Glycomacropeptide Reduces Intestinal Epithelial Cell Barrier Dysfunction and Adhesion of Entero-Hemorrhagic and Entero-Pathogenic Escherichia coli in Vitro



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bacteria and Culture Conditions
2.3. Mammalian Cell Culture
2.4. Adhesion Assay
2.4.1. Standard Inhibition Assay
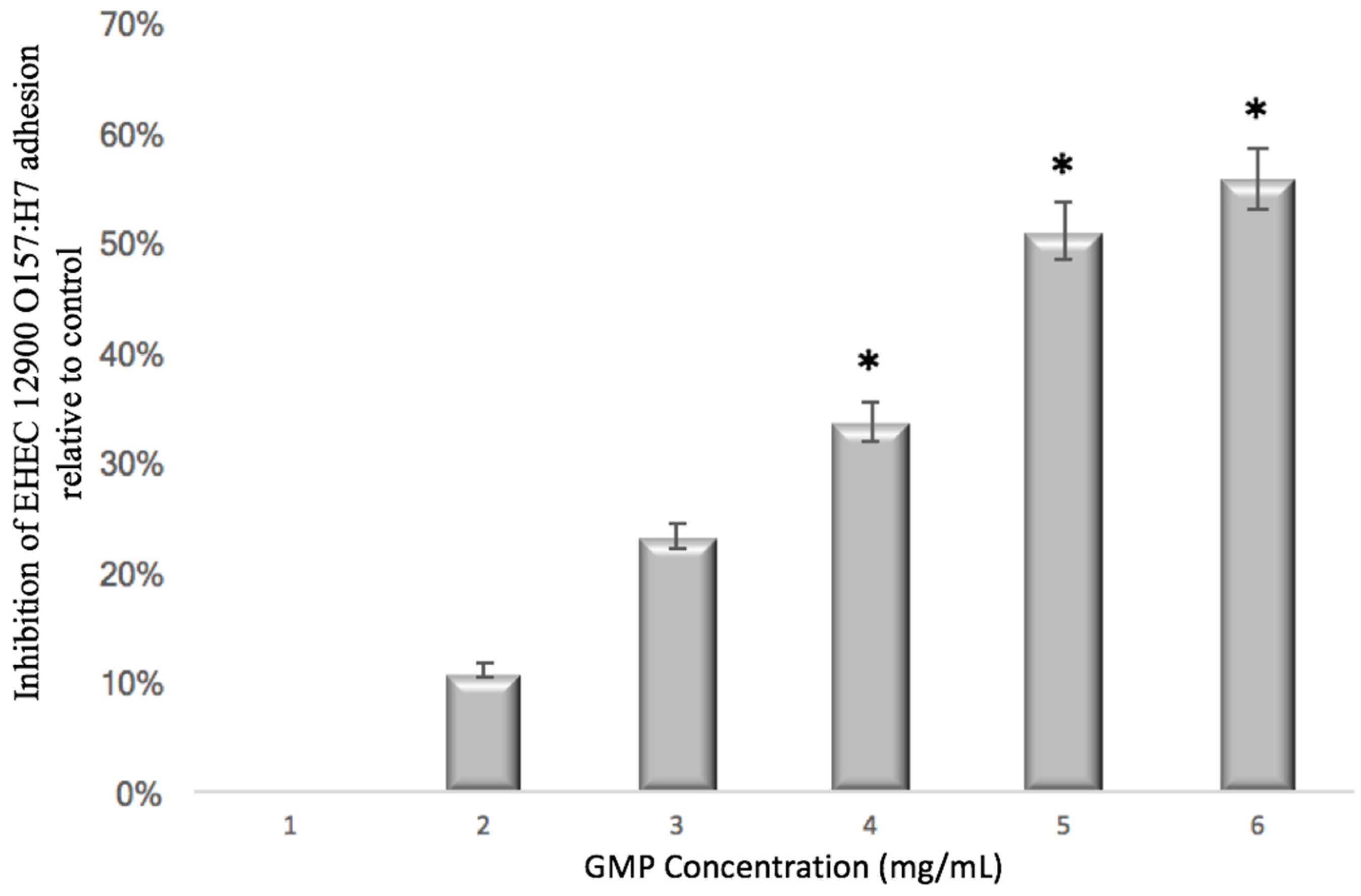
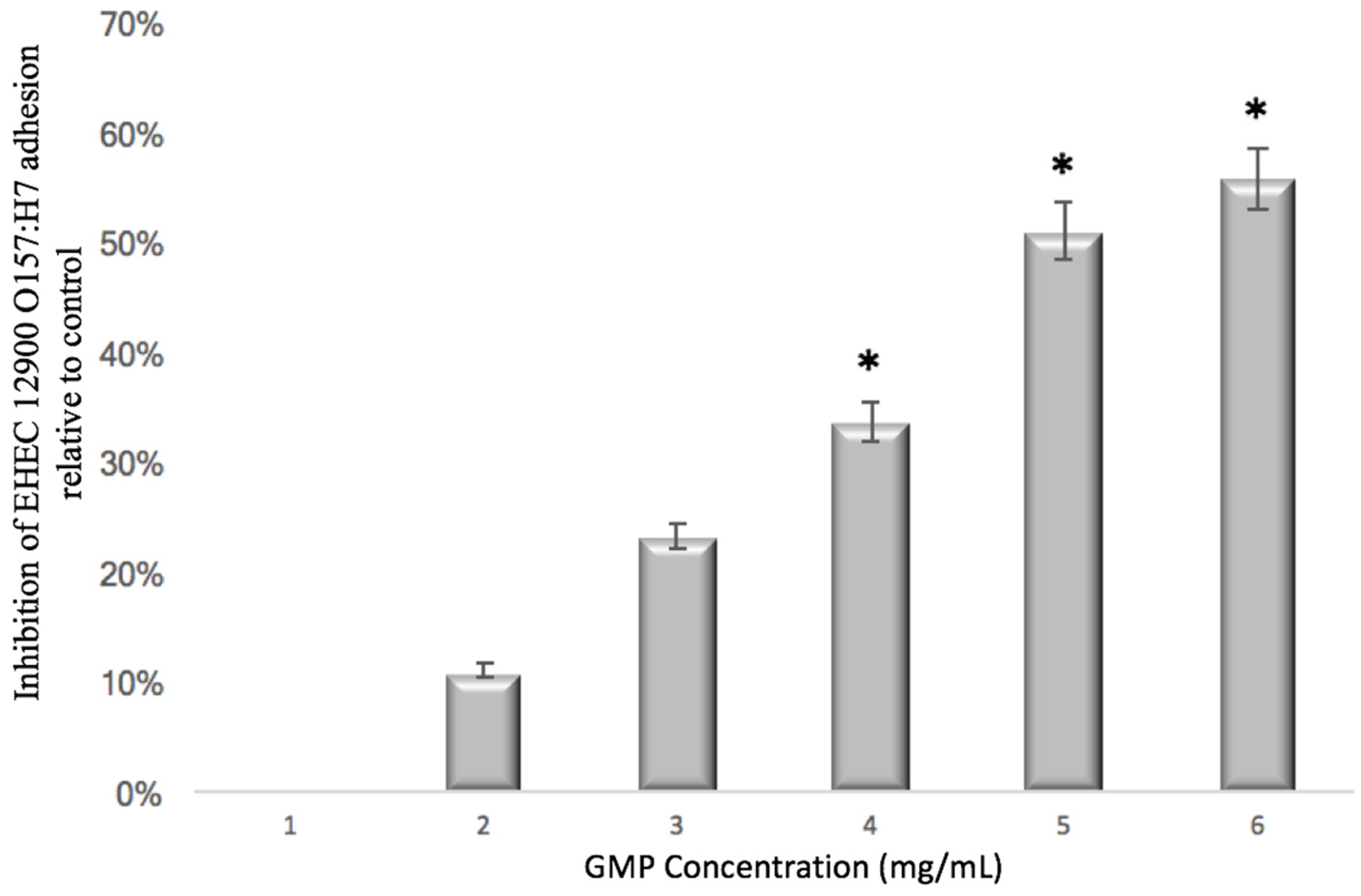
2.4.2. Concentration Dependency Assay
2.4.3. Bacterial Interaction with GMP
2.4.4. Cell-Line Interaction with GMP
2.4.5. Instantaneous Effect of GMP
2.5. Translocation and Transepithelial Electrical Resistance (TEER) Analysis
2.6. Statistical Analysis
3. Results and Discussion
3.1. Anti-Adhesive Activity of GMP
- (1)
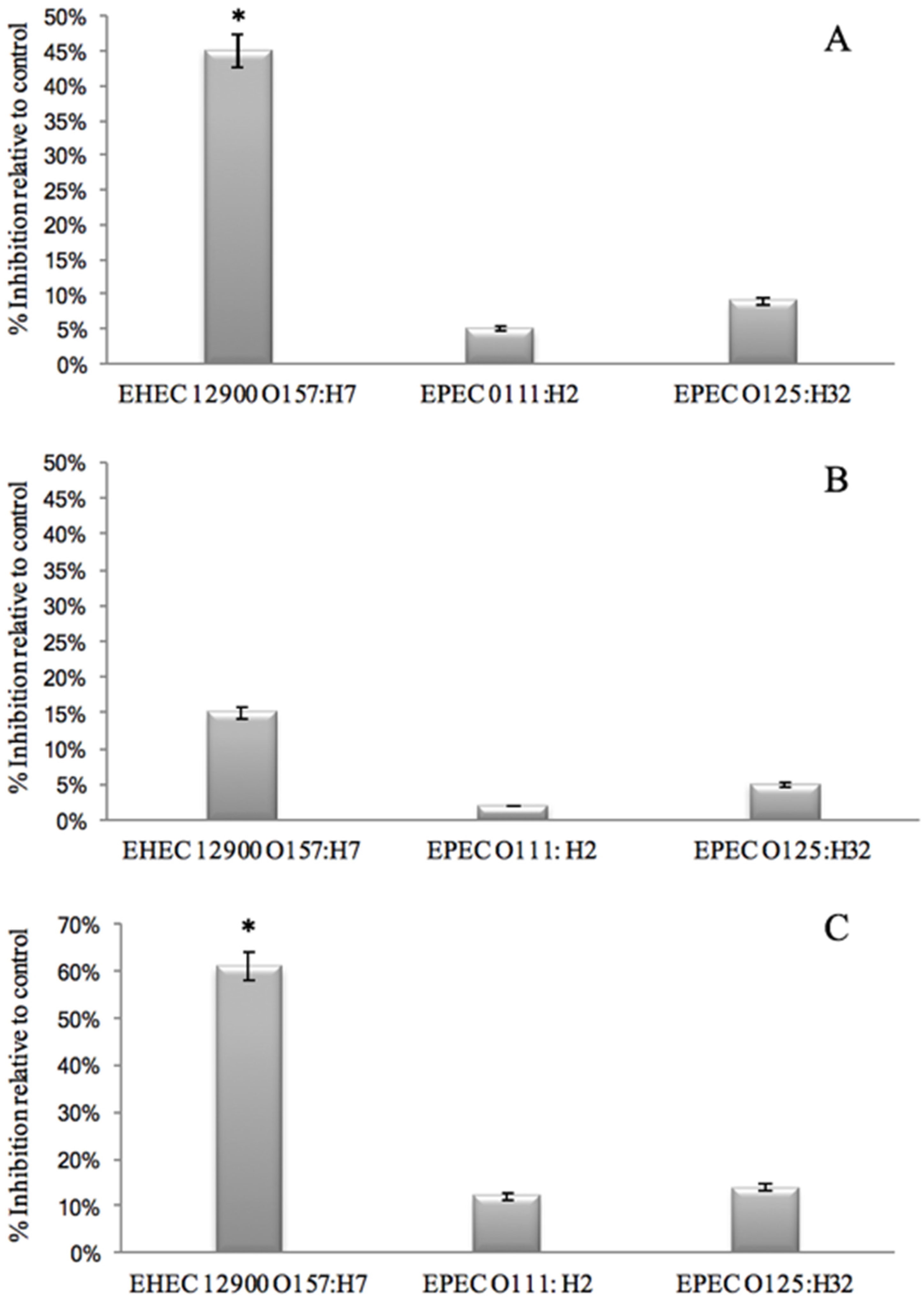
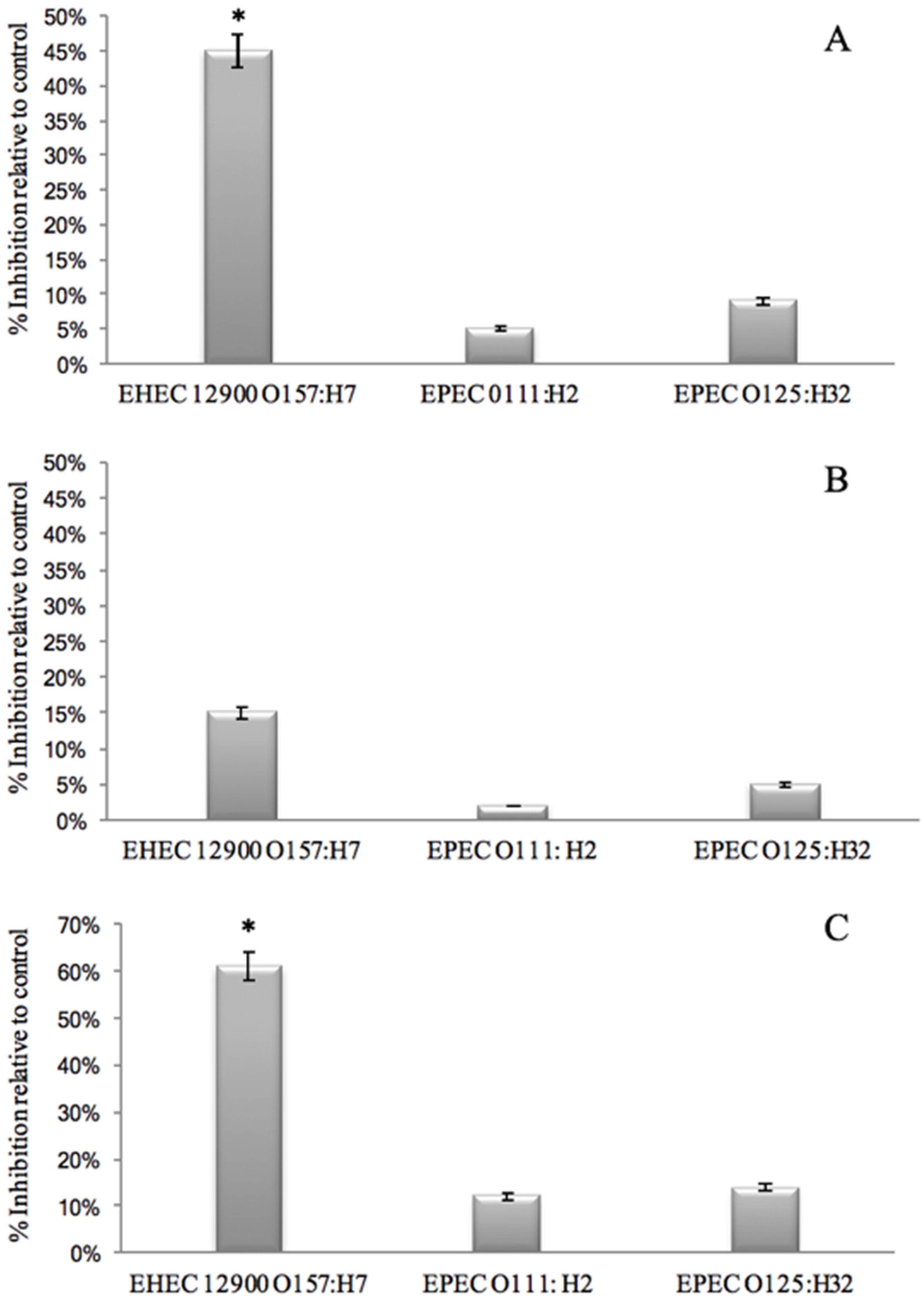
- That GMP interacts with bacterial binding sites, thereby preventing the bacteria from binding to HT29 cell receptors. This was investigated by removing unbound GMP from bacterial GMP mixture using centrifugation prior to infection. Anti-adhesive activity was still observed against EHEC 12900 adhesion to HT29 cells (p < 0.05). However, no significant anti-adhesive activity was evident against EPEC O111 and O125 (Figure 2A).
- (2)
- That GMP binds to epithelial cell receptors, thereby preventing bacteria from interacting with the host cell surface. This was investigated by pre-incubating GMP with the cell line and washing off unbound GMP prior to bacterial challenge. No inhibition was observed against either EHEC or EPEC strains. These results suggest that GMP interacts with the bacteria but not the mammalian cells. It is interesting to note that removing unbound GMP from the bacteria reduced the anti-adhesive activity compared to co-incubating GMP with the bacterial and mammalian cells (Figure 2B).
- (3)
- That pre-incubation of the bacteria with GMP is not required and that inhibition of adhesion can occur with simultaneous exposure of GMP and bacteria to the cell lines as would be more realistic of the in-vivo situation. No anti-infective activity was observed against the EPEC strains; however, an instantaneous anti-infective activity against EHEC 12900 (p < 0.05) was evident (Figure 2C) at levels comparable to using a pre-incubation step (Figure 2A). These results suggest that GMP does not require pre-incubation with the bacteria to exert its maximal inhibitory effect on E. coli cellular association, and a reduction in binding was evident instantaneously. No significant inhibition of adhesion of any of the strains was observed when using the Caco-2 cells in the three experiments outlined above.
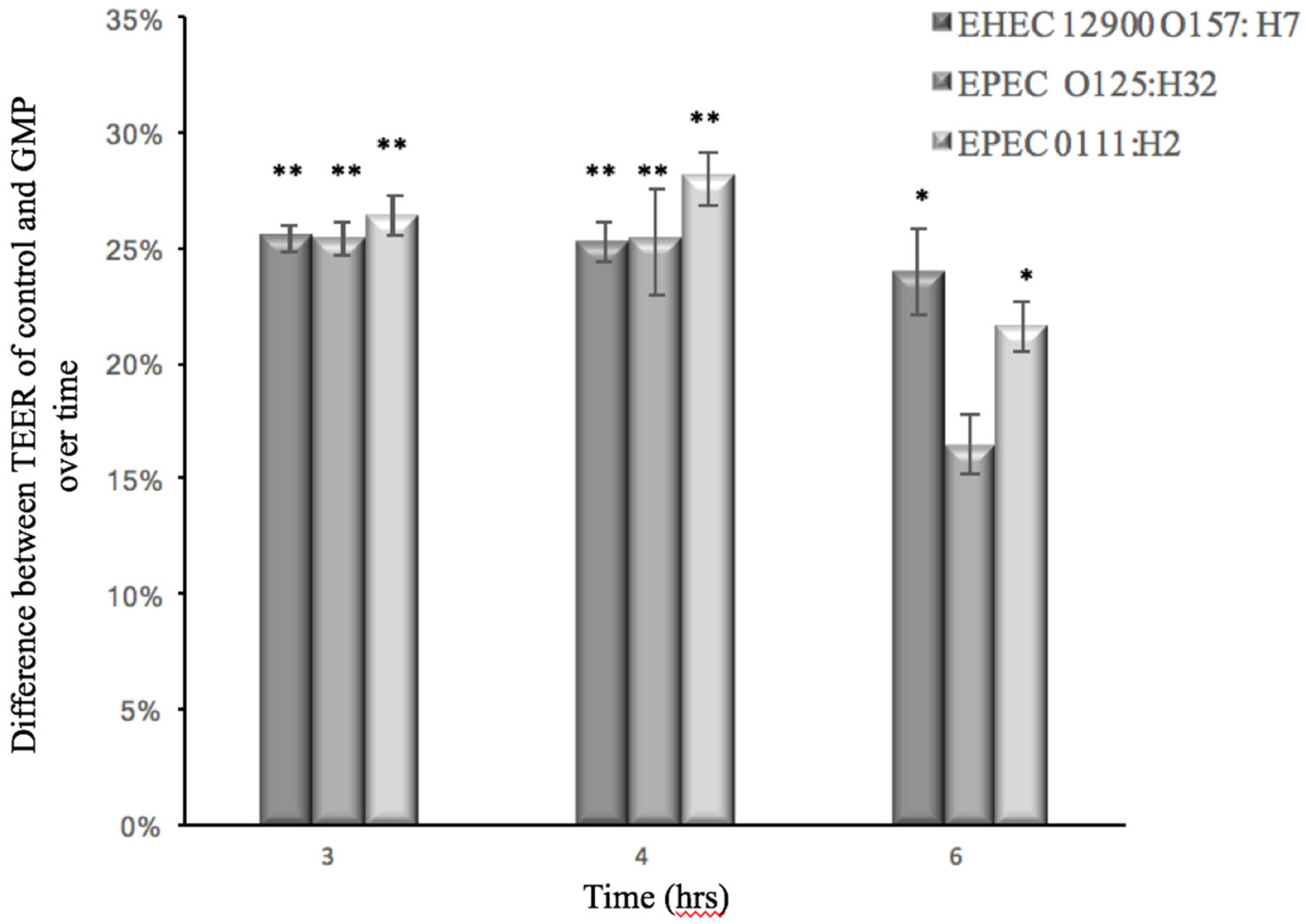
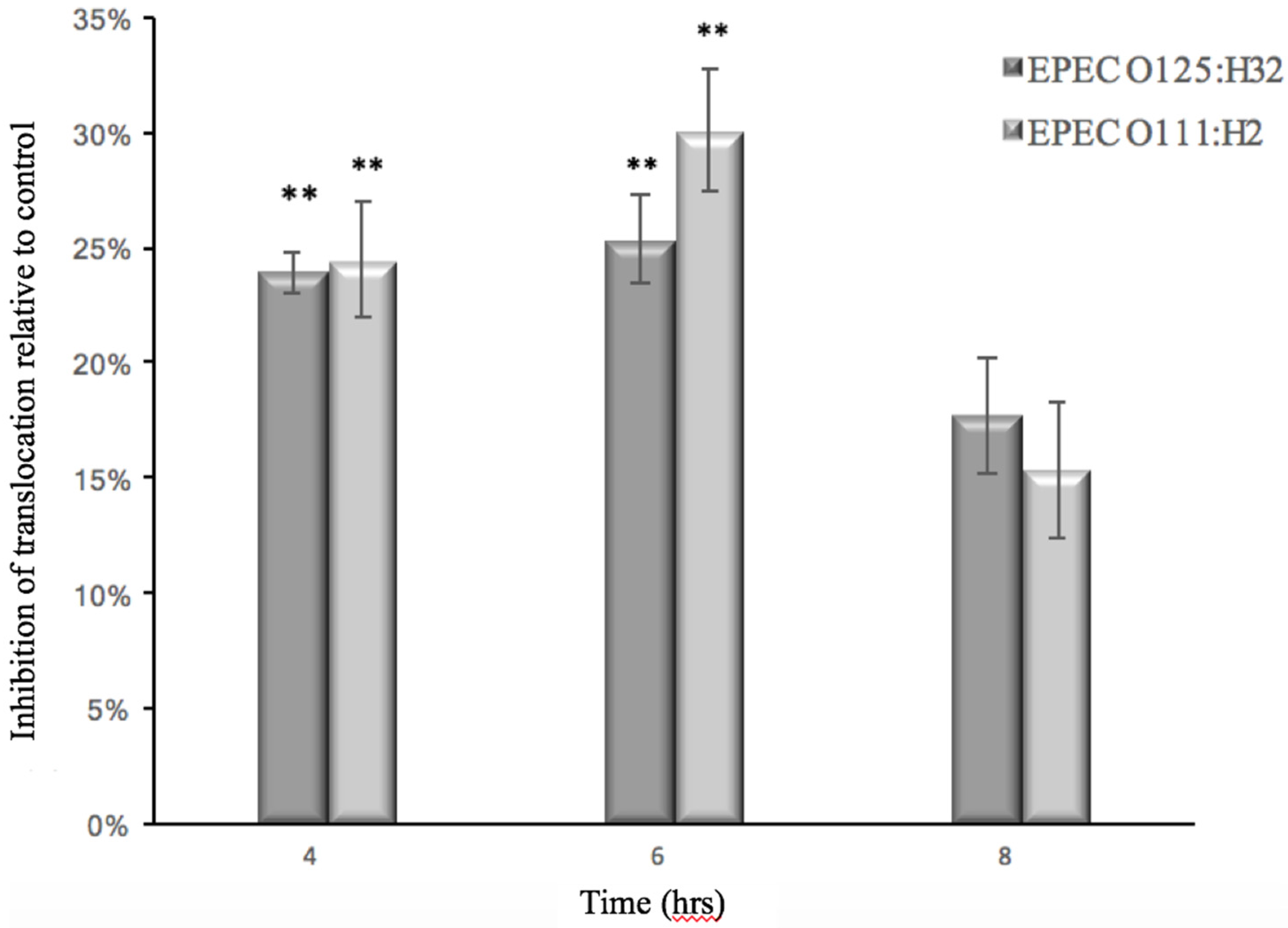
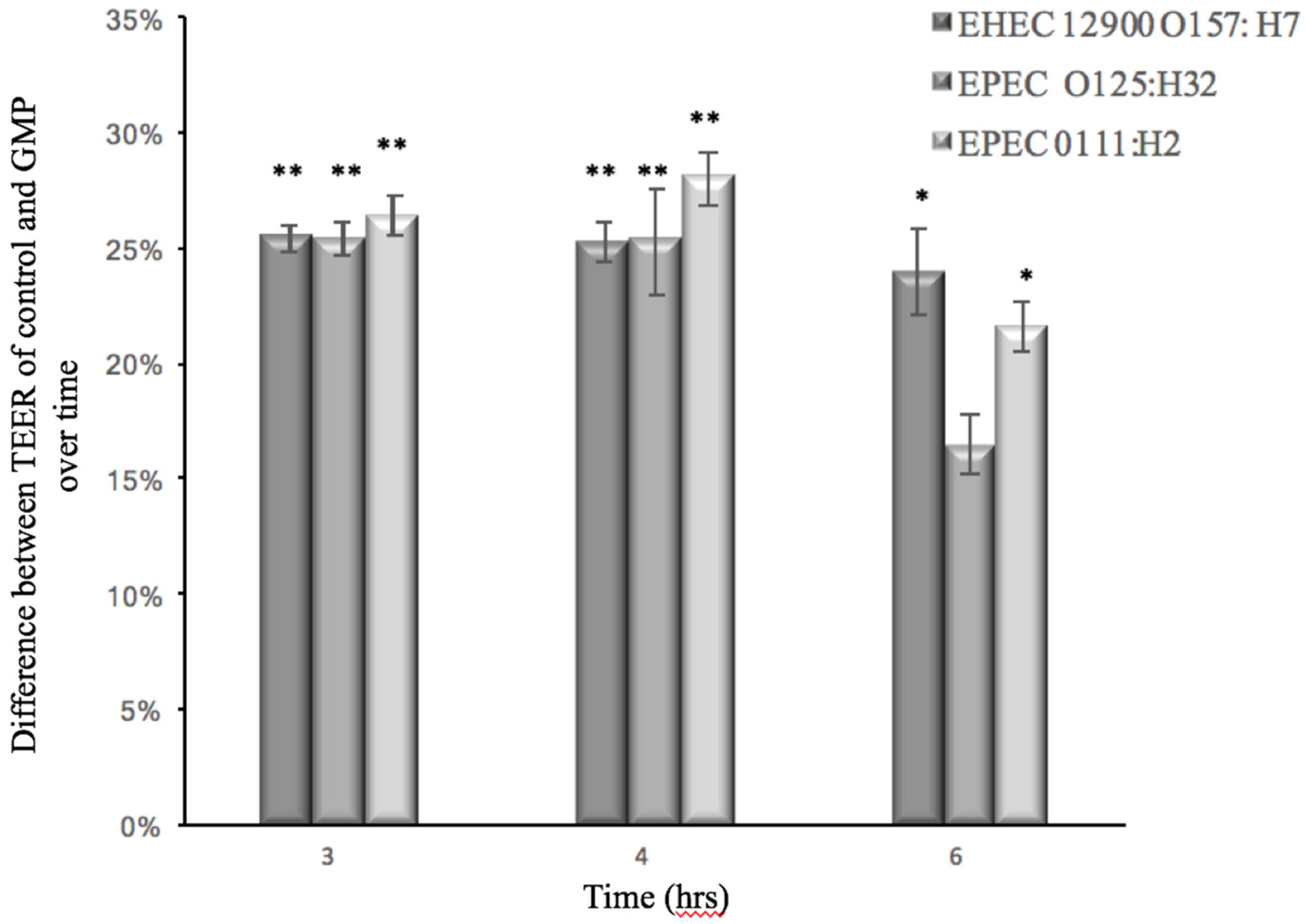
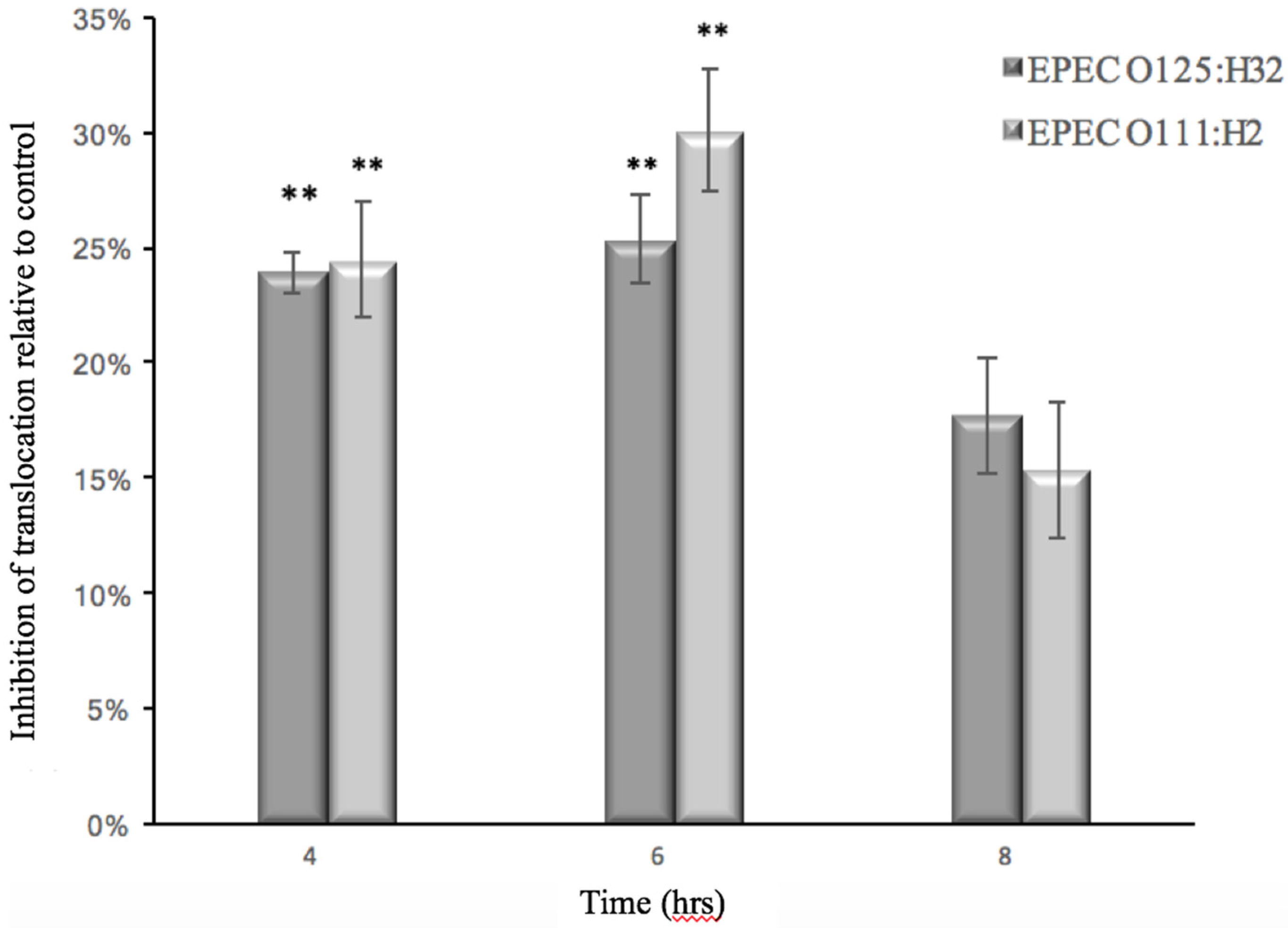
3.2. Effect of GMP on Caco-2 Tight Junction Integrity
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Klemm, P.; Vejborg, R.M.; Hancock, V. Prevention of bacterial adhesion. Appl. Microbiol. Biotechnol. 2010, 88, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Rhoades, J.R.; Gibson, G.R.; Formentin, K.; Beer, M.; Greenberg, N.; Rastall, R.A. Caseinoglycomacropeptide inhibits adhesion of pathogenic Escherichia coli strains to human cells in culture. J. Dairy Sci. 2005, 88, 3455–3459. [Google Scholar] [CrossRef]
- Ofek, I.; Hasty, D.L.; Sharon, N. Anti-adhesion therapy of bacterial diseases: Prospects and problems. FEMS Immunol. Med. Microbiol. 2003, 38, 181–191. [Google Scholar] [CrossRef]
- Wheeler, A.P.; Bernard, G.R. Treating patients with severe sepsis. N. Engl. J. Med. 1999, 340, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Ammori, B.J.; Fitzgerald, P.; Hawkey, P.; McMahon, M.J. The early increase in intestinal permeability and systemic endotoxin exposure in patients with severe acute pancreatitis is not associated with systemic bacterial translocation: Molecular investigation of microbial DNA in the blood. Pancreas 2003, 26, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Deitch, E.A.; Xu, D.Z.; Qi, L.; Berg, R.D. Bacterial translocation from the gut impairs systemic immunity. Surgery 1991, 109, 269–276. [Google Scholar] [PubMed]
- Ciancio, M.J.; Chang, E.B. Epithelial secretory response to inflammation. Ann. N. Y. Acad. Sci. 1992, 664, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Ciancio, M.J.; Vitiritti, L.; Dhar, A.; Chang, E.B. Endotoxin-induced alterations in rat colonic water and electrolyte transport. Gastroenterol 1992, 103, 1437–1443. [Google Scholar] [CrossRef]
- Deitch, E.A.; Specian, R.D.; Berg, R.D. Endotoxin-induced bacterial translocation and mucosal permeability: Role of xanthine oxidase, complement activation, and macrophage products. Crit. Care Med. 1991, 19, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Bernet-Camard, M.F.; Coconnier, M.H.; Hudault, S.; Servin, A.L. Differentiation-associated antimicrobial functions in human colon adenocarcinoma cell lines. Exp. Cell Res. 1996, 226, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Izumikawa, K.; Hirakata, Y.; Yamaguchi, T.; Takemura, H.; Maesaki, S.; Tomono, K.; Igimi, S.; Kaku, M.; Yamada, Y.; Kohno, S.; et al. Escherichia coli O157 interactions with human intestinal Caco-2 cells and the influence of fosfomycin. J. Antimicrob. Chemother. 1998, 42, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Hirotani, Y.; Ikeda, K.; Kato, R.; Myotoku, M.; Umeda, T.; Ijiri, Y.; Tanaka, K. Protective effects of lactoferrin against intestinal mucosal damage induced by lipopolysaccharide in human intestinal Caco-2 cells. Yakugaku Zasshi 2008, 128, 1363–1368. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.L.; Reardon, C.; Wang, A.; Nazli, A.; McKay, D.M. Transforming growth factor-beta regulation of epithelial tight junction proteins enhances barrier function and blocks enterohemorrhagic Escherichia coli O157:H7-induced increased permeability. Am. J. Pathol. 2005, 167, 1587–1597. [Google Scholar] [CrossRef]
- Requena, P.; Gonzalez, R.; Lopez-Posadas, R.; Abadia-Molina, A.; Suarez, M.D.; Zarzuelo, A.; de Medina, F.S.; Martinez-Augustin, O. The intestinal antiinflammatory agent Glycomacropeptide has immunomodulatory actions on rat splenocytes. Biochem. Pharmacol. 2010, 79, 1797–1804. [Google Scholar] [CrossRef] [PubMed]
- Posadas, S.J.; Caz, V.; Caballero, I.; Cendejas, E.; Quilez, I.; Largo, C.; Elvira, M.; De Miguel, E. Effects of mannoprotein E1 in liquid diet on inflammatory response and TLR5 expression in the gut of rats infected by Salmonella typhimurium. BMC Gastroenterol. 2010, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Coppa, G.V.; Zampini, L.; Galeazzi, T.; Facinelli, B.; Ferrante, L.; Capretti, R.; Orazio, G. Human milk oligosaccharides inhibit the adhesion to Caco-2 cells of diarrheal pathogens: Escherichia coli, Vibrio cholerae, and Salmonella fyris. Pediatr. Res. 2006, 59, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Cravioto, A.; Tello, A.; Villafan, H.; Ruiz, J.; del Vedovo, S.; Neeser, J.R. Inhibition of localized adhesion of enteropathogenic Escherichia coli to Hep-2 cells by immunoglobulin and oligosaccharide fractions of human colostrum and breast milk. J. Infect. Dis. 1991, 163, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Simon, P.M.; Goode, P.L.; Mobasseri, A.; Zopf, D. Inhibition of Helicobacter pylori binding to gastrointestinal epithelial cells by sialic acid-containing oligosaccharides. Infect. Immun. 1997, 65, 750–757. [Google Scholar] [PubMed]
- Delfour, A.; Jolles, J.; Alais, C.; Jolles, P. Casein-glycopeptides: Characterization of a methionine residue and of the N-terminal sequence. Biochem. Biophys. Res. Commun. 1965, 19, 425. [Google Scholar] [CrossRef]
- Bruck, W.M.; Kelleher, S.L.; Gibson, G.R.; Graverholt, G.; Lonnerdal, B.L. The effects of alpha-lactalbumin and Glycomacropeptide on the association of Caco-2 cells by enteropathogenic Escherichia coli, Salmonella typhimurium and Shigella flexneri. FEMS Microbiol. Lett. 2006, 259, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Bruck, W.M.; Redgrave, M.; Tuohy, K.M.; Lonnerdal, B.; Graverholt, G.; Hernell, O.; Gibson, G.R. Effects of bovine alpha-lactalbumin and casein Glycomacropeptide-enriched infant formulae on faecal microbiota in healthy term infants. J. Pediatr. Gastroenterol. Nutr. 2006, 43, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Ntemiri, A.; Ní Chonchúir, F.; O’Callaghan, T.F.; Stanton, C.; Ross, R.P.; O’Toole, P.W. Glycomacropeptide sustains microbiota diversity and promotes specific taxa in an artificial colon model of elderly gut microbiota. J. Agric. Food Chem. 2017, 65, 1836–1846. [Google Scholar] [CrossRef] [PubMed]
- Sawin, E.A.; De Wolfe, T.J.; Aktas, B.; Stroup, B.M.; Murali, S.G.; Steele, J.L.; Ney, D.M. Glycomacropeptide is a prebiotic that reduces Desulfovibrio bacteria, increases cecal short-chain fatty acids, and is anti-inflammatory in mice. Am. J. Physiol. Gastrointest Liver Physiol. 2015, 309, G590–G601. [Google Scholar] [CrossRef] [PubMed]
- Brody, E.P. Biological activities of bovine Glycomacropeptide. Br. J. Nutr. 2000, 84 (Suppl. 1), S39–S46. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Itoh, T. Variations and distributions of O-glycosidically linked sugar chains in bovine kappa-casein. J. Dairy Sci. 1992, 75, 1768–1774. [Google Scholar] [CrossRef]
- O’Riordan, N.; Kane, M.; Joshi, L.; Hickey, R.M. Structural and functional characteristics of bovine milk protein glycosylation. Glycobiology 2014, 24, 220–236. [Google Scholar] [CrossRef] [PubMed]
- Severi, E.; Hood, D.W.; Thomas, G.H. Sialic acid utilization by bacterial pathogens. Microbiology 2007, 153, 2817–2822. [Google Scholar] [CrossRef] [PubMed]
- Varki, N.M.; Varki, A. Diversity in cell surface sialic acid presentations: Implications for biology and disease. Lab. Investig. 2007, 87, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Wilbrink, M.H.; ten Kate, G.A.; van Leeuwen, S.S.; Sanders, P.; Sallomons, E.; Hage, J.A.; Dijkhuizen, L.; Kamerling, J.P. Galactosyl-lactose sialylation using Trypanosoma cruzi trans-sialidase as the biocatalyst and bovine kappa-casein-derived Glycomacropeptide as the donor substrate. Appl. Environ. Microbiol. 2014, 80, 5984–5991. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, K.; Tamura, N.; Kobayashi-Hattori, K.; Yoshida, T.; Hara-Kudo, Y.; Ikedo, M.; Sugita-Konishi, Y.; Hattori, M. Prevention of intestinal infection by Glycomacropeptide. Biosci. Biotechnol. Biochem. 2005, 69, 2294–2301. [Google Scholar] [CrossRef] [PubMed]
- Stromqvist, M.; Falk, P.; Bergstrom, S.; Hansson, L.; Lonnerdal, B.; Normark, S.; Hernell, O. Human milk kappa-casein and inhibition of Helicobacter pylori adhesion to human gastric mucosa. J. Pediatr. Gastroenterol. Nutr. 1995, 21, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Ortiz, G.; Hermes, R.G.; Jimenez-Diaz, R.; Perez, J.F.; Martin-Orue, S.M. Screening of extracts from natural feed ingredients for their ability to reduce enterotoxigenic Escherichia coli (ETEC) K88 adhesion to porcine intestinal epithelial cell-line IPEC-j2. Vet. Microbiol. 2013, 167, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Ortiz, G.; Perez, J.F.; Hermes, R.G.; Molist, F.; Jimenez-Diaz, R.; Martin-Orue, S.M. Screening the ability of natural feed ingredients to interfere with the adherence of enterotoxigenic Escherichia coli (ETEC) K88 to the porcine intestinal mucus. Br. J. Nutr. 2014, 111, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Lane, J.A.; Marino, K.; Naughton, J.; Kavanaugh, D.; Clyne, M.; Carrington, S.D.; Hickey, R.M. Anti-infective bovine colostrum oligosaccharides: Campylobacter jejuni as a case study. Int. J. Food Microbiol. 2012, 157, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Horemans, T.; Kerstens, M.; Clais, S.; Struijs, K.; van den Abbeele, P.; Van Assche, T.; Maes, L.; Cos, P. Evaluation of the anti-adhesive effect of milk fat globule membrane glycoproteins on Helicobacter pylori in the human NCI-N87 cell line and C57BL/6 mouse model. Helicobacter 2012, 17, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, J.; Barbera, R.; Matencio, E.; Alegria, A.; Lagarda, M.J. Gangliosides and sialic acid effects upon newborn pathogenic bacteria adhesion: An in vitro study. Food Chem. 2013, 136, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Thoma-Worringer, C.S.J.; Lopez-Findino, R. Health effects and technological features of caseinomacropeptide. Int. Dairy J. 2006, 16, 1324–1333. [Google Scholar] [CrossRef]
- Fairbrother, J.M.; Nadeau, E.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Croxen, M.A.; Law, R.J.; Scholz, R.; Keeney, K.M.; Wlodarska, M.; Finlay, B.B. Recent advances in understanding enteric pathogenic Escherichia coli. Clin. Microbiol. Rev. 2013, 26, 822–880. [Google Scholar] [CrossRef] [PubMed]
- Sharon, N.; Ofek, I. Safe as mother’s milk: Carbohydrates as future anti-adhesion drugs for bacterial diseases. Glycoconj. J. 2000, 17, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Spears, K.J.; Roe, A.J.; Gally, D.L. A comparison of enteropathogenic and enterohaemorrhagic Escherichia coli pathogenesis. FEMS Microbiol. Lett. 2006, 255, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Neelima; Sharma, R.; Rajput, Y.S.; Mann, B. Chemical and functional properties of Glycomacropeptide (GMP) and its role in the detection of cheese whey adulteration in milk: A review. Dairy Sci. Technol. 2013, 93, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Matthews, L.; Reeve, R.; Gally, D.L.; Low, J.C.; Woolhouse, M.E.; McAteer, S.P.; Locking, M.E.; Chase-Topping, M.E.; Haydon, D.T.; Allison, L.J.; et al. Predicting the public health benefit of vaccinating cattle against Escherichia coli O157. Proc. Natl. Acad. Sci. USA 2013, 110, 16265–16270. [Google Scholar] [CrossRef] [PubMed]
- Zahavi, E.E.; Lieberman, J.A.; Donnenberg, M.S.; Nitzan, M.; Baruch, K.; Rosenshine, I.; Turner, J.R.; Melamed-Book, N.; Feinstein, N.; Zlotkin-Rivkin, E.; et al. Bundle-forming pilus retraction enhances enteropathogenic Escherichia coli infectivity. Mol. Biol. Cell 2011, 22, 2436–2447. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.; Hanajima, M.; Matsuzawa, T.; Kobayashi, C.; Minami, M.; Abe, A.; Horiguchi, Y. Binding of intimin with TIR on the bacterial surface is prerequisite for the barrier disruption induced by enteropathogenic Escherichia coli. Biochem. Biophys. Res. Commun. 2005, 337, 922–927. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa, T.; Kuwae, A.; Abe, A. Enteropathogenic Escherichia coli type III effectors EspG and EspG2 alter epithelial paracellular permeability. Infect. Immun. 2005, 73, 6283–6289. [Google Scholar] [CrossRef] [PubMed]
- Bernet-Camard, M.F.; Coconnier, M.H.; Hudault, S.; Servin, A.L. Differential expression of complement proteins and regulatory decay accelerating factor in relation to differentiation of cultured human colon adenocarcinoma cell lines. Gut 1996, 38, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Welinder-Olsson, C.; Eriksson, E.; Kaijser, B. Virulence genes in verocytotoxigenic Escherichia coli strains isolated from humans and cattle. APMIS 2005, 113, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Welinder-Olsson, C.; Kaijser, B. Enterohemorrhagic Escherichia coli (EHEC). Scand. J. Infect. Dis. 2005, 37, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Glotfelty, L.G.; Hecht, G.A. Enteropathogenic E. coli effectors ESPG1/G2 disrupt tight junctions: New roles and mechanisms. Ann. N. Y. Acad. Sci. 2012, 1258, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Izumikawa, K.; Hirakata, Y.; Yamaguchi, T.; Yoshida, R.; Nakano, M.; Matsuda, J.; Mochida, C.; Maesaki, S.; Tomono, K.; Yamada, Y.; et al. Analysis of genetic relationships and antimicrobial susceptibility of verotoxin-producing Escherichia coli strains isolated in Nagasaki prefecture, Japan in 1996. Microbiol. Immunol. 1998, 42, 677–681. [Google Scholar] [CrossRef] [PubMed]
- Luck, S.N.; Turner, S.A.; Rajakumar, K.; Adler, B.; Sakellaris, H. Excision of the Shigella resistance locus pathogenicity island in Shigella flexneri is stimulated by a member of a new subgroup of recombination directionality factors. J. Bacteriol. 2004, 186, 5551–5554. [Google Scholar] [CrossRef] [PubMed]
- Savkovic, S.D.; Koutsouris, A.; Hecht, G. Activation of NF-kappaB in intestinal epithelial cells by enteropathogenic Escherichia coli. Am. J. Physiol. 1997, 273, C1160–C1167. [Google Scholar] [PubMed]
- Savkovic, S.D.; Ramaswamy, A.; Koutsouris, A.; Hecht, G. EPEC-activated ERK1/2 participate in inflammatory response but not tight junction barrier disruption. Am. J. Physiol. Gastrointest Liver Physiol. 2001, 281, G890–G898. [Google Scholar] [PubMed]
- Lang, A.; Lahav, M.; Sakhnini, E.; Barshack, I.; Fidder, H.H.; Avidan, B.; Bardan, E.; Hershkoviz, R.; Bar-Meir, S.; Chowers, Y. Allicin inhibits spontaneous and TNF-alpha induced secretion of proinflammatory cytokines and chemokines from intestinal epithelial cells. Clin. Nutr. 2004, 23, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Mukaida, N.; Okamoto, S.; Ishikawa, Y.; Matsushima, K. Molecular mechanism of Interleukin-8 gene expression. J. Leukoc. Biol. 1994, 56, 554–558. [Google Scholar] [PubMed]
- Al-Sadi, R.; Ye, D.; Dokladny, K.; Ma, T.Y. Mechanism of IL-1beta-induced increase in intestinal epithelial tight junction permeability. J. Immunol. 2008, 180, 5653–5661. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.Y.; Iwamoto, G.K.; Hoa, N.T.; Akotia, V.; Pedram, A.; Boivin, M.A.; Said, H.M. TNF-alpha-induced increase in intestinal epithelial tight junction permeability requires NF-kappaB activation. Am. J. Physiol. Gastrointest Liver Physiol. 2004, 286, G367–G376. [Google Scholar] [CrossRef] [PubMed]
- Ye, D.; Ma, I.; Ma, T.Y. Molecular mechanism of tumor necrosis factor-alpha modulation of intestinal epithelial tight junction barrier. Am. J. Physiol. Gastrointest Liver Physiol. 2006, 290, G496–G504. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Chen, Q.; Yan, Y.; Pang, G. Effect of casein Glycomacropeptide on subunit p65 of nuclear transcription factor-κB in lipopolysaccharide-stimulated human colorectal tumor HT-29 cells. Food Sci. Hum. Wellness 2014, 3, 51–55. [Google Scholar] [CrossRef]
- Rong, Y.; Lu, Z.; Zhang, H.; Zhang, L.; Song, D.; Wang, Y. Effects of casein Glycomacropeptide supplementation on growth performance, intestinal morphology, intestinal barrier permeability and inflammatory responses in Escherichia coli K88 challenged piglets. Anim. Nutr. 2015, 1, 54–59. [Google Scholar] [CrossRef]
- Yuhan, R.; Koutsouris, A.; Savkovic, S.D.; Hecht, G. Enteropathogenic Escherichia coli-induced myosin light chain phosphorylation alters intestinal epithelial permeability. Gastroenterology 1997, 113, 1873–1882. [Google Scholar] [CrossRef]
- Bellmeyer, A.; Cotton, C.; Kanteti, R.; Koutsouris, A.; Viswanathan, V.K.; Hecht, G. Enterohemorrhagic Escherichia coli suppresses inflammatory response to cytokines and its own toxin. Am. J. Physiol. Gastrointest Liver Physiol. 2009, 297, G576–G581. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, R.M.; Ma, T.Y. IL-1beta causes an increase in intestinal epithelial tight junction permeability. J. Immunol. 2007, 178, 4641–4649. [Google Scholar] [CrossRef] [PubMed]
- Daddaoua, A.; Puerta, V.; Zarzuelo, A.; Suarez, M.D.; Sanchez de Medina, F.; Martinez-Augustin, O. Bovine Glycomacropeptide is anti-inflammatory in rats with hapten-induced colitis. J. Nutr. 2005, 135, 1164–1170. [Google Scholar] [PubMed]
- Monnai, M.O.H. Effect of bovine k-caseinoglycopeptide on secretion of interleukin-1 family cytokines by P388D1 cells, a line derived from mouse monocyte/macrophage. Milchwissenschaft 1997, 52, 192–196. [Google Scholar]
- Hvas, C.L.; Dige, A.; Bendix, M.; Wernlund, P.G.; Christensen, L.A.; Dahlerup, J.F.; Agnholt, J. Casein Glycomacropeptide for active distal ulcerative colitis: A randomized pilot study. Eur. J. Clin. Investig. 2016, 46, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Kreuß, M.; Krause, I.; Kulozik, U. Separation of a glycosylated and non-glycosylated fraction of caseinomacropeptide using different anion-exchange stationary phases. J. Chromatogr. A 2008, 1208, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Kreuß, M.; Krause, I.; Kulozik, U. Influence of glycosylation on foaming properties of bovine caseinomacropeptide. Int. Dairy J. 2009, 19, 715–720. [Google Scholar] [CrossRef]
- Kreuß, M.; Strixner, T.; Kulozik, U. The effect of glycosylation in the interfacial properties of bovine caseinomacropeptide. Food Hydrocoll. 2009, 23, 1818–1826. [Google Scholar] [CrossRef]
- Kreuß, M.; Kulozik, U. Separation of glycosylated caseinomacropeptide at pilot scale using membrane adsorption in direct-capture mode. J. Chromatogr. A 2009, 1216, 8771–8777. [Google Scholar] [CrossRef] [PubMed]
- Koliwer-Brandl, H.; Siegert, N.; Umus, K.; Kelm, A.; Tolkach, A.; Kulozik, U.; Kuballa, J.; Cartellieri, S.; Kelm, S. Lectin inhibition assays for the analysis of bioactive milk sialoglycoconjugates. Int. Dairy J. 2011, 21, 413–420. [Google Scholar] [CrossRef]
- Siegert, N.; Tolkach, A.; Kulozik, U. The pH-dependent thermal and storage stability of glycosylated caseinomacropeptide. LWT-Food Sci. Technol. 2012, 47, 407–412. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| E. coli strain | % Inhibition | |
|---|---|---|
| HT-29 | Caco-2 | |
| EHEC 12900 O157:H7 | 70 *** | 62 *** |
| EPEC O111:H2 | 26 * | 25 * |
| EPEC O125:H32 | 24 * | 25 * |
| EHEC DAF 454 | 21 | N/T |
| EHEC DPC 6055 | 15 | N/T |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feeney, S.; Ryan, J.T.; Kilcoyne, M.; Joshi, L.; Hickey, R. Glycomacropeptide Reduces Intestinal Epithelial Cell Barrier Dysfunction and Adhesion of Entero-Hemorrhagic and Entero-Pathogenic Escherichia coli in Vitro. Foods 2017, 6, 93. https://doi.org/10.3390/foods6110093
Feeney S, Ryan JT, Kilcoyne M, Joshi L, Hickey R. Glycomacropeptide Reduces Intestinal Epithelial Cell Barrier Dysfunction and Adhesion of Entero-Hemorrhagic and Entero-Pathogenic Escherichia coli in Vitro. Foods. 2017; 6(11):93. https://doi.org/10.3390/foods6110093
Chicago/Turabian StyleFeeney, Shane, Joseph Thomas Ryan, Michelle Kilcoyne, Lokesh Joshi, and Rita Hickey. 2017. "Glycomacropeptide Reduces Intestinal Epithelial Cell Barrier Dysfunction and Adhesion of Entero-Hemorrhagic and Entero-Pathogenic Escherichia coli in Vitro" Foods 6, no. 11: 93. https://doi.org/10.3390/foods6110093
APA StyleFeeney, S., Ryan, J. T., Kilcoyne, M., Joshi, L., & Hickey, R. (2017). Glycomacropeptide Reduces Intestinal Epithelial Cell Barrier Dysfunction and Adhesion of Entero-Hemorrhagic and Entero-Pathogenic Escherichia coli in Vitro. Foods, 6(11), 93. https://doi.org/10.3390/foods6110093