Lutein Esterification in Wheat Flour Increases the Carotenoid Retention and Is Induced by Storage Temperatures


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Sample Preparation and Storage Conditions
2.2. Chemicals and Reagents
2.3. Extraction of Carotenoids
2.4. HPLC Analysis of Carotenoids
2.5. Degradation Kinetics Model
2.6. Statistical Analysis
3. Results and Discussion
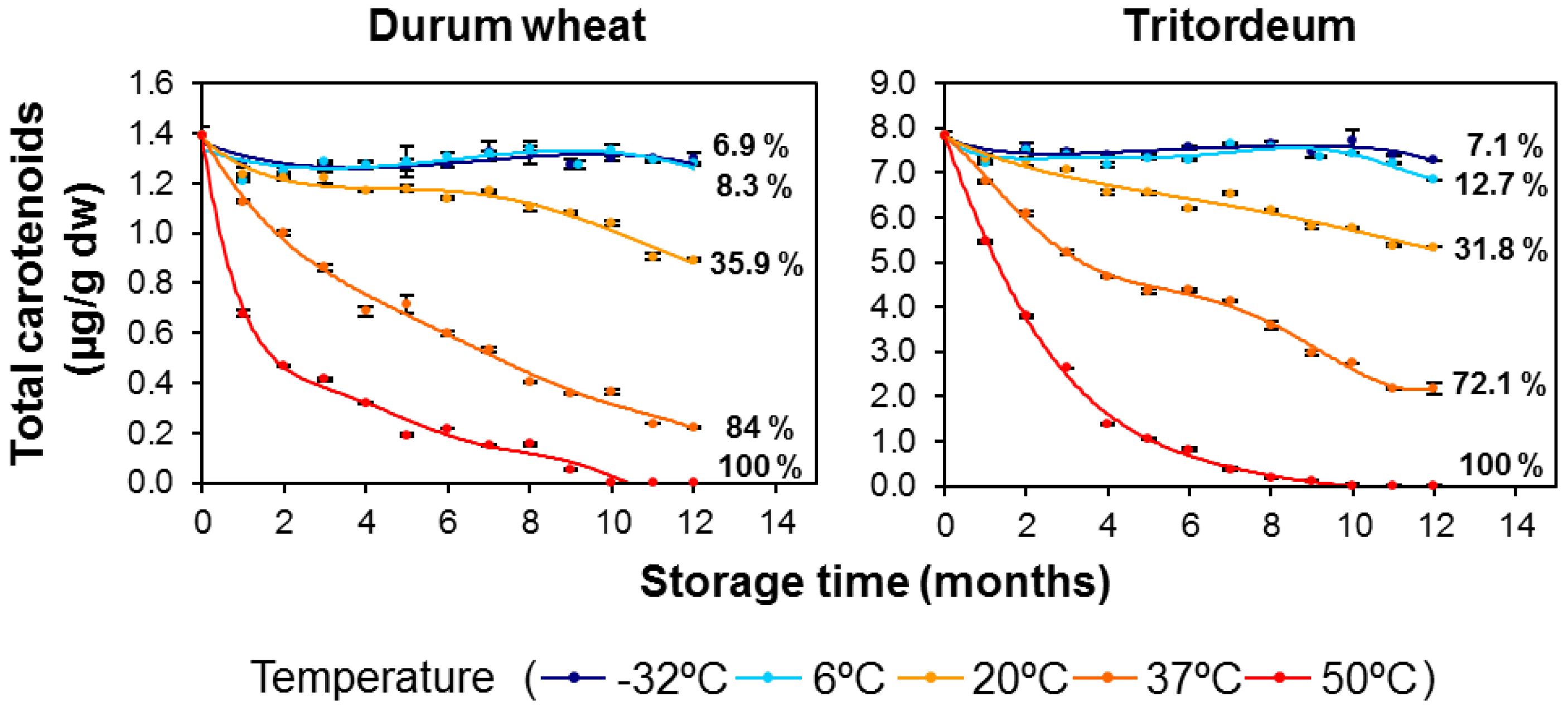
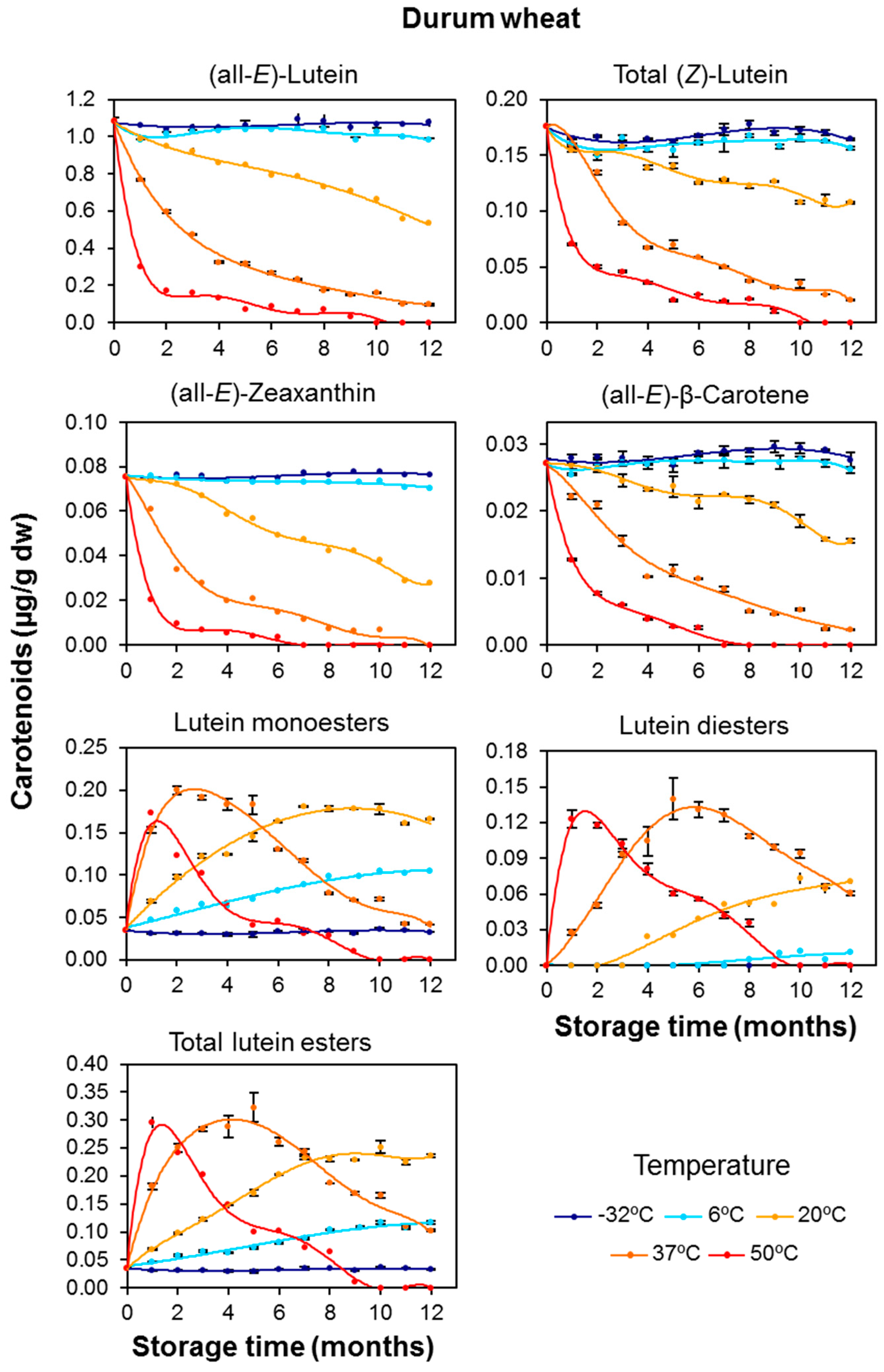
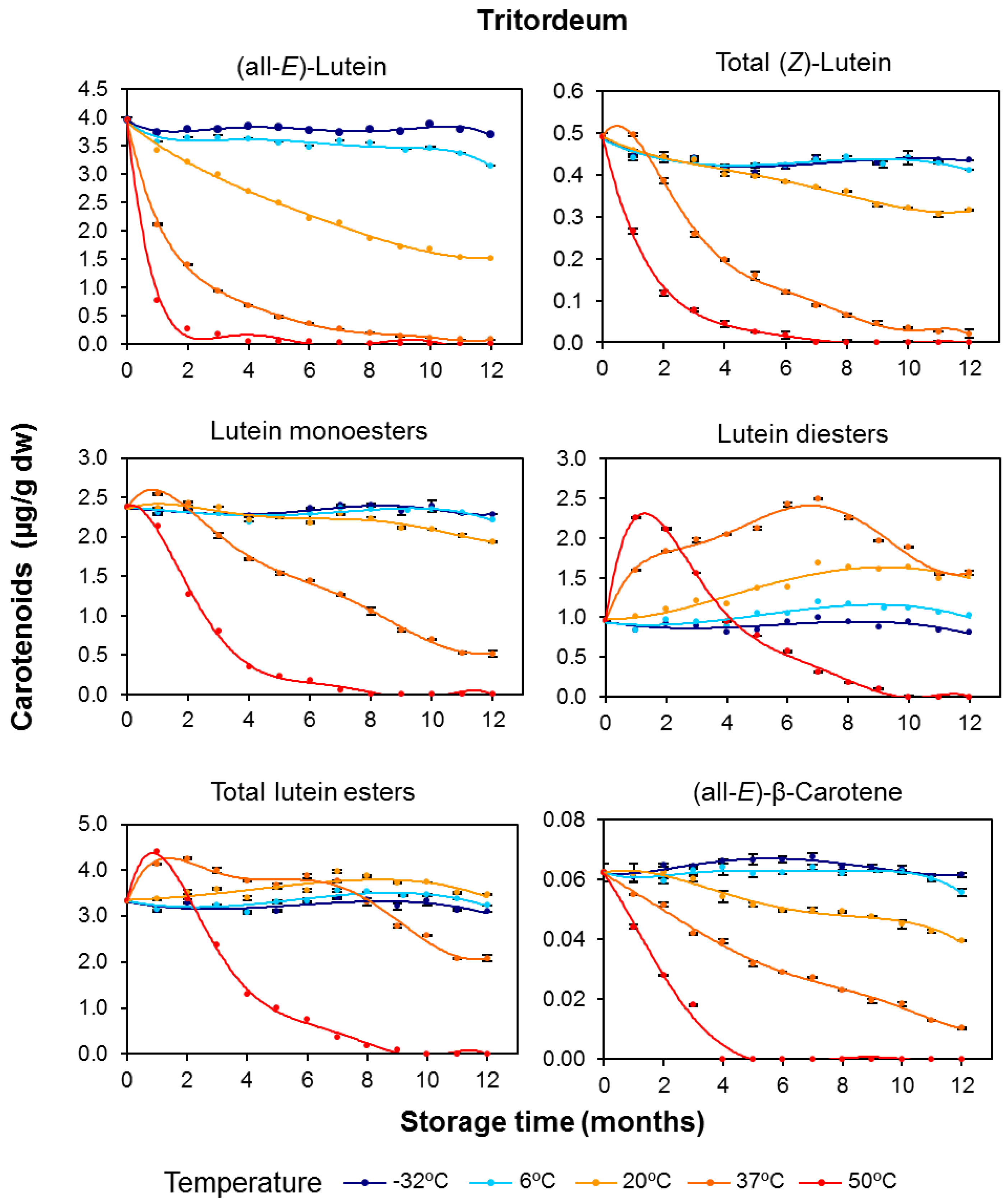
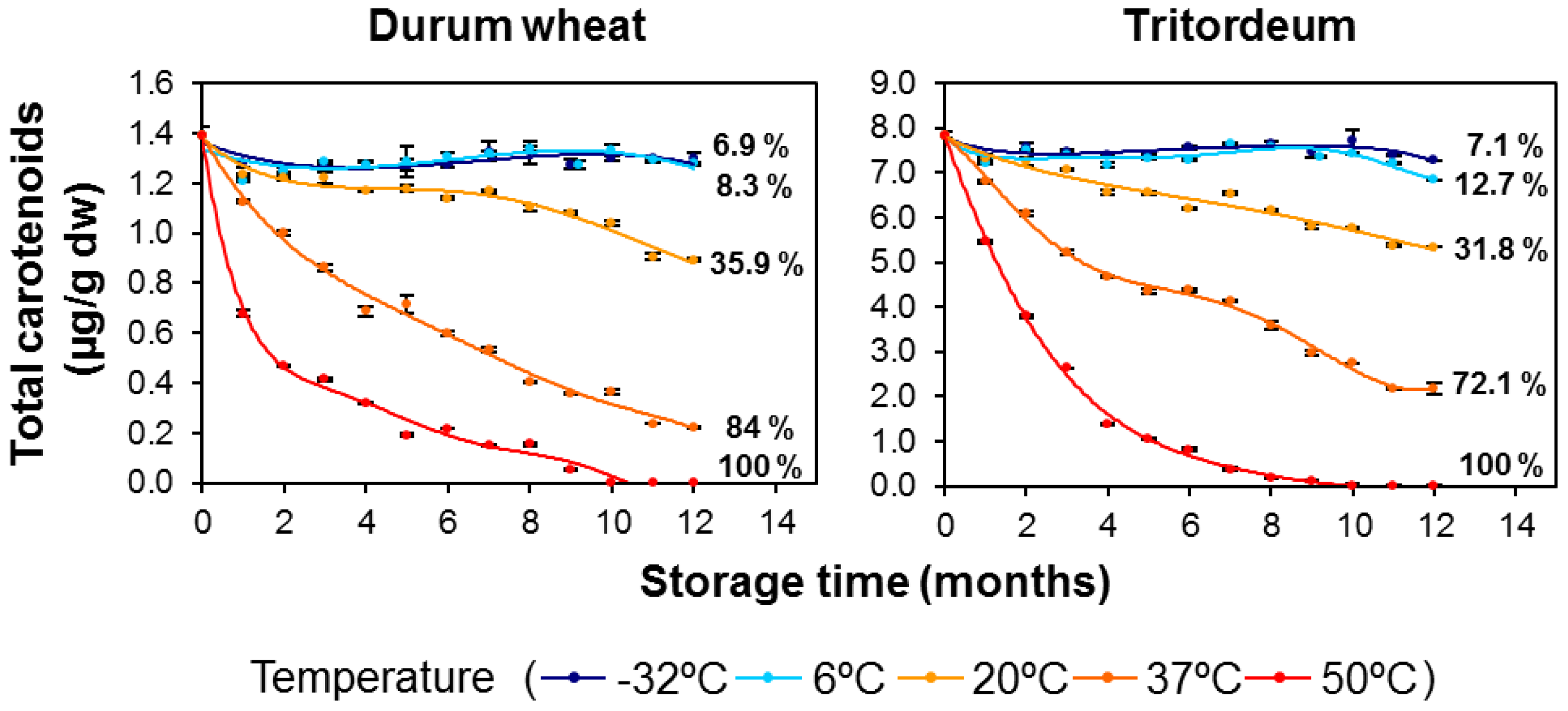
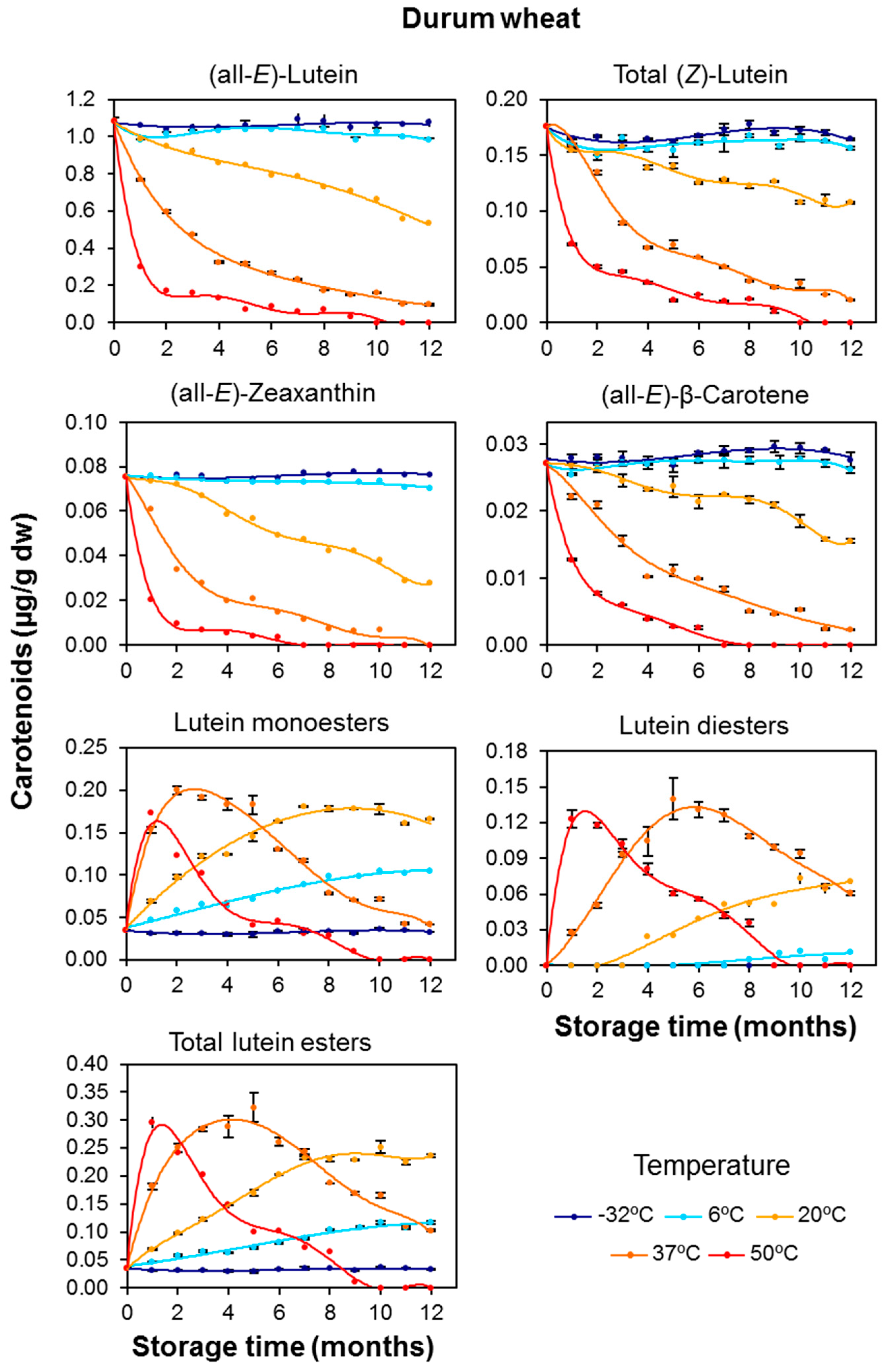
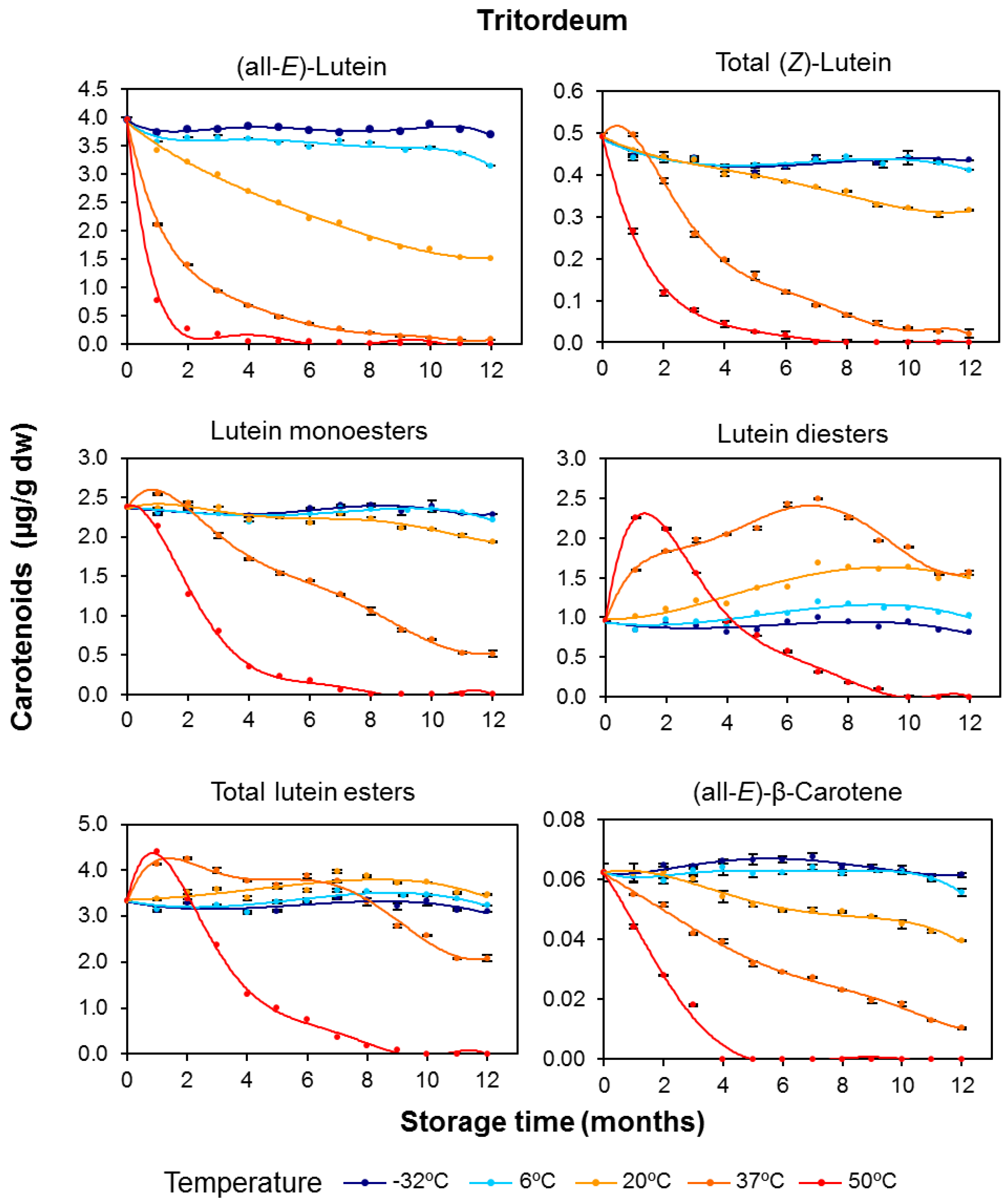
3.1. Carotenoid Content in Whole-Grain Flours: Effect of Long-Term Storage
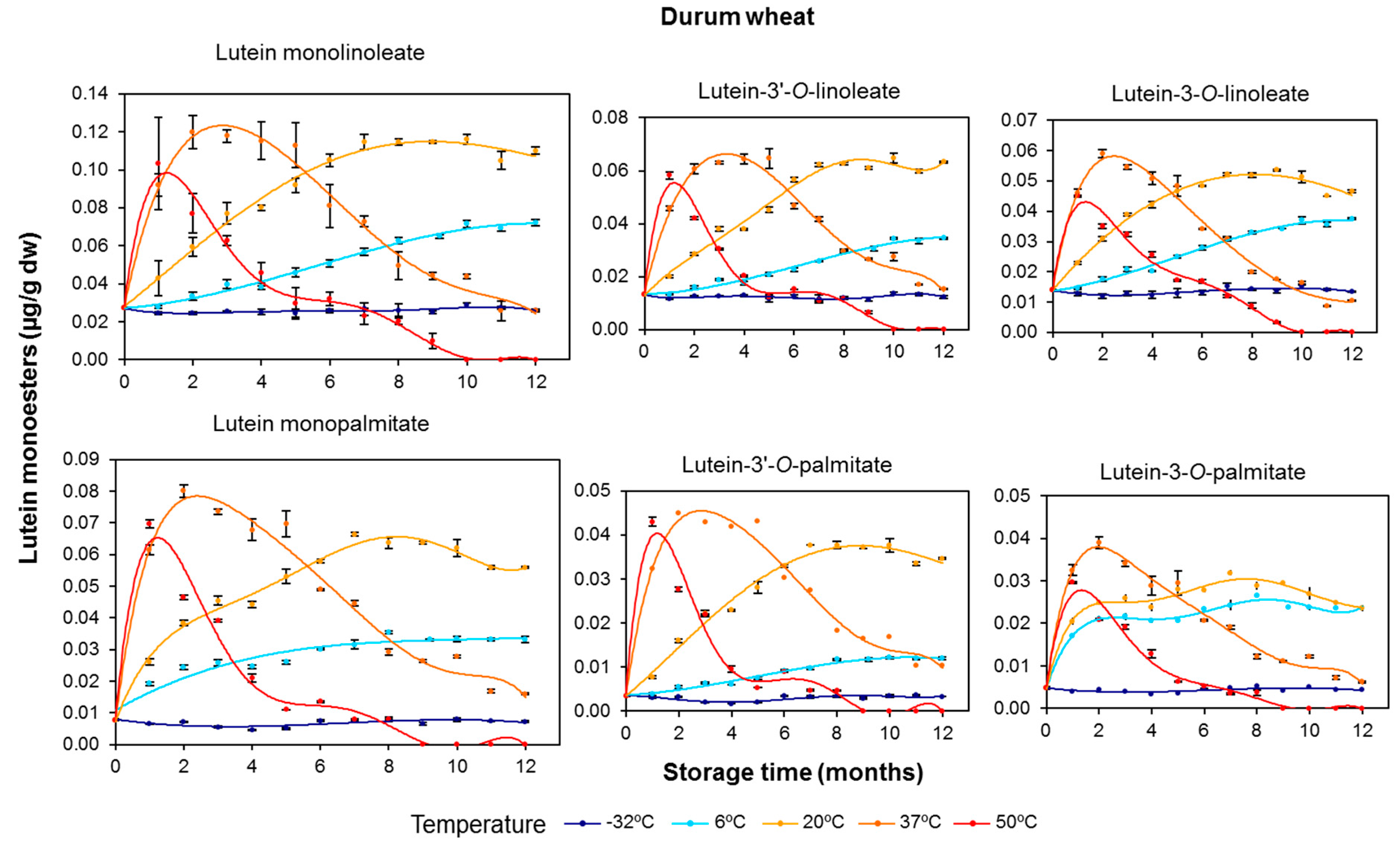
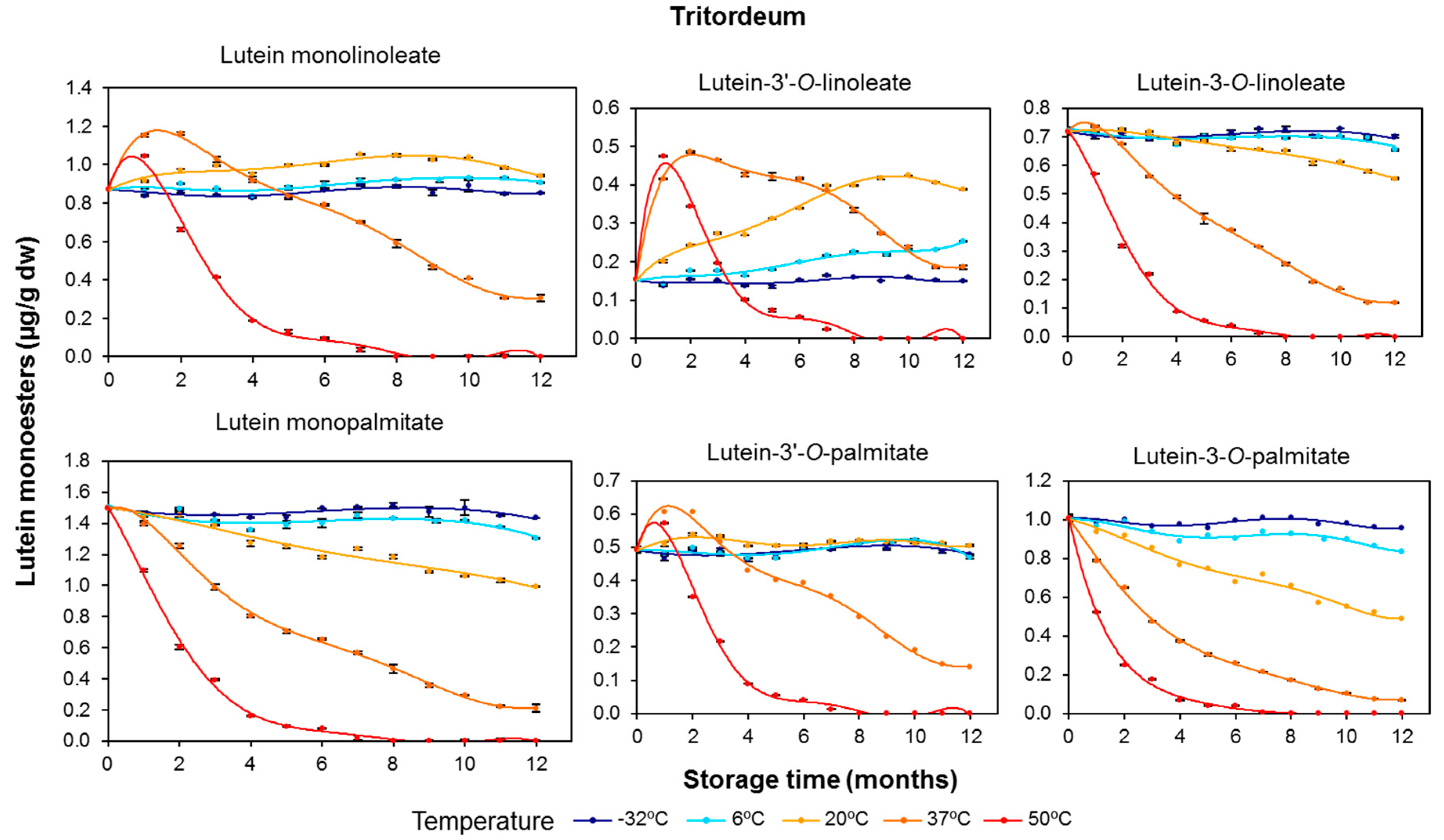
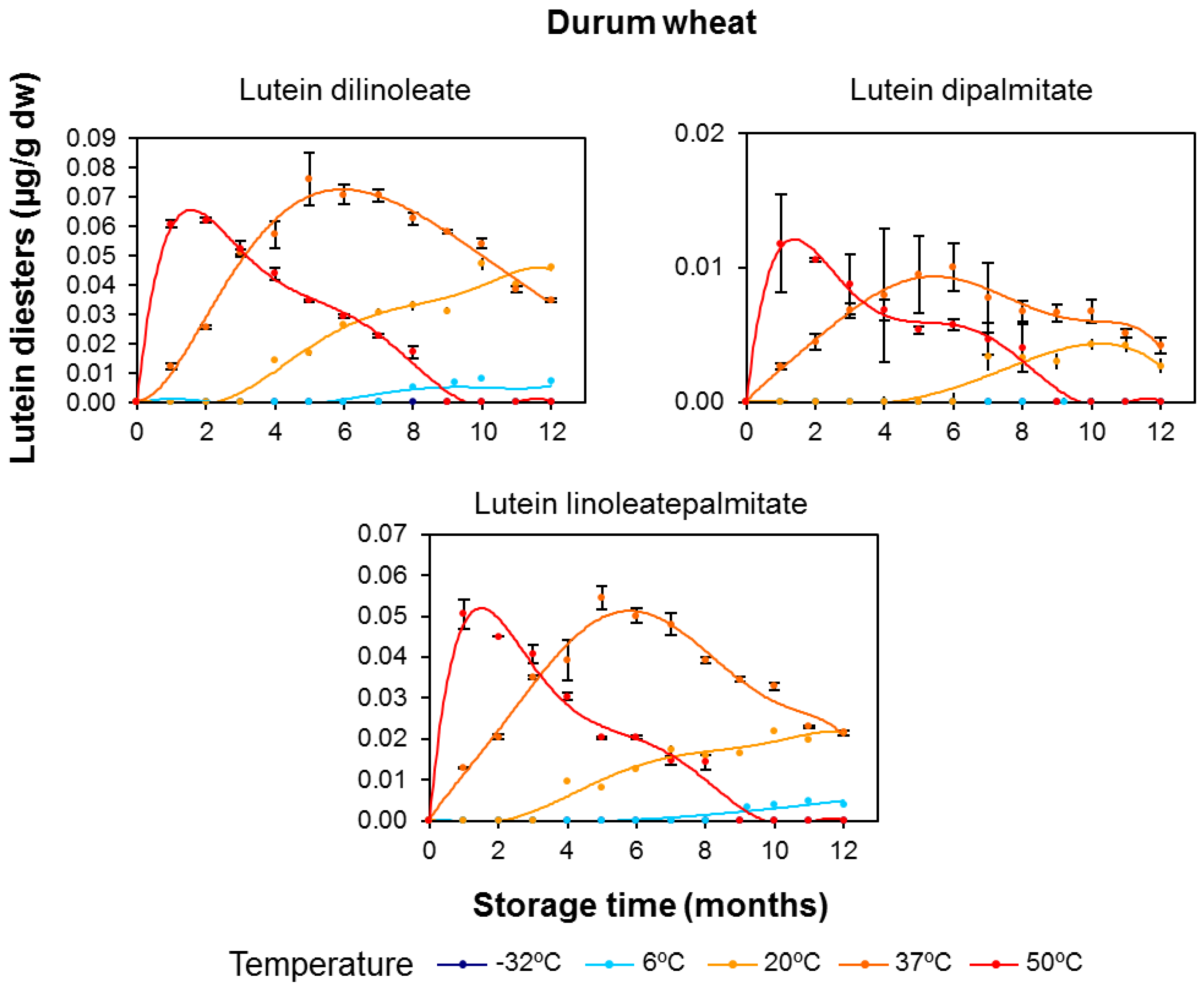
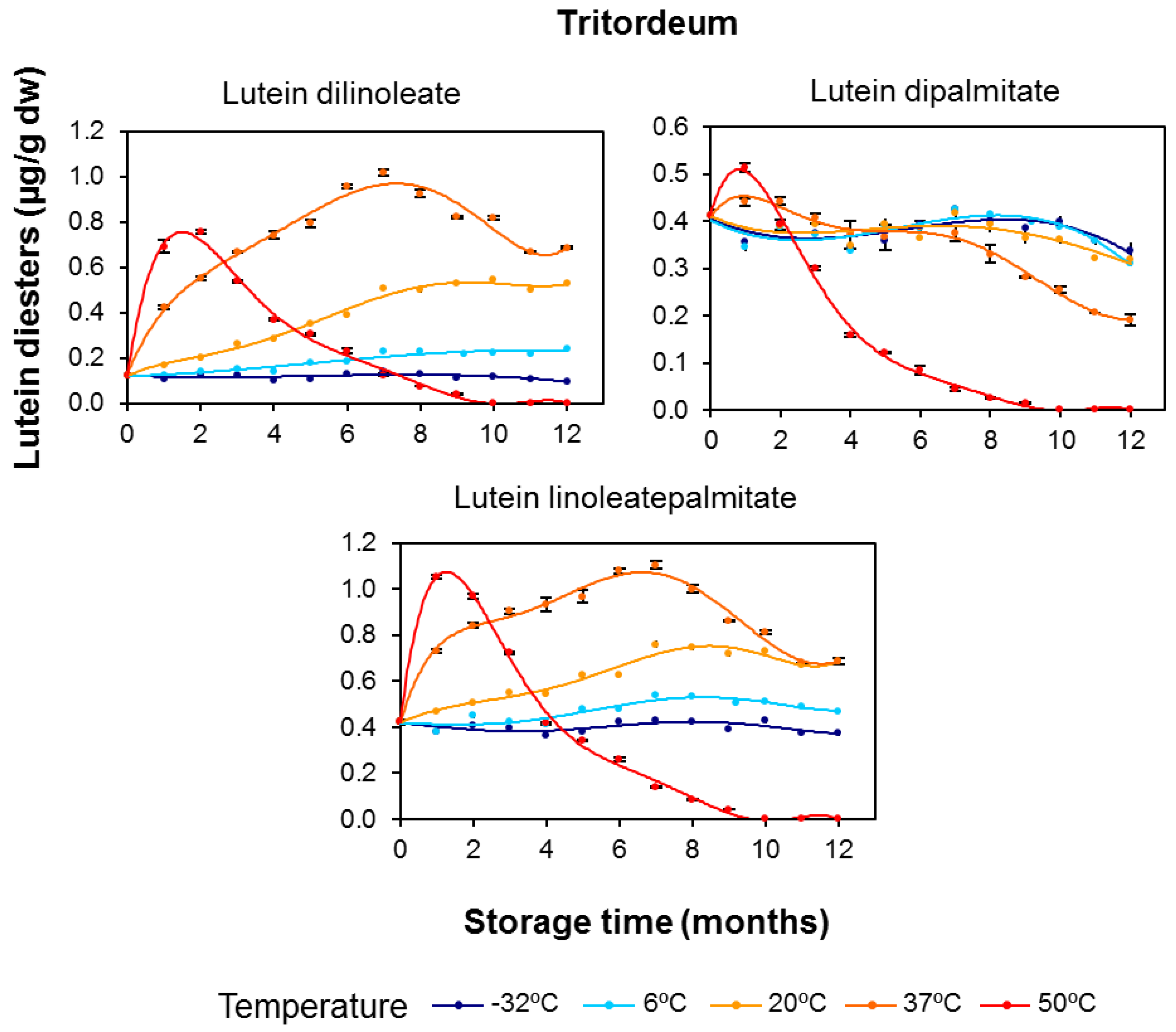
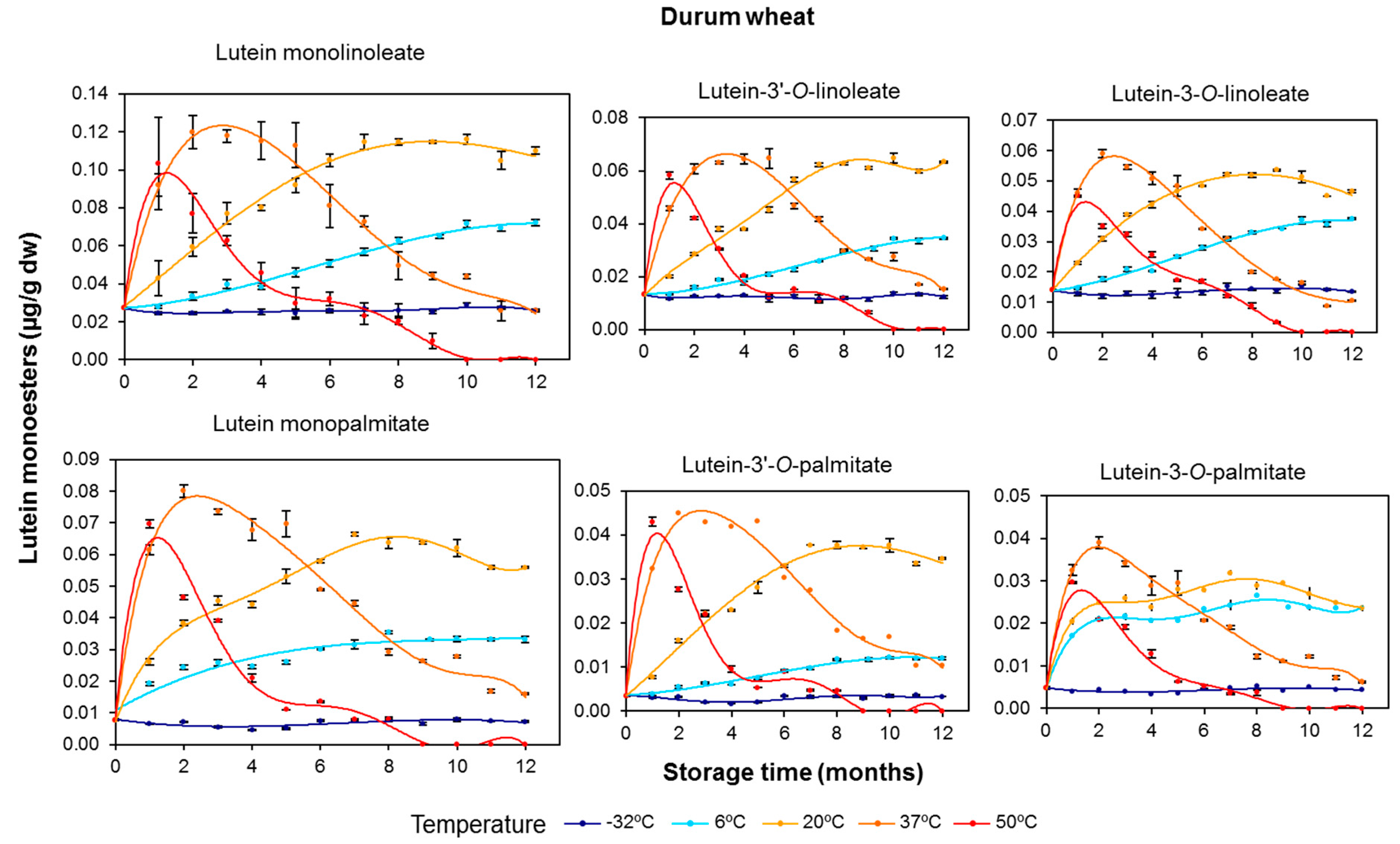
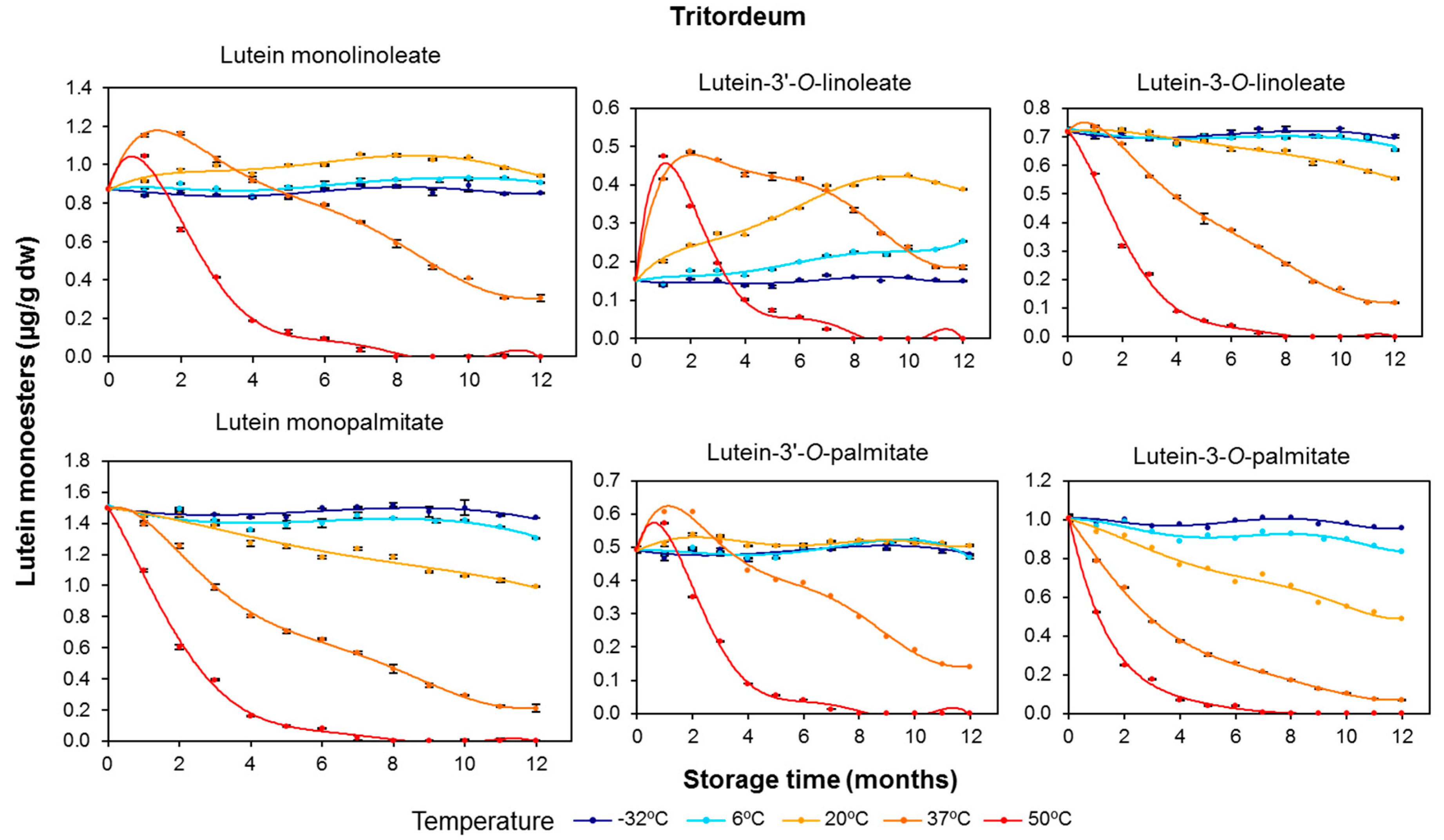
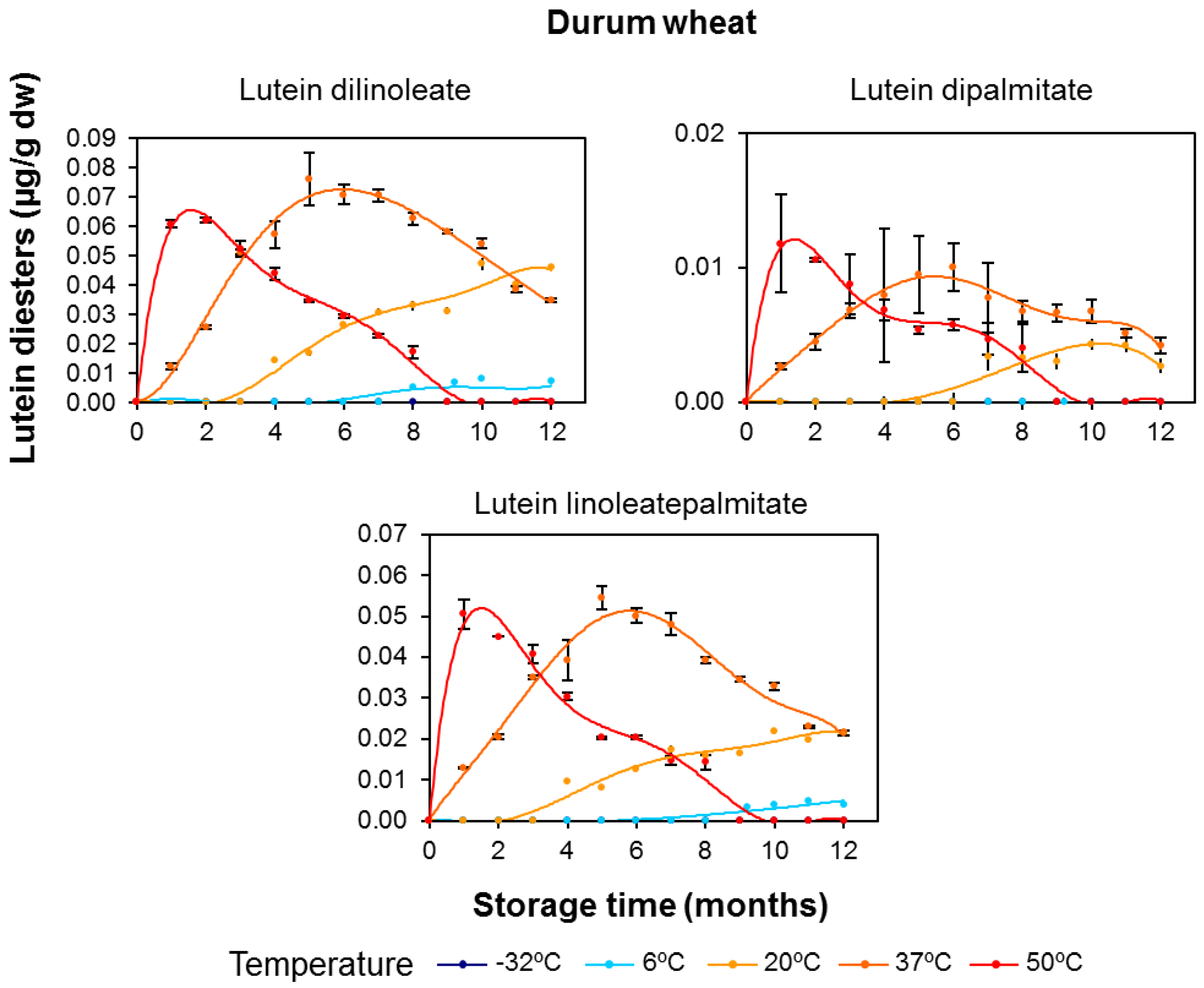
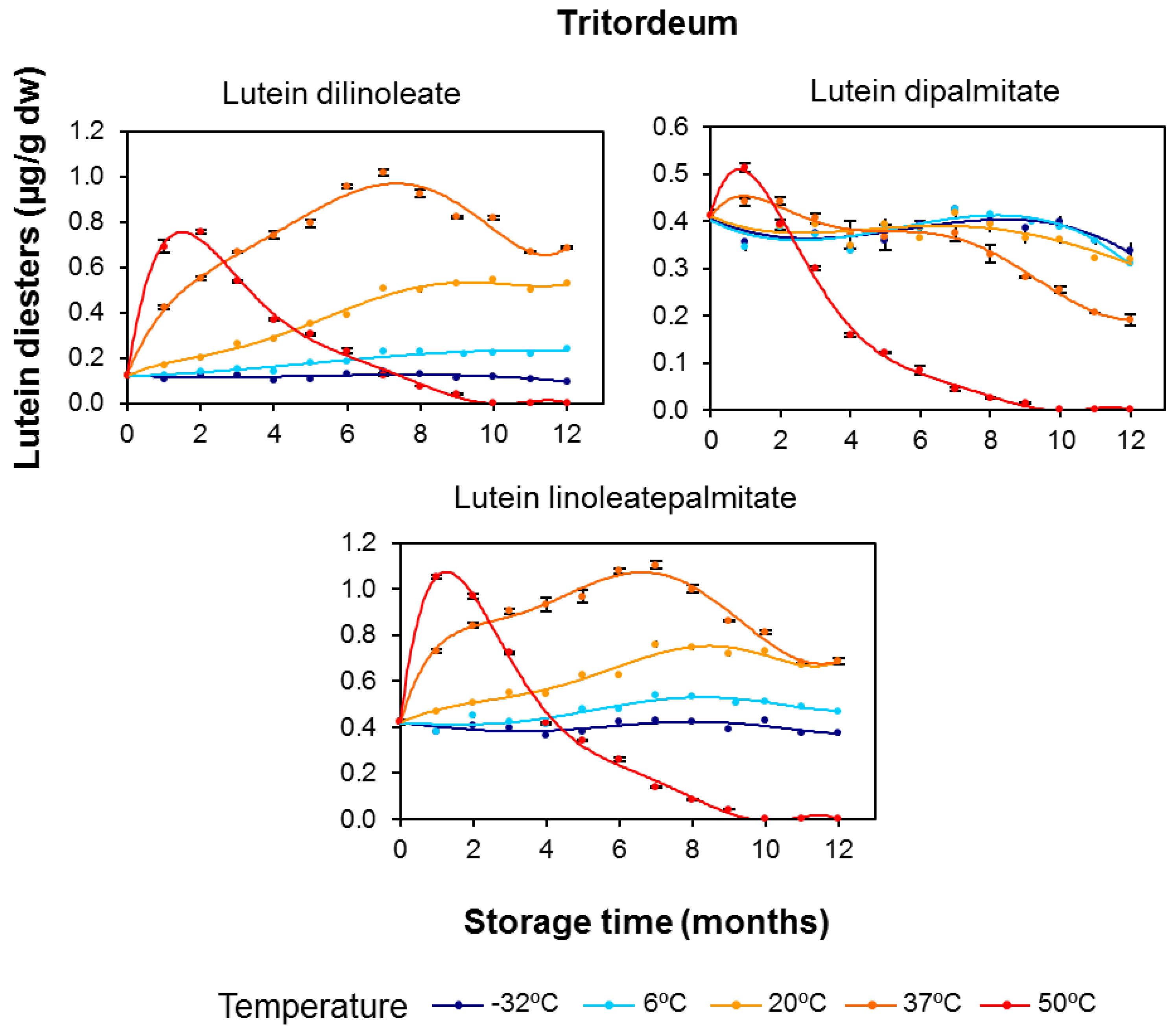
3.2. Effect of Long-Term Storage on the Esterified Lutein Fractions
3.3. Kinetics of Retention of Carotenoids during the Long-Term Storage of Wheat Flours
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. Carotenoids. Volume 1A: Isolation and Analysis; Birkhäuser Verlag: Basel, Switzerland, 1995. [Google Scholar]
- Howitt, C.A.; Pogson, B.J. Carotenoid accumulation and function in seeds and non-green tissues. Plant Cell Environ. 2006, 29, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Mayne, S.T. β-Carotene, carotenoids and disease prevention in humans. FASEB J. 1996, 10, 690–701. [Google Scholar] [PubMed]
- Cooper, D.A. Carotenoids in health and disease: Recent scientific evaluations, research recommendations and the consumer. J. Nutr. 2004, 134, 221S–224S. [Google Scholar] [PubMed]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. Carotenoids Volume 5: Nutrition and Health; Birkhäuser Verlag: Basel, Switzerland, 2009. [Google Scholar]
- Landrum, J.T.; Bone, R.A. Lutein, zeaxanthin, and the macular pigment. Arch. Biochem. Biophys. 2001, 385, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, V.; Kranl, K.; Hollmann, J.; Lindhauer, M.G.; Böhm, V.; Bitsch, R. Spectrophotometric determination of yellow pigment content and evaluation of carotenoids by high-performance liquid chromatography in durum wheat grain. J. Agric. Food Chem. 2002, 50, 6663–6668. [Google Scholar] [CrossRef] [PubMed]
- Ficco, D.B.M.; Mastrangelo, A.M.; Trono, D.; Borrelli, G.M.; De Vita, P.; Fares, C.; Beleggia, R.; Platani, C.; Papa, R. The colours of durum wheat: A review. Crop Pasture Sci. 2014, 65, 1–15. [Google Scholar] [CrossRef]
- Bai, C.; Twyman, R.M.; Farré, G.; Sanahuja, G.; Christou, P.; Capell, T.; Zhu, C.A. Golden era—Provitamin A enhancement in diverse crops. In Vitro Cell. Dev. Biol. Plant 2011, 47, 205–221. [Google Scholar] [CrossRef]
- Martín, A.; Sanchez-Monge, E.L. Citology and morphology of the amphiploid Hordeum chilense-Triticum turgidum conv. Durum. Euphytica 1982, 31, 261–267. [Google Scholar] [CrossRef]
- Atienza, S.G.; Ballesteros, J.; Martín, A.; Hornero-Méndez, D. Genetic variability of carotenoid concentration and degree of esterification among tritordeum (×Tritordeum Ascherson et Graebner) and durum wheat accessions. J. Agric. Food Chem. 2007, 55, 4244–4251. [Google Scholar] [CrossRef] [PubMed]
- Mellado-Ortega, E.; Hornero-Méndez, D. Isolation and identification of lutein esters, including their regioisomers, in tritordeum (×Tritordeum Ascherson et Graebner) grains. Evidences for a preferential xanthophyll acyltransferase activity. Food Chem. 2012, 135, 1344–1352. [Google Scholar] [CrossRef] [PubMed]
- Mellado-Ortega, E.; Hornero-Méndez, D. Carotenoid profiling of Hordeum chilense grains: The parental proof for the origin of the high carotenoid content and esterification pattern of tritordeum. J. Cereal Sci. 2015, 62, 15–21. [Google Scholar] [CrossRef]
- Chavez, A.L.; Sanchez, T.; Ceballos, H.; Rodriguez-Amaya, D.B.; Nestel, P.; Tohme, J.; Ishitani, M. Retention of carotenoids in cassava roots submitted to different processing methods. J. Sci. Food Agric. 2007, 87, 388–393. [Google Scholar] [CrossRef]
- Mugode, L.; Ha, B.; Kaunda, A.; Sikombe, T.; Phiri, S.; Mutale, R.; Davis, C.; Tanumihardjo, S.; De Moura, F. Carotenoid retention of biofortified provitamin A maize (Zea mays L.) after Zambian traditional methods of milling, cooking and storage. J. Agric. Food Chem. 2014, 62, 6317–6325. [Google Scholar] [CrossRef] [PubMed]
- Fardet, A. New hypotheses for the health-protective mechanisms of wholegrain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef] [PubMed]
- Britton, G.; Khachik, F. Carotenoids in food. In Carotenoids Volume 5: Nutrition and Health; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser Verlag: Basel, Switzerland, 2009; pp. 45–66. [Google Scholar]
- De Moura, F.F.; Miloff, A.; Boy, E. Retention of provitamin A carotenoids in staple crops targeted or biofortification in Africa: Cassava, maize, and sweet potato. Crit. Rev. Food Sci. Nutr. 2015, 55, 1246–1269. [Google Scholar] [CrossRef] [PubMed]
- Hemery, Y.; Rouau, X.; Lullien-Pellerin, V.; Barron, C.; Abecassis, J. Dry processes to develop wheat fractions and products with enhanced nutritional quality. J. Cereal Sci. 2007, 46, 327–347. [Google Scholar] [CrossRef]
- Li, L.; Yong, Y.; Qiang, X.; Owsiany, K.; Welsch, R.; Chitchumroonchokchai, C.; Lu, S.; Van Eck, J.; Deng, X.; Failla, M.; et al. The Or gene enhances carotenoid accumulation and stability during post-harvest storage of potato tubers. Mol. Plant 2012, 5, 339–352. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, D.; Rocheford, T.; Ferruzzi, M.G. Influence of temperature and humidity on the stability of carotenoids in biofortified maize (Zea mays L.) genotypes during controlled postharvest storage. J. Agric. Food Chem. 2016, 64, 2727–2736. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.T.; Asenstorfer, R.E.; Soriano, I.R.; Mares, D.J. Effect of temperature on lutein esterification and lutein stability in wheat grain. J. Cereal Sci. 2013, 58, 408–413. [Google Scholar] [CrossRef]
- Mellado-Ortega, E.; Hornero-Méndez, D. Carotenoid evolution during short-storage period of durum wheat (Triticum turgidum conv. durum) and tritordeum (×Tritordeum Ascherson et Graebner) whole-grain flours. Food Chem. 2016, 192, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Mellado-Ortega, E.; Hornero-Méndez, D. Effect of long-term storage on free and esterified carotenoids in durum wheat (Triticum turgidum conv. durum) and tritordeum (×Tritordeum Ascherson et Graebner) grains. Food Res. Int. 2017, 99, 877–890. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, J.B.; Ramírez, M.C.; Martínez, C.; Atienza, S.G.; Martín, A. Registration of HT621, a high carotenoid content tritordeum germplasm line. Crop Sci. 2005, 45, 2662–2663. [Google Scholar] [CrossRef]
- Mellado-Ortega, E.; Atienza, S.G.; Hornero-Méndez, D. Carotenoid evolution during postharvest storage of durum wheat (Triticum turgidum conv. durum) and tritordeum (×Tritordeum Ascherson et Graebner) grains. J. Cereal Sci. 2015, 62, 134–142. [Google Scholar] [CrossRef]
- Mínguez-Mosquera, M.I.; Hornero-Méndez, D. Separation and quantification of the carotenoid pigments in red peppers (Capsicum annuum L.), paprika and oleoresin by reversed-phase HPLC. J. Agric. Food Chem. 1993, 43, 1613–1620. [Google Scholar] [CrossRef]
- Britton, G. UV/visible spectroscopy. In Carotenoids. Volume 1B: Spectroscopy; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser Verlag: Basel, Switzerland, 1995; pp. 13–62. [Google Scholar]
- Upadhyay, S.K. Elementary. In Chemical Kinetics and Reaction Dynamics; Anamaya Publishers: New Delhi, India, 1996; pp. 1–45. [Google Scholar]
- Gayen, D.; Ali, N.; Sarkar, S.N.; Datta, S.K.; Datta, K. Down-regulation of lipoxygenase gene reduces degradation of carotenoids of golden rice during storage. Planta 2015, 242, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Rani, K.U.; Prasada Rao, U.J.S.; Leelavathi, K.; Haridas Rao, P. Distribution of enzymes in wheat flour mill streams. J. Cereal Sci. 2001, 34, 233–242. [Google Scholar] [CrossRef]
- Li, D.; Song, J.; Liu, C. Kinetic stability of lutein in freeze-dried sweet corn powder stored under different conditions. Food Sci. Technol. Res. 2014, 20, 65–70. [Google Scholar] [CrossRef]
- Choe, E.; Lee, J.; Park, K.; Lee, S. Effects of heat pretreatment on lipid and pigments of freeze-dried spinach. J. Food Sci. 2001, 66, 1074–1079. [Google Scholar] [CrossRef]
- Urquhart, A.A.; Altosaar, I.; Matlashewski, G.J.; Sahasrabudhe, M.R. Localization of lipase activity in oat grains and milled oat fractions. Cereal Chem. 1983, 60, 181–183. [Google Scholar]
- Barros, M.; Fleuri, L.F.; Macedo, G.A. Seed lipases: Sources, applications and properties—A review. Braz. J. Chem. Eng. 2010, 27, 15–29. [Google Scholar] [CrossRef]
- Farrington, F.F.; Warwick, M.J.; Shearer, G. Changes in the carotenoids and sterol fractions during the prolonged storage of wheat flour. J. Sci. Food Agric. 1981, 32, 948–950. [Google Scholar] [CrossRef]
- Ahmad, F.T.; Mather, D.E.; Law, H.; Li, M.; Yousif, S.; Chalmers, K.J.; Asenstorfer, R.E.; Mares, D.J. Genetic control of lutein esterification in wheat (Triticum aestivum L.) grain. J. Cereal Sci. 2015, 64, 109–115. [Google Scholar] [CrossRef]
- Mattera, G.; Cabrera, A.; Hornero-Méndez, D.; Atienza, S.G. Lutein esterification in wheat endosperm is controlled by the homoeologous group 7, and is increased by the simultaneous presence of chromosomes 7D and 7Hch from Hordeum chilense. Crop Pasture Sci. 2015, 66, 912–921. [Google Scholar] [CrossRef]
- Mattera, G.; Hornero-Méndez, D.; Atienza, S.G. Lutein ester profile in wheat and tritordeum can be modulated by temperature: Evidences for regioselectivity and fatty acid preferential of enzymes encoded by genes on chromosomes 7D and 7Hch. Food Chem. 2017, 219, 199–206. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.; Perry, H.J.; Harwood, J.L. A comparison of lipase activity in various cereal grains. J. Cereal Sci. 1992, 16, 153–163. [Google Scholar] [CrossRef]
- Hidalgo, A.; Brandolini, A. Kinetics of carotenoids degradation during the storage of einkorn (Triticum monococcum L. ssp. monococcum) and bread wheat (Triticum aestivum L. ssp. aestivum) flours. J. Agric. Food Chem. 2008, 56, 11300–11305. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, G.M.; Troccoli, A.; Di Fonzo, N.; Fares, C. Durum wheat lipoxygenase activity and other quality parameters that affect pasta colour. Cereal Chem. 1999, 76, 335–340. [Google Scholar] [CrossRef]
- Leenhardt, F.; Lyan, B.; Rock, E.; Boussard, A.; Potus, J.; Chanliaud, E.; Remesy, C. Wheat lipoxygenase activity induces greater loss of carotenoids than vitamin E during breadmaking. J. Agric. Food Chem. 2006, 54, 1710–1715. [Google Scholar] [CrossRef] [PubMed]
- Fratianni, A.; Di Criscio, T.; Mignogna, R.; Panfili, G. Carotenoids, tocols and retinols evolution during egg pasta-making processes. Food Chem. 2012, 131, 590–595. [Google Scholar] [CrossRef]
- Dhuique-Mayer, C.; Tbatou, M.; Carail, M.; Caris-Veyrat, C.; Dornier, M.; Amiot, M.J. Thermal degradation of antioxidant micronutrients in citrus juice: Kinetics and newly formed compounds. J. Agric. Food Chem. 2007, 55, 4209–4216. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reaction Order | Reaction Rate Expression | Integrated Expression | Graphical Representation | Half-Life a (t1/2) | D b (t1/10) |
|---|---|---|---|---|---|
| Zero n = 0 | −dC/dt = kC0 = k | C-C0 = −kt | C-C0 vs. t Slope = −k | t1/2 = C0/2k | t1/10 = 0.9C0/k |
| First n = 1 | −dC/dt = kC1 = kC | Ln(C/C0) = −kt | Ln(C/C0) vs. t Slope = −k | t1/2 = Ln(2)/k | t1/10 = Ln(10)/k |
| HPLC Peak a | Pigment | Concentration (μg/g Dry Weight) b | |
|---|---|---|---|
| Durum Wheat (Don Pedro Variety) | Tritordeum (HT621 Advanced Line) | ||
| 1 | (all-E)-Zeaxanthin | 0.08 ± 0.00 | - |
| 2 | (all-E)-Lutein | 1.08 ± 0.02 | 3.95 ± 0.04 |
| 3 | (9Z)-Lutein | 0.06 ± 0.00 | 0.19 ± 0.01 |
| 4 | (13Z)-Lutein | 0.12 ± 0.00 | 0.30 ± 0.01 |
| 5 | Lutein-3′-O-linoleate | 0.01 ± 0.00 | 0.15 ± 0.00 |
| 6 | Lutein-3-O-linoleate | 0.01 ± 0.00 | 0.72 ± 0.01 |
| 5 + 6 | Lutein monolinoleate | 0.03 ± 0.00 | 0.87 ± 0.00 |
| 7 | Lutein-3′-O-palmitate | 0.00 ± 0.00 | 0.49 ± 0.01 |
| 8 | Lutein-3-O-palmitate | 0.00 ± 0.00 | 1.01 ± 0.02 |
| 7 + 8 | Lutein monopalmitate | 0.01 ± 0.00 | 1.50 ± 0.01 |
| 9 | (all-E)-β-Carotene | 0.02 ± 0.00 | 0.06 ± 0.00 |
| 10 | Lutein-3,3′-dilinoleate | n.d. c | 0.12 ± 0.00 |
| 11 | Lutein-3′-O-linoleate-3-O-palmitate plus Lutein-3′-O-palmitate-3-O-linoleate | n.d. | 0.42 ± 0.01 |
| 12 | Lutein-3,3′-dipalmitate | n.d. | 0.41 ± 0.01 |
| Lutein monoesters | 0.04 ± 0.00 | 2.37 ± 0.01 | |
| Lutein diesters | - | 0.96 ± 0.01 | |
| Total lutein esters | 0.04 ± 0.00 | 3.33 ± 0.02 | |
| Total free lutein | 1.26 ± 0.02 | 4.44 ± 0.09 | |
| Total lutein | 1.29 ± 0.02 | 7.77 ± 0.07 | |
| Total carotenoids | 1.39 ± 0.03 | 7.83 ± 0.07 | |
| Regioisomers ratios | |||
| Lutein-3-O-linoleate/Lutein-3′-O-linoleate | 1 | 5 | |
| Lutein-3-O-palmitate/Lutein-3′-O-palmitate | 1 | 2 | |
| Pigment | T (°C) | Durum Wheat (Don Pedro Variety) | Tritordeum (HT621 Advanced Line) | ||
|---|---|---|---|---|---|
| k (×10−3 Month−1) | R2 | k (×10−3 Month−1) | R2 | ||
| (all-E)-Zeaxanthin | −32 | 0.2 | 0.33 | - | - |
| 6 | 0.4 | 0.80 | - | - | |
| 20 | 4 | 0.98 | - | - | |
| 37 | 5 | 0.80 | - | - | |
| 50 | 3 | 0.43 | - | - | |
| (all-E)-Lutein | −32 | 1 | 0.06 | 7 | 0.15 |
| 6 | 3 | 0.18 | 41 | 0.72 | |
| 20 | 42 | 0.98 | 198 | 0.96 | |
| 37 | 68 | 0.80 | 232 | 0.65 | |
| 50 | 51 | 0.47 | 160 | 0.33 | |
| Total free (Z)-Lutein | −32 | 0.3 | 0.04 | 2 | 0.14 |
| 6 | 0.2 | 0.01 | 3 | 0.31 | |
| 20 | 5 | 0.92 | 15 | 0.97 | |
| 37 | 12 | 0.86 | 42 | 0.87 | |
| 50 | 10 | 0.63 | 28 | 0.57 | |
| (all-E)-β-Carotene | −32 | 0.1 | 0.35 | 0.1 | 0.02 |
| 6 | 0.5 | 0.06 | 0.2 | 0.12 | |
| 20 | 8 | 0.93 | 2 | 0.95 | |
| 37 | 2 | 0.90 | 4 | 0.97 | |
| 50 | 1 | 0.62 | 4 | 0.62 | |
| Total free lutein | −32 | 1 | 0.06 | 9 | 0.19 |
| 6 | 3 | 0.15 | 42 | 0.70 | |
| 20 | 47 | 0.98 | 213 | 0.95 | |
| 37 | 80 | 0.82 | 273 | 0.70 | |
| 50 | 60 | 0.50 | 188 | 0.36 | |
| Total free carotenoids | −32 | 1 | 0.01 | 9 | 0.19 |
| 6 | 6 | 0.20 | 44 | 0.70 | |
| 20 | 50 | 0.96 | 215 | 0.96 | |
| 37 | 85 | 0.80 | 277 | 0.70 | |
| 50 | 64 | 0.48 | 192 | 0.36 | |
| Total lutein | −32 | 2 | 0.08 | 10 | 0.07 |
| 6 | 4 | 0.15 | 29 | 0.21 | |
| 20 | 23 | 0.88 | 191 | 0.95 | |
| 37 | 85 | 0.95 | 432 | 0.95 | |
| 50 | 80 | 0.73 | 546 | 0.75 | |
| Total carotenoids | −32 | 0.8 | 0.01 | 10 | 0.07 |
| 6 | 1 | 0.01 | 29 | 0.21 | |
| 20 | 33 | 0.88 | 192 | 0.95 | |
| 37 | 90 | 0.94 | 436 | 0.95 | |
| 50 | 83 | 0.71 | 550 | 0.75 | |
| Pigment | T (°C) | Durum Wheat (Don Pedro Variety) | Tritordeum (HT621 Advanced Line) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| k (×10−3 Month−1) | R2 | t1/2 (Months) | t1/10 (D) (Months) | k (×10−3 Month−1) | R2 | t1/2 (Months) | t1/10 (D) (Months) | ||
| (all-E)-Zeaxanthin | −32 | 3 | 0.33 | 277 | 921 | - | - | - | - |
| 6 | 5 | 0.79 | 141 | 470 | - | - | - | - | |
| 20 | 85 | 0.96 | 8 | 27 | - | - | - | - | |
| 37 | 252 | 0.97 | 3 | 9 | - | - | - | - | |
| 50 | 473 | 0.87 | 1 | 5 | - | - | - | - | |
| (all-E)-Lutein | −32 | 1 | 0.06 | 693 | 2302 | 2 | 0.15 | 385 | 1279 |
| 6 | 3 | 0.18 | 217 | 719 | 12 | 0.72 | 60 | 198 | |
| 20 | 53 | 0.96 | 13 | 43 | 82 | 0.99 | 8 | 28 | |
| 37 | 191 | 0.97 | 4 | 12 | 329 | 0.99 | 2 | 7 | |
| 50 | 300 | 0.84 | 2 | 8 | 596 | 0.88 | 1 | 4 | |
| Total free (Z)-lutein | −32 | 2 | 0.04 | 385 | 1279 | 4 | 0.13 | 165 | 548 |
| 6 | 0.7 | 0.00 | 990 | 3289 | 6 | 0.31 | 110 | 365 | |
| 20 | 38 | 0.93 | 18 | 60 | 39 | 0.98 | 18 | 59 | |
| 37 | 177 | 0.98 | 4 | 13 | 283 | 0.99 | 2 | 8 | |
| 50 | 249 | 0.88 | 3 | 9 | 559 | 0.99 | 1 | 4 | |
| (all-E)-β-Carotene | −32 | 5 | 0.35 | 139 | 460 | 1 | 0.02 | 576 | 1918 |
| 6 | 2 | 0.06 | 462 | 1535 | 3 | 0.12 | 210 | 698 | |
| 20 | 45 | 0.90 | 16 | 52 | 38 | 0.95 | 19 | 62 | |
| 37 | 203 | 0.96 | 3 | 11 | 141 | 0.98 | 5 | 16 | |
| 50 | 387 | 0.95 | 2 | 6 | 417 | 0.99 | 2 | 6 | |
| Total free lutein | −32 | 1 | 0.07 | 693 | 2302 | 2 | 0.19 | 330 | 1096 |
| 6 | 3 | 0.15 | 248 | 822 | 11 | 0.71 | 63 | 209 | |
| 20 | 51 | 0.97 | 14 | 45 | 76 | 0.99 | 9 | 30 | |
| 37 | 189 | 0.98 | 4 | 12 | 321 | 0.99 | 2 | 7 | |
| 50 | 290 | 0.85 | 2 | 8 | 630 | 0.94 | 1 | 4 | |
| Total free carotenoids | −32 | 0.7 | 0.01 | 990 | 3289 | 2 | 0.19 | 347 | 1151 |
| 6 | 5 | 0.20 | 151 | 500 | 11 | 0.70 | 64 | 211 | |
| 20 | 53 | 0.96 | 13 | 44 | 75 | 0.99 | 9 | 31 | |
| 37 | 191 | 0.97 | 4 | 12 | 311 | 0.99 | 2 | 7 | |
| 50 | 296 | 0.85 | 2 | 8 | 636 | 0.94 | 1 | 4 | |
| Total lutein | −32 | 1 | 0.08 | 533 | 1771 | 1 | 0.07 | 495 | 1644 |
| 6 | 3 | 0.15 | 248 | 822 | 4 | 0.21 | 178 | 590 | |
| 20 | 27 | 0.86 | 25 | 84 | 30 | 0.96 | 23 | 77 | |
| 37 | 145 | 0.98 | 5 | 16 | 103 | 0.98 | 7 | 22 | |
| 50 | 285 | 0.93 | 2 | 8 | 465 | 0.98 | 1 | 5 | |
| Total carotenoids | −32 | 0.5 | 0.00 | 1386 | 4604 | 1 | 0.07 | 495 | 1644 |
| 6 | 0.9 | 0.01 | 770 | 2558 | 4 | 0.21 | 178 | 590 | |
| 20 | 30 | 0.87 | 23 | 78 | 30 | 0.96 | 23 | 77 | |
| 37 | 148 | 0.98 | 5 | 16 | 103 | 0.98 | 7 | 22 | |
| 50 | 290 | 0.93 | 2 | 8 | 466 | 0.98 | 1 | 5 | |
| Pigment | T (°C) | Durum Wheat (Don Pedro Variety) | Tritordeum (HT621 Advanced Line) | ||
|---|---|---|---|---|---|
| k (×10−3 Month−1) | R2 | k (×10−3 Month−1) | R2 | ||
| Lutein monolinoleate | 20 | - | - | - | - |
| 37 | 210 | 0.96 | 113 | 0.87 | |
| 50 | 262 | 0.96 | 388 | 0.90 | |
| Lutein-3′-O-linoleate | 20 | - | - | - | - |
| 37 | 184 | 0.94 | 103 | 0.91 | |
| 50 | 230 | 0.90 | 487 | 0.98 | |
| Lutein-3-O-linoleate | 20 | - | - | 22 | 0.92 |
| 37 | 200 | 0.95 | 140 | 0.94 | |
| 50 | 284 | 0.91 | 526 | 0.96 | |
| Lutein monopalmitate | 20 | - | - | 34 | 0.97 |
| 37 | 204 | 0.95 | 158 | 0.98 | |
| 50 | 332 | 0.94 | 544 | 0.96 | |
| Lutein-3′-O-palmitate | 20 | - | - | - | - |
| 37 | 201 | 0.95 | 147 | 0.97 | |
| 50 | 329 | 0.87 | 606 | 0.98 | |
| Lutein-3-O-palmitate | 20 | - | - | 59 | 0.98 |
| 37 | 186 | 0.96 | 228 | 1 | |
| 50 | 336 | 0.96 | 632 | 0.96 | |
| Total monoesters | 20 | - | - | 13 | 0.78 |
| 37 | 208 | 0.95 | 114 | 0.91 | |
| 50 | 310 | 0.94 | 471 | 0.95 | |
| Lutein-3,3′-dilinoleate | 20 | - | - | - | - |
| 37 | 111 | 0.90 | 85 | 0.92 | |
| 50 | 188 | 0.97 | 370 | 0.97 | |
| Lutein-3′-O-linoleate-3-O-palmitate plus Lutein-3′-O-palmitate-3-O-linoleate | 20 | - | - | - | - |
| 37 | 139 | 0.95 | 103 | 0.96 | |
| 50 | 201 | 0.96 | 400 | 0.97 | |
| Lutein-3,3′-dipalmitate | 20 | - | - | - | - |
| 37 | 90 | 0.78 | 66 | 0.83 | |
| 50 | 156 | 0.96 | 451 | 0.98 | |
| Total diesters | 20 | - | - | - | - |
| 37 | 122 | 0.93 | 88 | 0.93 | |
| 50 | 190 | 0.98 | 395 | 0.97 | |
| Total esters | 20 | - | - | - | - |
| 37 | 162 | 0.96 | - | - | |
| 50 | 333 | 0.81 | 472 | 0.98 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mellado-Ortega, E.; Hornero-Méndez, D. Lutein Esterification in Wheat Flour Increases the Carotenoid Retention and Is Induced by Storage Temperatures. Foods 2017, 6, 111. https://doi.org/10.3390/foods6120111
Mellado-Ortega E, Hornero-Méndez D. Lutein Esterification in Wheat Flour Increases the Carotenoid Retention and Is Induced by Storage Temperatures. Foods. 2017; 6(12):111. https://doi.org/10.3390/foods6120111
Chicago/Turabian StyleMellado-Ortega, Elena, and Dámaso Hornero-Méndez. 2017. "Lutein Esterification in Wheat Flour Increases the Carotenoid Retention and Is Induced by Storage Temperatures" Foods 6, no. 12: 111. https://doi.org/10.3390/foods6120111
APA StyleMellado-Ortega, E., & Hornero-Méndez, D. (2017). Lutein Esterification in Wheat Flour Increases the Carotenoid Retention and Is Induced by Storage Temperatures. Foods, 6(12), 111. https://doi.org/10.3390/foods6120111