard Reaction by Phytochemicals Composing an Aqueous Coffee Silverskin Extract via a Mixed Mechanism of Action. Foods. 2019; 8(10):438. https://doi.org/10.3390/foods8100438
Chicago/Turabian StyleRebollo-Hernanz, Miguel, Beatriz Fernández-Gómez, Miguel Herrero, Yolanda Aguilera, María A. Martín-Cabrejas, Jaime Uribarri, and María Dolores del Castillo. 2019. "Inhibition of the Maillard Reaction by Phytochemicals Composing an Aqueous Coffee Silverskin Extract via a Mixed Mechanism of Action" Foods 8, no. 10: 438. https://doi.org/10.3390/foods8100438
APA StyleRebollo-Hernanz, M., Fernández-Gómez, B., Herrero, M., Aguilera, Y., Martín-Cabrejas, M. A., Uribarri, J., & del Castillo, M. D. (2019). Inhibition of the Maillard Reaction by Phytochemicals Composing an Aqueous Coffee Silverskin Extract via a Mixed Mechanism of Action. Foods, 8(10), 438. https://doi.org/10.3390/foods8100438