Screening Method to Evaluate Amino Acid-Decarboxylase Activity of Bacteria Present in Spanish Artisanal Ripened Cheeses


Abstract
1. Introduction
2. Materials and Methods
2.1. Cheese Manufacturing
2.2. Strain Isolation
2.3. Preparation of Decarboxylase Media
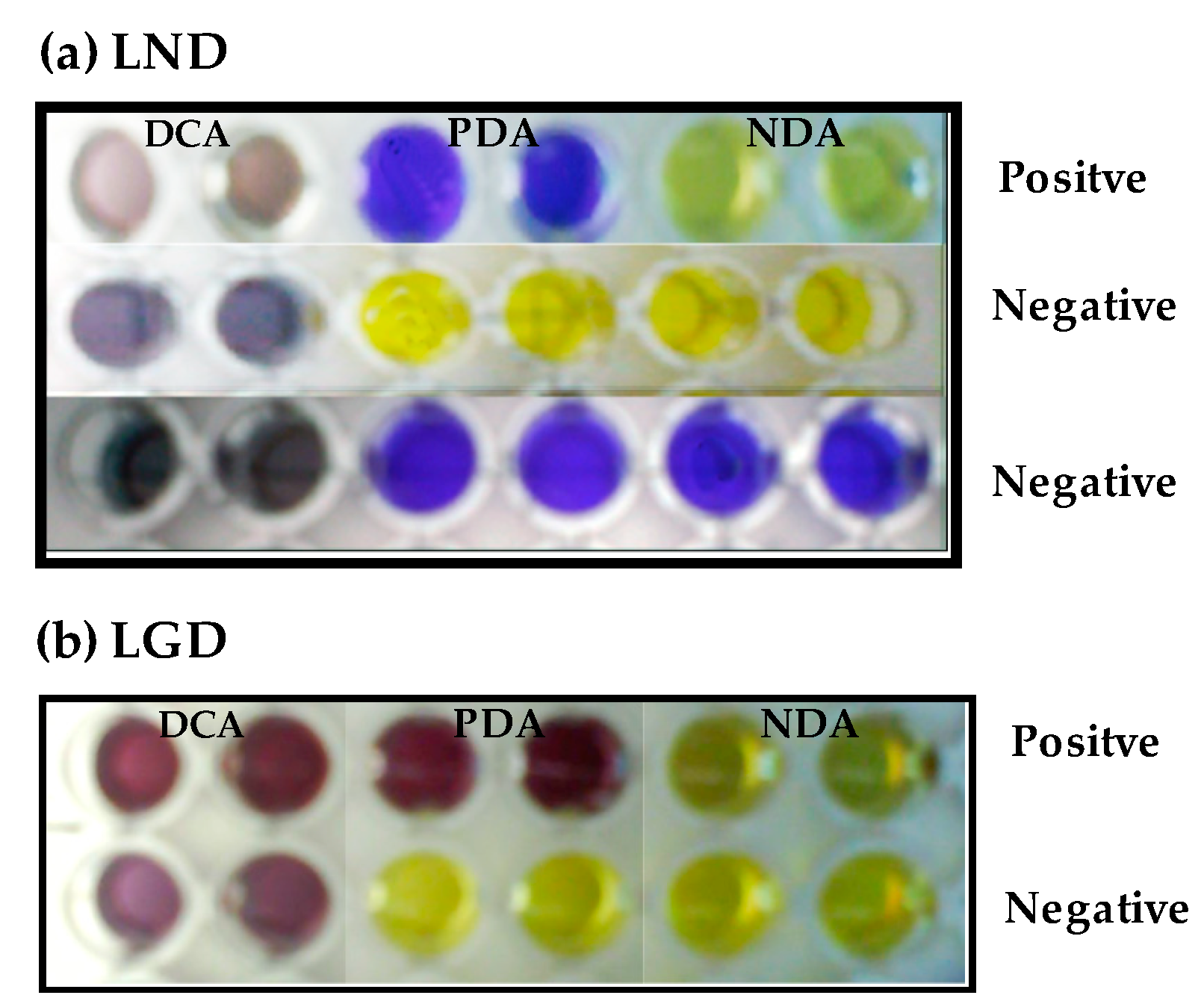
2.4. Assessment of Amino Acid Decarboxylase Activity
2.5. Confirmation of Amino Acid Decarboxylase Activity by HPLC
2.6. Analytical Validation of the Qualitative Microplate Method of Amino Acid Decarboxylase Activity
2.7. Identification of Strains with Decarboxylase Activity
3. Results and Discussion
3.1. Validation of the Qualitative Microplate Method of Amino Acid Decarboxylase Activity
3.2. Amino Acid Decarboxylase Activity of the Control Strains
3.3. Biogenic Amine Production by Isolates from Goat’s and Ewe’s Milk Cheeses
3.3.1. Total Amino-Acid Decarboxylase Activity of the Isolated Strains
3.3.2. Specific Amino Acid Decarboxylase Activity of the Isolated Strains
3.3.3. Identification of BA-Producing Strains
3.4. Consequences of HHP Treatments on the Formation of BA
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Karovicova, J.; Kohajdova, Z. Biogenic amines in food. ChemInform 2005, 36, 70–79. [Google Scholar] [CrossRef]
- Collins, J.D.; Noerrung, B.; Budka, H.; Andreoletti, O.; Buncic, S.; Griffin, J.; Hald, T.; Havelaar, A.; Hope, J.; Klein, G.; et al. Scientific Opinion on risk based control of biogenic amine formation in fermented foods. EFSA J. 2011, 9, 2393. [Google Scholar] [CrossRef]
- Costa, M.P.; Rodrigues, B.L.; Frasao, B.S.; Conte-junior, C.A. Chemical Risk for Human Consumption; Elsevier Inc.: Amsterdam, The Netherlands, 2018; ISBN 9780128114421. [Google Scholar]
- Benkerroum, N. Biogenic amines in dairy products: Origin, incidence, and control means. Compr. Rev. Food Sci. Food Saf. 2016, 15, 801–826. [Google Scholar] [CrossRef]
- Naila, A.; Flint, S.; Fletcher, G.; Bremer, P.; Meerdink, G. Control of biogenic amines in food-existing and emerging approaches. J. Food Sci. 2010, 75, R139–R150. [Google Scholar] [CrossRef] [PubMed]
- Novella-Rodriguez, S.; Veciana-Nogues, M.T.; Izquierdo-Pulido, M.; Vidal-Carou, M.C. Distribution of biogenic amines and polyamines in cheese. J. Food Sci. 2003, 68, 750–756. [Google Scholar] [CrossRef]
- Roig-Sagués, A.X.; Ruiz-Capillas, C.; Espinosa, D.; Hernández, M. The decarboxylating bacteria present in foodstuffs and the effect of emerging technologies on their formation. In Biological Aspects of Biogenic Amines, Polyamines and Conjugates; Dandrifosse, G., Ed.; Transworld Research Network: Trivandrum, India, 2009; pp. 201–230. [Google Scholar]
- Roig-Sagués, A.; Molina, A.; Hernández-Herrero, M. Histamine and tyramine-forming microorganisms in Spanish traditional cheeses. Eur. Food Res. Technol. 2002, 215, 96–100. [Google Scholar] [CrossRef]
- Hernández-Herrero, M.M.; Roig-Sagués, A.X.; Rodríguez-Jerez, J.J.; Mora-Ventura, M.T. Halotolerant and halophilic histamine-forming bacteria isolated during the ripening of salted anchovies (Engraulis encrasicholus). J. Food Prot. 1999, 62, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Kalac, P.; Abreu Gloria, M.B. Biogenic amine in cheeses, wines, beers and sauerkraut. In Biological Aspects of Biogenic Amines, Poliamines and Conjugates; Dandrifosse, G., Ed.; Transworld Research Network: Kerala, India, 2009; pp. 267–285. ISBN 9788178952499. [Google Scholar]
- Bover-Cid, S.; Hugas, M.; Izquierdo-Pulido, M.; Vidal-Carou, M.C. Amino acid-decarboxylase activity of bacteria isolated from fermented pork sausages. Int. J. Food Microbiol. 2001, 66, 185–189. [Google Scholar] [CrossRef]
- Ladero, V.; Fernández, M.; Calles-Enríquez, M.; Sánchez-Llana, E.; Cañedo, E.; Martín, M.C.; Alvarez, M.A. Is the production of the biogenic amines tyramine and putrescine a species-level trait in enterococci? Food Microbiol. 2012, 30, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Linares, D.M.; Martín, M.; Ladero, V.; Alvarez, M.A.; Fernández, M. Biogenic amines in dairy products. Crit. Rev. Food Sci. Nutr. 2011, 51, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Latorre-Moratalla, M.L.; Bover-Cid, S.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Control of biogenic amines in fermented sausages: Role of starter cultures. Front. Microbiol. 2012, 3, 169. [Google Scholar] [CrossRef] [PubMed]
- De las Rivas, B.; Ruiz-Capillas, C.; Carrascosa, A.V.; Curiel, J.A.; Jiménez-Colmenero, F.; Muñoz, R. Biogenic amine production by Gram-positive bacteria isolated from Spanish dry-cured “chorizo” sausage treated with high pressure and kept in chilled storage. Meat Sci. 2008, 80, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Novella-Rodríguez, S.; Veciana-Nogués, M.T.; Roig-Sagués, A.X.; Trujillo-Mesa, A.J.; Vidal-Carou, M.C. Influence of starter and nonstarter on the formation of biogenic amine in goat cheese during ripening. J. Dairy Sci. 2002, 85, 2471–2478. [Google Scholar] [CrossRef]
- Torracca, B.; Pedonese, F.; Turchi, B.; Fratini, F.; Nuvoloni, R. Qualitative and quantitative evaluation of biogenic amines in vitro production by bacteria isolated from ewes’ milk cheeses. Eur. Food Res. Technol. 2018, 244, 721–728. [Google Scholar] [CrossRef]
- Ladero, V.; Martín, M.C.; Redruello, B.; Mayo, B.; Flórez, A.B.; Fernández, M.; Alvarez, M.A. Genetic and functional analysis of biogenic amine production capacity among starter and non-starter lactic acid bacteria isolated from artisanal cheeses. Eur. Food Res. Technol. 2015, 241, 377–383. [Google Scholar] [CrossRef]
- Sumner, S.S.; Speckhard, M.W.; Somers, E.B.; Taylor, S.L. Isolation of histamine-producing Lactobacillus buchneri from Swiss cheese implicated in a food poisoning outbreak. Appl. Environ. Microbiol. 1985, 50, 1094–1096. [Google Scholar] [PubMed]
- Diaz, M.; del Rio, B.; Sanchez-Llana, E.; Ladero, V.; Redruello, B.; Fernández, M.; Martin, M.C.; Alvarez, M.A. Lactobacillus parabuchneri produces histamine in refrigerated cheese at a temperature-dependent rate. Int. J. Food Sci. Technol. 2018, 53, 2342–2348. [Google Scholar] [CrossRef]
- Pircher, A.; Bauer, F.; Paulsen, P. Formation of cadaverine, histamine, putrescine and tyramine by bacteria isolated from meat, fermented sausages and cheeses. Eur. Food Res. Technol. 2007, 226, 225–231. [Google Scholar] [CrossRef]
- Rodriguez-Jerez, J.J.; Giaccone, V.; Colavita, G.; Parisi, E. Bacillus macerans—A new potent histamine producing micro-organism isolated from Italian cheese. Food Microbiol. 1994, 11, 409–415. [Google Scholar] [CrossRef]
- Bover-Cid, S.; Holzapfel, W.H. Improved screening procedure for biogenic amine production by lactic acid bacteria. Int. J. Food Microbiol. 1999, 53, 33–41. [Google Scholar] [CrossRef]
- Marcobal, Á.; de las Rivas, B.; Moreno-Arribas, M.V.; Muñoz, R. Multiplex PCR method for the simultaneous detection of histamine-, tyramine-, and putrescine-producing lactic acid bacteria in foods. J. Food Prot. 2005, 68, 874–878. [Google Scholar] [CrossRef] [PubMed]
- Roig-Sagués, A.X.; Hernàndez-Herrero, M.M.; López-Sabater, E.I.; Rodríguez-Jerez, J.J.; Mora-Ventura, M.T. Evaluation of three decarboxylating agar media to detect histamine and tyramine-producing bacteria in ripened sausages. Lett. Appl. Microbiol. 1997, 25, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Maijala, R.L. Formation of histamine and tyramine by some lactic acid bacteria in MRS-broth and modified decarboxylation agar. Lett. Appl. Microbiol. 1993, 17, 40–43. [Google Scholar] [CrossRef]
- Espinosa-Pesqueira, D.; Hernández-Herrero, M.; Roig-Sagués, A.; Espinosa-Pesqueira, D.; Hernández-Herrero, M.M.; Roig-Sagués, A.X. High Hydrostatic Pressure as a tool to reduce formation of biogenic amines in artisanal Spanish cheeses. Foods 2018, 7, 137. [Google Scholar] [CrossRef] [PubMed]
- Eerola, S.; Hinkkanen, R.; Lindfors, E.; Hirvi, T. Liquid chromatographic determination of biogenic amines in dry sausages. J. AOAC Int. 1993, 76, 575–577. [Google Scholar] [PubMed]
- Altman, D.G.; Bland, J.M. Statistics Notes: Diagnostic tests 1: Sensitivity and specificity. BMJ 1994, 308, 1552. [Google Scholar] [CrossRef] [PubMed]
- Altman, D.G.; Bland, J.M. Statistics Notes: Diagnostic tests 2: Predictive values. BMJ 1994, 309, 102. [Google Scholar] [CrossRef] [PubMed]
- Altman, D.G.; Bland, J.M. Statistics Notes: Diagnostic tests 3: Receiver operating characteristic plots. BMJ 1994, 309, 188. [Google Scholar] [CrossRef] [PubMed]
- Zweig, M.H.; Campbell, G. Receiver-operating characteristic (ROC) plots: A fundamental evaluation tool in clinical medicine. Clin. Chem. 1993, 39, 561–577. [Google Scholar] [PubMed]
- Park, S.H.; Goo, J.M.; Jo, C.-H. Receiver Operating Characteristic (ROC) Curve: Practical review for radiologists. Korean J. Radiol. 2004, 5, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Hanley, J.A.; McNeil, B.J. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 1982, 143, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Harrigan, W.F. Laboratory Methods in Food Microbiology; Academic Press: Cambridge, MA, USA, 1998; ISBN 0123260434. [Google Scholar]
- Roig-Sagues, A.X.; Hernandez-Herrero, M.; Lopez-Sabater, E.I.; Rodriguez-Jerez, J.J.; Mora-Ventura, M.T. Histidine decarboxylase activity of bacteria isolated from raw and ripened salchichón, a Spanish cured sausage. J. Food Prot. 1996, 59, 516–520. [Google Scholar] [CrossRef]
- Joosten, H.M.; Northolt, M.D. Detection, growth, and amine-producing capacity of lactobacilli in cheese. Appl. Environ. Microbiol. 1989, 55, 2356–2359. [Google Scholar] [PubMed]
- Moreno-Arribas, M.V.; Polo, M.C.; Jorganes, F.; Muñoz, R. Screening of biogenic amine production by lactic acid bacteria isolated from grape must and wine. Int. J. Food Microbiol. 2003, 84, 117–123. [Google Scholar] [CrossRef]
- Marino, M.; Maifreni, M.; Bartolomeoli, I.; Rondinini, G. Evaluation of amino acid-decarboxylative microbiota throughout the ripening of an Italian PDO cheese produced using different manufacturing practices. J. Appl. Microbiol. 2008, 105, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Suzzi, G.; Gardini, F. Biogenic amines in dry fermented sausages: A review. Int. J. Food Microbiol. 2003, 88, 41–54. [Google Scholar] [CrossRef]
- Marino, M.; Maifreni, M.; Moret, S.; Rondinini, G. The capacity of Enterobacteriaceae species to produce biogenic amines in cheese. Lett. Appl. Microbiol. 2000, 31, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Halász, A.; Baráth, Á.; Simon-Sarkadi, L.; Holzapfel, W. Biogenic amines and their production by microorganisms in food. Trends Food Sci. Technol. 1994, 5, 42–49. [Google Scholar] [CrossRef]
- Martín, B.; Garriga, M.; Hugas, M.; Bover-Cid, S.; Veciana-Nogués, M.T.; Aymerich, T. Molecular, technological and safety characterization of Gram-positive catalase-positive cocci from slightly fermented sausages. Int. J. Food Microbiol. 2006, 107, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Pons-Sánchez-Cascado, S.; Vidal-Carou, M.C.; Mariné-Font, A.; Veciana-Nogués, M.T. Influence of the Freshness grade of raw fish on the formation of volatile and biogenic amines during the manufacture and storage of vinegar-marinated anchovies. J. Agric. Food Chem. 2005, 53, 8586–8592. [Google Scholar] [CrossRef] [PubMed]
- Masson, F.; Talon, R.; Montel, M.C. Histamine and tyramine production by bacteria from meat products. Int. J. Food Microbiol. 1996, 32, 199–207. [Google Scholar] [CrossRef]
- Santos, M.H. Amino acid decarboxylase capability of microorganisms isolated in Spanish fermented meat products. Int. J. Food Microbiol. 1998, 39, 227–230. [Google Scholar] [CrossRef]
- Drosinos, E.H.; Paramithiotis, S.; Kolovos, G.; Tsikouras, I.; Metaxopoulos, I. Phenotypic and technological diversity of lactic acid bacteria and staphylococci isolated from traditionally fermented sausages in Southern Greece. Food Microbiol. 2007, 24, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Foulquié Moreno, M.R.; Sarantinopoulos, P.; Tsakalidou, E.; De Vuyst, L. The role and application of enterococci in food and health. Int. J. Food Microbiol. 2006, 106, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Leuschner, R.G.K.; Kurihara, R.; Hammes, W.P. Formation of biogenic amines by proteolytic enterococci during cheese ripening. J. Sci. Food Agric. 1999, 79, 1141–1144. [Google Scholar] [CrossRef]
- Galgano, F.; Suzzi, G.; Favati, F.; Caruso, M.; Martuscelli, M.; Gardini, F.; Salzano, G. Biogenic amines during ripening in ‘Semicotto Caprino’ cheese: Role of enterococci. Int. J. Food Sci. Technol. 2001, 36, 153–160. [Google Scholar] [CrossRef]
- Landete, J.M.; Pardo, I.; Ferrer, S. Tyramine and phenylethylamine production among lactic acid bacteria isolated from wine. Int. J. Food Microbiol. 2007, 115, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Burdychova, R.; Komprda, T. Biogenic amine-forming microbial communities in cheese. FEMS Microbiol. Lett. 2007, 276, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Martuscelli, M.; Gardini, F.; Torriani, S.; Mastrocola, D.; Serio, A.; Chaves-López, C.; Schirone, M.; Suzzi, G. Production of biogenic amines during the ripening of Pecorino Abruzzese cheese. Int. Dairy J. 2005, 15, 571–578. [Google Scholar] [CrossRef]
- Tham, W.; Karp, G.; Danielsson-Tham, M.L. Histamine formation by enterococci in goat cheese. Int. J. Food Microbiol. 1990, 11, 225–229. [Google Scholar] [CrossRef]
- Perez, M.; Ladero, V.; del Rio, B.; Redruello, B.; de Jong, A.; Kuipers, O.; Kok, J.; Martin, M.C.; Fernandez, M.; Alvarez, M.A. The relationship among tyrosine decarboxylase and agmatine deiminase pathways in Enterococcus faecalis. Front. Microbiol. 2017, 8, 2107. [Google Scholar] [CrossRef] [PubMed]
- Novella-Rodriguez, S.; Veciana-Nogues, M.T.; Trujillo-Mesa, A.J.; Vidal-Carou, M.C. Profile of biogenic amines in goat cheese made from pasteurized and pressurized milks. J. Food Sci. 2002, 67, 2940–2944. [Google Scholar] [CrossRef]
- Fernández-García, E.; Tomillo, J.; Núñez, M. Effect of added proteinases and level of starter culture on the formation of biogenic amines in raw milk Manchego cheese. Int. J. Food Microbiol. 1999, 52, 189–196. [Google Scholar] [CrossRef]
- Sumner, S.S.; Taylor, S.L. Detection method for histamine-producing, dairy-related bacteria using diamine oxidase and leucocrystal violet. J. Food Prot. 1989, 52, 105–108. [Google Scholar] [CrossRef]
- Ascone, P.; Maurer, J.; Haldemann, J.; Irmler, S.; Berthoud, H.; Portmann, R.; Fröhlich-Wyder, M.-T.; Wechsler, D. Prevalence and diversity of histamine-forming Lactobacillus parabuchneri strains in raw milk and cheese—A case study. Int. Dairy J. 2017, 70, 26–33. [Google Scholar] [CrossRef]
- Kung, H.F.; Tsai, Y.H.; Hwang, C.C.; Lee, Y.H.; Hwang, J.H.; Wei, C.I.; Hwang, D.F. Hygienic quality and incidence of histamine-forming Lactobacillus species in natural and processed cheese in Taiwan. J. Food Drug Anal. 2005, 13, 51–56. [Google Scholar]
- Leuschner, R.G.; Heidel, M.; Hammes, W.P. Histamine and tyramine degradation by food fermenting microorganisms. Int. J. Food Microbiol. 1998, 39, 1–10. [Google Scholar] [CrossRef]
- Straub, B.W.; Kicherer, M.; Schilcher, S.M.; Hammes, W.P. The formation of biogenic amines by fermentation organisms. Z. Lebensm. Unters. Forsch. 1995, 201, 79–82. [Google Scholar] [CrossRef] [PubMed]
- González de Llano, D.; Ramos, M.; Rodriguez, A.; Montilla, A.; Juarez, M. Microbiological and physicochemical characteristics of Gamonedo blue cheese during ripening. Int. Dairy J. 1992, 2, 121–135. [Google Scholar] [CrossRef]
- Fontecha, J.; Peláez, C.; Juárez, M.; Requena, T.; Gómez, C.; Ramos, M. Biochemical and microbiological characteristics of artisanal hard goat’s cheese. J. Dairy Sci. 1990, 73, 1150–1157. [Google Scholar] [CrossRef]


{kind=link}
| Reagent | Low Nitrogen Decarboxylase Broth (LND) | Low Glucose Decarboxylase Broth (LGD) | Adjusted pH |
|---|---|---|---|
| Tryptone | 0.125 | 0.25 | |
| Yeast extract | 0.125 | 0.25 | |
| NaCl | 0.25 | 0.25 | |
| CaCO3 | 0.01 | 0.01 | |
| Pyridoxal-5-phosphate | 0.03 | 0.03 | |
| Glucose | 0.05 | 0.001 | |
| Bromocresol purple | 0.01 | 0.01 | |
| All amino acids | 1.0 | 1.0 | 5.5 |
| l-Lysine | 1.0 | 1.0 | 5.0 |
| l-Ornithine | 1.0 | 1.0 | 5.5 |
| l-Histidine | 1.0 | 1.0 | 5.7 |
| l-Tyrosine | 0.25 | 0.25 | 5.5 |
| Broth | Data of ROC Analysis | Ornithine | Lysine | Histidine | Tyrosine |
|---|---|---|---|---|---|
| LND | AUC | 0.999 (0.974–1.00) | 0.992 (0.961–1.00) | 0.737 (0.659–0.806) | 0.980 (0.943–0.996) |
| Optimal cut-off (mg/L) | 10 | 15 | 50 | 25 | |
| Sensitivity at optimal cut-off (%) | 98.53 | 98.68 | 65.38 | 92.25 | |
| Specificity at optimal cut-off (%) | 100 | 93.15 | 91.75 | 100 | |
| PPV (%) | 100 | 83.7 | 81 | 100 | |
| NPV (%) | 99.0 | 98.6 | 83.2 | 66.7 | |
| FN at cut-off | 3 | 6 | 11 | 1 | |
| FP at cut-off | 1 | 1 | 17 | 6 | |
| LGD | AUC | 0.907 (0.849–0.948) | 0.935 (0.883–0.969) | 0.592 (0.509–0.672) | 0.989 (0.956–0.999) |
| Optimal cut-off (mg/L) | 15 | 10 | 50 | 20 | |
| Sensitivity at optimal cut-off (%) | 84.37 | 88 | 30 | 93.06 | |
| Specificity at optimal cut-off (%) | 97.5 | 97.6 | 100 | 100 | |
| PPV (%) | 90 | 88 | 100 | 100 | |
| NPV (%) | 95.8 | 97.6 | 90.3 | 37.5 | |
| FN at cut-off | 5 | 5 | 2 | 0 | |
| FP at cut-off | 5 | 3 | 14 | 8 |
| Strain | PU | CA | HI | TY | |||||
|---|---|---|---|---|---|---|---|---|---|
| Broth | DMA | HPLC | DMA | HPLC | DMA | HPLC | DMA | HPLC | |
| L. brevis | LGD | (−) | 1.11 | (−) | ND | ND | ND | (+) | 109.8 |
| L. casei | LGD | (−) | 0.78 | (−) | ND | ND | ND | (+) | 77.14 |
| S. epidermidis | LND | (−) | 0.85 | (−) | ND | (+) | 46.48 | (−) | ND |
| Medium | BAP | PU | CA | HI | TY | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ± | + | ++ | +++ | ± | + | ++ | +++ | ± | + | ++ | +++ | ± | + | ++ | +++ | ||
| VRBG | 25 | 1 | 2 | 17 | 4 | 0 | 0 | 17 | 7 | 8 | 10 | 4 | 0 | 3 | 6 | 9 | 0 |
| BPA | 15 | 1 | 1 | 3 | 3 | 0 | 0 | 4 | 3 | 4 | 4 | 1 | 0 | 2 | 2 | 7 | 0 |
| KF | 89 | 5 | 3 | 22 | 4 | 13 | 0 | 22 | 9 | 19 | 21 | 4 | 0 | 4 | 8 | 71 | 0 |
| M-17 | 98 | 7 | 2 | 5 | 0 | 7 | 1 | 2 | 1 | 14 | 6 | 2 | 0 | 5 | 6 | 69 | 0 |
| MRS | 72 | 2 | 2 | 2 | 3 | 0 | 0 | 2 | 3 | 7 | 0 | 0 | 0 | 5 | 8 | 50 | 0 |
| Total | 299 | 16 | 10 | 49 | 14 | 20 | 1 | 47 | 23 | 52 | 41 | 11 | 0 | 19 | 30 | 206 | 0 |
| Identification | N | PU | CA | HI | TY | ||||
|---|---|---|---|---|---|---|---|---|---|
| Gram Negative | |||||||||
| Hafnia alvei | 4 | 4 | 537.7–889.54 | 2 | 641–1001.30 | 4 | 30.43–95.68 | 4 | 22.44–151.54 |
| Enterobacteriaceae | 1 | 1 | 1037.52 | 1 | 1173.24 | 1 | 111.43 | 1 | 73.41 |
| Gram Positive | |||||||||
| Staphylococcus cohni subsp. cohni | 1 | - | 1 | 1.50 | 1 | 1.57 | 1 | 25.78 | |
| Staphylococcus warneri | 2 | 2 | 69.29–753.55 | 2 | 240.94–694.22 | 1 | 88.63 | 1 | 65.50 |
| Staphylococcus capitis | 1 | 1 | 310.30 | 1 | 246.37 | 1 | 95.13 | - | |
| Staphylococcus lentus | 2 | 1 | 23.44 | 2 | 5.41–19 | - | 2 | 39.58–191.69 | |
| Staphylococcus hominis | 2 | 1 | 890.96 | 1 | 998.20 | 1 | 91.63 | 2 | 102.42–245.22 |
| Enterococcus faecalis | 8 | 8 | 39.66–884.27 | 8 | 32.44–972 | 6 | 25.62–92.35 | 8 | 327.5–477.20 |
| Enterococcus durans | 2 | - | - | - | 2 | 337.40–357.44 | |||
| Enterococcus avium | 1 | - | 1 | 26.52 | 1 | 56.51 | 1 | 24.88 | |
| Enterococcus faecium | 6 | 3 | 19.68–1113.80 | 3 | 1.32–1281.50 | 3 | 20.22–111.38 | 6 | 9.91–366.47 |
| Lactococcus lactis subsp. lactis | 10 | 1 | 7.25 | 1 | 7.50 | 2 | 21.24–33.38 | 10 | 198.77–450.77 |
| Pediococcus pentosaceus | 1 | - | - | 1 | 33.44 | 1 | 55.71 | ||
| Lactobacillus brevis | 5 | - | - | 1 | 22.51 | 5 | 212.58–519.52 | ||
| Lactobacillus plantarum | 3 | 1 | 22.41 | - | 1 | 24.5 | 3 | 307.62–528.45 | |
| Lactobacillus paracasei subps. paracasei | 1 | - | - | - | 1 | 307.03 | |||
| Leuconostoc spp. | 3 | 2 | 252.88–749.05 | 2 | 14.47–28.83 | 2 | 22.74–25.75 | 3 | 330.01–417 |
| Bacillus macerans | 1 | - | - | - | 1 | 418.10 | |||
| Bacillus licheniformis | 1 | - | - | - | 1 | 403.07 | |||
| Identification | N | PU | CA | HI | TY | ||||
|---|---|---|---|---|---|---|---|---|---|
| Gram Negative | |||||||||
| Escherichia coli | 2 | 2 | 746–857.48 | 2 | 762.35–983.93 | 2 | 41.37–74.12 | 2 | 21.7–281.48 |
| Hafnia alvei | 5 | 5 | 738.50–1049.51 | 5 | 787.20–1180.12 | 4 | 43.52–185.54 | 5 | 48.83–184.79 |
| Klebsiella oxytoca | 2 | 2 | 30.63–458.50 | 2 | 493.41–866.96 | 2 | 16.04–27.22 | 2 | 5.01–167.24 |
| Citrobacter freundii | 1 | 1 | 67.1 | 1 | 1095.3 | 1 | 45.62 | 1 | 372.38 |
| Enterobacteriaceae | 1 | 1 | 832.76 | 1 | 846.95 | 1 | 83.95 | 1 | 69.42 |
| Gram Positive | |||||||||
| Staphylococcus xylosus | 2 | - | - | 2 | 42.31–68.5 | 2 | 92.09–475.35 | ||
| Staphylococcus chromogenes | 1 | 1 | 1142.92 | 1 | 1760.06 | 1 | 31.12 | 1 | 441.40 |
| Staphylococcus aureus | 1 | - | - | 1 | 100.54 | - | |||
| Enterococcus faecalis | 5 | 3 | 860.29–978.9 | 4 | 35.43–1394.87 | 5 | 20.7–92.98 | 5 | 338.01–461.50 |
| Enterococcus durans | 9 | 9 | 12.63–1160.44 | 8 | 13.32–1773.03 | 9 | 30.8–179.32 | 9 | 46.09–747.33 |
| Enterococcus faecium | 2 | 2 | 24.89–847.05 | 2 | 877.24–941.69 | 2 | 28.98–29.97 | 2 | 100.54–149.41 |
| Enterococcus hirae | 2 | 2 | 552.4–579.25 | 2 | 600.67–615.75 | 2 | 32.36–45.95 | 2 | 108.01–187.58 |
| Enterococcus avium | 2 | - | 1 | 15.17 | 1 | 2.18 | 2 | 19.93–434.25 | |
| Streptococcus salivarius | 1 | - | - | - | 1 | 742.1 | |||
| Lactococcus lactis subsp. lactis | 9 | 3 | 34.39–795.26 | 2 | 14.95–844.13 | 2 | 16.38–30.72 | 9 | 211.93–566.2 |
| Lactococcus lactis subsp. cremoris | 4 | 3 | 4.78–37.63 | 3 | 5.23–22.94 | 2 | 11.15–19.03 | 4 | 229.47–406.87 |
| Pediococcus pentosaceus | 4 | 3 | 9.91–897.67 | 3 | 10.25–1018.4 | 3 | 27.06–88.78 | 4 | 34.58–411.66 |
| Lactobacillus paracasei subsp. paracasei | 6 | 2 | 17.42–45.05 | 1 | 17.18 | 1 | 51.93 | 6 | 365.79–575.73 |
| Lactobacillus plantarum | 4 | 1 | 8.58 | 1 | 17.58 | 1 | 26.26 | 4 | 45.1–353.31 |
| Lactobacillus brevis | 1 | - | - | - | 1 | 240.08 | |||
| Lactobacillus pentosus | 1 | 1 | 50.79 | - | - | 1 | 422.75 | ||
| Leuconostoc spp. | 5 | 2 | 11.13–1162.76 | 2 | 22.52–1781.26 | 3 | 29.63–33.82 | 5 | 392.01–626.42 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espinosa-Pesqueira, D.; Roig-Sagués, A.X.; Hernández-Herrero, M.M. Screening Method to Evaluate Amino Acid-Decarboxylase Activity of Bacteria Present in Spanish Artisanal Ripened Cheeses. Foods 2018, 7, 182. https://doi.org/10.3390/foods7110182
Espinosa-Pesqueira D, Roig-Sagués AX, Hernández-Herrero MM. Screening Method to Evaluate Amino Acid-Decarboxylase Activity of Bacteria Present in Spanish Artisanal Ripened Cheeses. Foods. 2018; 7(11):182. https://doi.org/10.3390/foods7110182
Chicago/Turabian StyleEspinosa-Pesqueira, Diana, Artur X. Roig-Sagués, and M. Manuela Hernández-Herrero. 2018. "Screening Method to Evaluate Amino Acid-Decarboxylase Activity of Bacteria Present in Spanish Artisanal Ripened Cheeses" Foods 7, no. 11: 182. https://doi.org/10.3390/foods7110182
APA StyleEspinosa-Pesqueira, D., Roig-Sagués, A. X., & Hernández-Herrero, M. M. (2018). Screening Method to Evaluate Amino Acid-Decarboxylase Activity of Bacteria Present in Spanish Artisanal Ripened Cheeses. Foods, 7(11), 182. https://doi.org/10.3390/foods7110182