

Low-Molecular-Weight Fucoidan from Undaria pinnatifida Mitigates Salmonella-Induced Injury Through Gut Microbiota and Immune Regulation

 ,
,  ,
,
Abstract
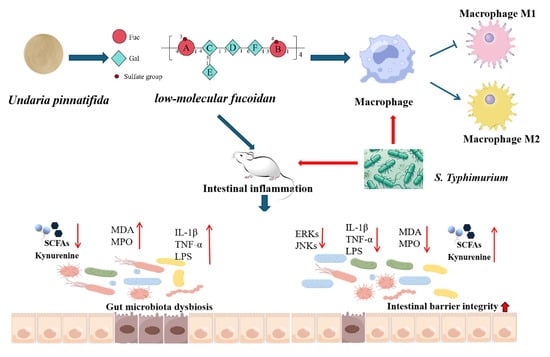
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Salmonella Strains and Chemical Reagents
2.2. Preparation of LUPF
2.3. Structural Characterization of LUPF
2.3.1. Chemical Composition of LUPF
2.3.2. Monosaccharide Composition
2.3.3. Determination of Mw
2.3.4. Desulfation of LUPF
2.3.5. Fourier-Transform Infrared Spectroscopy (FT-IR)
2.3.6. Nuclear Magnetic Resonance (NMR) Spectroscopy
2.3.7. Methylation Analysis
2.4. Preparation of S. typhimurium (ST) Suspension
2.5. Mice Experiment
2.6. Quantification of S. typhimurium in Organ Tissues
2.7. Histological Analysis
2.8. Measurement of Biochemical Parameters
2.9. Measurement of Short-Chain Fatty Acids (SCFAs)
2.10. Sequencing Analysis of the Fecal Microbiota
2.11. Metabolomic Analysis of Serum Samples
2.12. Effect of LUPF on ST-Induced Inflammation In Vitro
2.12.1. Cell Culture
2.12.2. Immunofluorescence Staining
2.12.3. Quantitative Real-Time PCR
2.13. Western Blotting
2.14. Statistical Analysis
3. Results and Discussion
3.1. Structural Characterization of LUPF
3.1.1. General Structural Features of LUPF
3.1.2. FT-IR Analysis
3.1.3. Methylation and NMR Analysis
3.2. LUPF Alleviated ST-Induced Multi-Organ Damage
3.3. LUPF Improved ST-Induced Serum Metabolic Abnormalities
3.4. LUPF Protected Intestinal Barrier Integrity in ST-Treated Mice
3.5. LUPF Alleviated ST Infection-Induced Inflammation and Oxidative Stress
3.6. LUPF Modulated ST Infection-Induced Gut Microbiota Dysbiosis
3.7. LUPF Promoted the Production of Intestinal SCFAs
3.8. LUPF Alleviated Inflammation by Regulating Macrophage Polarization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coburn, B.; Grassl, G.A.; Finlay, B.B. Salmonella, the host and disease: A brief review. Immunol. Cell Biol. 2007, 85, 112–118. [Google Scholar] [CrossRef]
- Fierer, J.; Guiney, D.G. Diverse virulence traits underlying different clinical outcomes of salmonella infection. J. Clin. Investig. 2001, 107, 775–780. [Google Scholar] [CrossRef]
- Khan, M.; Shamim, S. Understanding the mechanism of antimicrobial resistance and pathogenesis of salmonella enterica serovar typhi. Microorganisms 2022, 10, 2006. [Google Scholar] [CrossRef]
- Gut, A.M.; Vasiljevic, T.; Yeager, T.; Donkor, O.N. Salmonella infection—Prevention and treatment by antibiotics and probiotic yeasts: A review. Microbiology 2018, 164, 1327–1344. [Google Scholar] [CrossRef] [PubMed]
- Kamada, N.; Chen, G.Y.; Inohara, N.; Núñez, G. Control of pathogens and pathobionts by the gut microbiota. Nat. Immunol. 2013, 14, 685–690. [Google Scholar] [CrossRef]
- Kim, J.-S.; Kim, Y. The inhibitory effect of natural bioactives on the growth of pathogenic bacteria. Nutr. Res. Pract. 2007, 1, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zheng, Y.; Wang, J.; Ma, S.; Yu, Y.; White, W.L.; Yang, S.; Yang, F.; Lu, J. Fucoidan extracted from undaria pinnatifida: Source for nutraceuticals/functional foods. Mar. Drugs 2018, 16, 321. [Google Scholar] [CrossRef]
- Synytsya, A.; Kim, W.J.; Kim, S.M.; Pohl, R.; Synytsya, A.; Kvasnička, F.; Čopíková, J.; Park, Y.I. Structure and antitumour activity of fucoidan isolated from sporophyll of korean brown seaweed undaria pinnatifida. Carbohydr. Polym. 2010, 81, 41–48. [Google Scholar] [CrossRef]
- Lee, H.H.; Cho, Y.J.; Kim, G.H.; Cho, H. Undaria pinnatifida fucoidan-rich extract recovers immunity of immunosuppressed mice. J. Microbiol. Biotechnol. 2020, 30, 439–447. [Google Scholar] [CrossRef]
- Wang, L.; Park, Y.J.; Jeon, Y.J.; Ryu, B. Bioactivities of the edible brown seaweed, undaria pinnatifida: A review. Aquaculture 2018, 495, 873–880. [Google Scholar] [CrossRef]
- Men, Q.; Zhang, P.; Zheng, W.; Song, S.; Ai, C. Fucoidan alleviates salmonella-induced inflammation and mortality by modulating gut microbiota and metabolites, protecting intestinal barrier, and inhibiting nf-κb pathway. Food Biosci. 2023, 56, 103209. [Google Scholar] [CrossRef]
- Choi, J.-I.; Kim, H.-J. Preparation of low molecular weight fucoidan by gamma-irradiation and its anticancer activity. Carbohydr. Polym. 2013, 97, 358–362. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Guo, F.; Hu, J.; Zhang, L.; Xue, C.; Zhang, Z.; Li, B. Antithrombotic activity of oral administered low molecular weight fucoidan from laminaria japonica. Thromb. Res. 2016, 144, 46–52. [Google Scholar] [CrossRef]
- Yang, C.; Dwan, C.; Wimmer, B.C.; Ahamed, S.K.; James, F.; Thinley, J.; Wilson, R.; Johnson, L.; Caruso, V. Anti-inflammatory and neuroprotective effects of undaria pinnatifida fucoidan. Mar. Drugs 2025, 23, 350. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Kuo, C.-H.; Lee, C.-H. Antibacterial and antioxidant capacities and attenuation of lipid accumulation in 3t3-l1 adipocytes by low-molecular-weight fucoidans prepared from compressional-puffing-pretreated sargassum crassifolium. Mar. Drugs 2018, 16, 24. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, T.; Eapen, M.S.; Ishaq, M.; Park, A.Y.; Karpiniec, S.S.; Stringer, D.N.; Sohal, S.S.; Fitton, J.H.; Guven, N.; Caruso, V.; et al. Anti-inflammatory activity of fucoidan extracts in vitro. Mar. Drugs 2021, 19, 702. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wang, J.; Zhang, P.; Jiang, Z.; Chen, S.; Liang, S.; Ma, T.; Liao, H.; Tan, W.; Niu, C.; et al. Low molecular weight fucoidan induces m2 macrophage polarization to attenuate inflammation through activation of the ampk/mtor autophagy pathway. Eur. J. Pharmacol. 2025, 986, 177134. [Google Scholar] [CrossRef]
- Qiu, Z.; Xiang, L.; Han, Y.; Zhang, B.; Qiao, X.; Zheng, Z.; Xiao, H. Structure-anti-inflammatory activity relationship of garlic fructans in mice with dextran sulfate sodium-induced colitis: Impact of chain length. Carbohydr. Polym. 2024, 346, 122582. [Google Scholar] [CrossRef]
- Gao, N.; Chen, R.; Mou, R.R.; Xiang, J.Y.; Zhou, K.; Li, Z.; Zhao, J.H. Purification, structural characterization and anticoagulant activities of four sulfated polysaccharides from sea cucumber holothuria fuscopunctata. Int. J. Biol. Macromol. 2020, 164, 3421–3428. [Google Scholar] [CrossRef]
- Yang, Z.; Xu, J.; Fu, Q.; Fu, X.; Shu, T.; Bi, Y.; Song, B. Antitumor activity of a polysaccharide from pleurotus eryngii on mice bearing renal cancer. Carbohydr. Polym. 2013, 95, 615–620. [Google Scholar] [CrossRef]
- Yan, T.; Zhang, Y.; Lu, H.; Zhao, J.; Wen, C.; Song, S.; Ai, C.; Yang, J. The protective effect of enteromorpha prolifera polysaccharide on alcoholic liver injury in c57bl/6 mice. Int. J. Biol. Macromol. 2024, 261, 129908. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.S.; Chen, F.; Huang, G.L. Extraction and analysis of polysaccharide from Momordica charantia. Ind. Crops Prod. 2020, 153, 112588. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Y.H.; Yang, S.; Lv, Z.H. Separation, purification, structures and anticoagulant activities of fucosylated chondroitin sulfates from Holothuria scabra. Int. J. Biol. Macromol. 2018, 108, 710–718. [Google Scholar] [CrossRef]
- Yang, J.; Zhu, B.; Zheng, J.; Sun, L.; Zhou, D.; Dong, X.; Yu, C. Stimulation of lymphocyte proliferation by oyster glycogen sulfated at c-6 position. Carbohydr. Polym. 2013, 94, 301–308. [Google Scholar] [CrossRef]
- Song, S.; Zhang, B.; Wu, S.; Huang, L.; Ai, C.; Pan, J.; Su, Y.-C.; Wang, Z.; Wen, C. Structural characterization and osteogenic bioactivity of a sulfated polysaccharide from pacific abalone (haliotis discus hannai Ino). Carbohydr. Polym. 2018, 182, 207–214. [Google Scholar] [CrossRef]
- Li, J.; Liu, H.Z.; Fu, H.Y.; Yang, Y.; Wu, Z.L. An isofibrous diet with fiber konjac glucomannan ameliorates salmonella typhimurium-induced colonic injury by regulating tlr2-nf-κb signaling and intestinal microbiota in mice. J. Agric. Food Chem. 2024, 72, 13415–13430. [Google Scholar] [CrossRef]
- You, Y.; Song, H.; Wang, L.; Liu, Z.; Guo, X.; Ai, C.; Song, S.; Zhu, B. Supplement of Caulerpa lentillifera polysaccharide by pre-prandial gavage and free feeding demonstrates differences to prevent obesity and gut microbiota disturbance in mice. J. Sci. Food Agric. 2022, 103, 3840–3849. [Google Scholar] [CrossRef]
- Yorulmaz, E.; Yorulmaz, H.; Gökmen, E.S.; Altınay, S.; Küçük, S.H.; Zengi, O.; Çelik, D.S.; Şit, D. Therapeutic effectiveness of rectally administered fish oil and mesalazine in trinitrobenzenesulfonic acid-induced colitis. Biomed. Pharmacother. 2019, 118, 109247. [Google Scholar] [CrossRef]
- Bai, D.; Sun, T.; Zhao, J.; Du, J.; Bu, X.; Cao, W.; Zhao, Y.; Lu, N. Oroxylin a maintains the colonic mucus barrier to reduce disease susceptibility by reconstituting a dietary fiber-deprived gut microbiota. Cancer Lett. 2021, 515, 73–85. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, F.; Cai, Z.; Zhou, Q.; Chen, B.; Zhang, C.; Liu, H.; Hong, L.; Zhang, T.; Zhong, J.; et al. Improving drug utilization platform with injectable mucoadhesive hydrogel for treating ulcerative colitis. Chem. Eng. J. 2021, 424, 130464. [Google Scholar] [CrossRef]
- Sun, Y.Y.; Men, Q.Y.; Ren, X.M.; Yan, C.H.; Song, S.; Ai, C.Q. Low molecular fucoidan alleviated alcohol-induced liver injury in balb/c mice by regulating the gut microbiota-bile acid-liver axis. Int. J. Biol. Macromol. 2024, 282, 136930. [Google Scholar] [CrossRef]
- Cao, C.; Wang, L.L.; Ai, C.Q.; Gong, G.P.; Wang, Z.F.; Huang, L.J.; Song, S.; Zhu, B.W. Impact of Lycium barbarum arabinogalactan on the fecal metabolome in a dss-induced chronic colitis mouse model. Food Funct. 2022, 13, 8703–8716. [Google Scholar] [CrossRef]
- Jia, J.; Zheng, W.; Tang, S.; Song, S.; Ai, C. Scytosiphon lomentaria fucoidan ameliorates dss-induced colitis in dietary fiber-deficient mice via modulating the gut microbiota and inhibiting the tlr4/nf-κb/mlck pathway. Int. J. Biol. Macromol. 2023, 253, 127337. [Google Scholar] [CrossRef]
- Dong, N.; Li, X.; Xue, C.; Wang, C.; Xu, X.; Bi, C.; Shan, A.; Li, D. Astragalus polysaccharides attenuated inflammation and balanced the gut microflora in mice challenged with Salmonella typhimurium. Int. Immunopharmacol. 2019, 74, 105681. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.N.; Liu, Y.L.; Jiang, P.R.; Song, S.; Ai, C.Q. Interaction of sulfated polysaccharides with intestinal bacteroidales plays an important role in its biological activities. Int. J. Biol. Macromol. 2021, 168, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.Q.; Wang, Y.; Wei, X.R.; Yang, L.; Liu, S.; Wang, Y.; Xu, Y.; Wang, Z.Y.; Zhang, C.; Zhang, M.; et al. Improvement in phenotype homeostasis of macrophages by chitosan nanoparticles and subsequent impacts on liver injury and tumor treatment. Carbohydr. Polym. 2022, 277, 118891. [Google Scholar] [CrossRef]
- Cao, Y.F.; Wang, H.; Sun, Y.; Tong, B.B.; Shi, W.Q.; Peng, L.; Zhang, Y.M.; Wu, Y.Q.; Fu, T.; Zou, H.Y.; et al. Nuclear anln regulates transcription initiation related pol ii clustering and target gene expression. Nat. Commun. 2025, 16, 1271. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.H.; Zheng, W.Y.; Zhang, C.X.; Zhang, P.P.; Guo, X.M.; Song, S.; Ai, C.Q. Fucoidan from Scytosiphon lomentaria protects against destruction of intestinal barrier, inflammation and lipid abnormality by modulating the gut microbiota in dietary fibers-deficient mice. Int. J. Biol. Macromol. 2023, 224, 556–567. [Google Scholar] [CrossRef]
- Chen, Y.; Mao, W.J.; Gao, Y.; Teng, X.C.; Zhu, W.M.; Chen, Y.L.; Zhao, C.Q.; Li, N.; Wang, C.Y.; Yan, M.X.; et al. Structural elucidation of an extracellular polysaccharide produced by the marine fungus Aspergillus versicolor. Carbohydr. Polym. 2013, 93, 478–483. [Google Scholar] [CrossRef]
- Ma, G.Y.; Hou, L.K.; Guo, J.H.; Sun, J.C.; Nong, Q.N.; Wang, Y.F.; Hu, S.W.; Zhao, W.J.; Tan, J.; Liu, X.F.; et al. The structures of two polysaccharides from fructus corni and their protective effect on insulin resistance. Carbohydr. Polym. 2025, 353, 123290. [Google Scholar] [CrossRef]
- Li, R.; Zhou, Q.L.; Yang, R.Y.; Chen, S.T.; Ding, R.; Liu, X.F.; Luo, L.X.; Xia, Q.Y.; Zhong, S.Y.; Qi, Y.; et al. Determining the potent immunostimulation potential arising from the heteropolysaccharide structure of a novel fucoidan, derived from Sargassum zhangii. Food Chem. X 2023, 18, 100712. [Google Scholar] [CrossRef]
- Li, N.; Liu, X.; He, X.X.; Wang, S.Y.; Cao, S.J.; Xia, Z.; Xian, H.L.; Qin, L.; Mao, W.J. Structure and anticoagulant property of a sulfated polysaccharide isolated from the green seaweed monostroma angicava. Carbohydr. Polym. 2017, 159, 195–206. [Google Scholar] [CrossRef]
- Luo, D.H.; Yuan, X.M.; Zeng, Y.W.; Nie, K.Y.; Li, Z.M.; Wang, Z.J. Structure elucidation of a major fucopyranose-rich heteropolysaccharide (stp-ii) from sargassum thunbergii. Carbohydr. Polym. 2016, 143, 1–8. [Google Scholar] [CrossRef]
- Pang, J.; Wang, S.; Wang, Z.; Wu, Y.; Zhang, X.; Pi, Y.; Han, D.; Zhang, S.; Wang, J. Xylo-oligosaccharide alleviates salmonella induced inflammation by stimulating bifidobacterium animalis and inhibiting salmonella colonization. FASEB J. 2021, 35, e21977. [Google Scholar] [CrossRef]
- Richards, P.J.; Almutrafy, A.; Liang, L.; Lafontaine, G.M.F.; King, E.; Fish, N.M.; Connerton, A.J.; Connerton, P.L.; Connerton, I.F. Prebiotic galactooligosaccharide feed modifies the chicken gut microbiota to efficiently clear Salmonella. mSystems 2024, 9, e00754-24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Sun, H.; Wang, X. Serum metabolomics as a novel diagnostic approach for disease: A systematic review. Anal. Bioanal. Chem. 2012, 404, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Araújo, R.; Bento, L.F.N.; Fonseca, T.A.H.; Von Rekowski, C.P.; da Cunha, B.R.; Calado, C.R.C. Infection biomarkers based on metabolomics. Metabolites 2022, 12, 92. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Cui, H.; Zhang, Y.; Xie, W.; Lin, Y.; Guo, Y.; Huang, T.; Xue, B.; Guo, W.; Huang, Z.; et al. Baicalin ameliorates multidrug-resistant Pseudomonas aeruginosa induced pulmonary inflammation in rat via arginine biosynthesis. Biomed. Pharmacother. 2023, 162, 114660. [Google Scholar] [CrossRef]
- Chen, M.-L.; Jiang, H.-y.; Zeng, J.; Huang, L.; Lv, C.-z. Caffeic acid attenuates tissue damage and inflammatory response in Klebsiella pneumonia by modulating ahr-src-stat3-il-10 signaling pathway. Environ. Toxicol. 2024, 39, 2254–2264. [Google Scholar] [CrossRef]
- Li, J.H.; Zou, P.C.; Xiao, R.Q.; Wang, Y.J. Indole-3-propionic acid alleviates dss-induced colitis in mice through macrophage glycolipid metabolism. Int. Immunopharmacol. 2025, 152, 114388. [Google Scholar] [CrossRef]
- Xiao, P.; Hu, Z.W.; Lang, J.H.; Pan, T.Y.; Mertens, R.T.; Zhang, H.L.; Guo, K.; Shen, M.L.; Cheng, H.Q.; Zhang, X.; et al. Mannose metabolism normalizes gut homeostasis by blocking the tnf-α-mediated proinflammatory circuit. Cell. Mol. Immunol. 2023, 20, 119–130. [Google Scholar] [CrossRef]
- Tang, Y.; Wang, Z.; Zhou, F.; Li, L.; Sun, C.; Li, L.; Tang, F.; Huang, D.; Li, Z.; Tan, Y.; et al. Benzoylpaeoniflorin alleviates ulcerative colitis by inhibiting ferroptosis through targeting phosphogluconic dehydrogenase. Phytomedicine 2025, 147, 157111. [Google Scholar] [CrossRef]
- Fu, Y.P.; Li, C.Y.; Peng, X.; Wangensteen, H.; Inngjerdingen, K.T.; Zou, Y.F. Pectic polysaccharides from Aconitum carmichaelii leaves protects against dss-induced ulcerative colitis in mice through modulations of metabolism and microbiota composition. Biomed. Pharmacother. 2022, 155, 113767. [Google Scholar] [CrossRef]
- Li, S.; Yang, S.Y.; Zhang, Y.; Huang, K.; Liang, T.; Chen, Y.; Guan, Y.J.; Shang, R.Z.; Guan, T.; Wu, J.; et al. Amino acid-balanced diets improved dss-induced colitis by alleviating inflammation and regulating gut microbiota. Eur. J. Nutr. 2022, 61, 3531–3543. [Google Scholar] [CrossRef]
- Barreau, F.; Hugot, J.P. Intestinal barrier dysfunction triggered by invasive bacteria. Curr. Opin. Microbiol. 2014, 17, 91–98. [Google Scholar] [CrossRef]
- McGuckin, M.A.; Linden, S.K.; Sutton, P.; Florin, T.H. Mucin dynamics and enteric pathogens. Nat. Rev. Microbiol. 2011, 9, 265–278. [Google Scholar] [CrossRef]
- Horowitz, A.; Chanez-Paredes, S.D.; Haest, X.; Turner, J.R. Paracellular permeability and tight junction regulation in gut health and disease. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 417–432. [Google Scholar] [CrossRef]
- Song, J.; Li, Q.; Everaert, N.; Liu, R.; Zheng, M.; Zhao, G.; Wen, J. Effects of inulin supplementation on intestinal barrier function and immunity in specific pathogen-free chickens with salmonella infection. J. Anim. Sci. 2020, 98, skz396. [Google Scholar] [CrossRef]
- Zou, Y.; Wang, J.; Wang, Y.; Peng, B.; Liu, J.; Zhang, B.; Lv, H.; Wang, S. Protection of galacto-oligosaccharide against E. coli o157 colonization through enhancing gut barrier function and modulating gut microbiota. Foods 2020, 9, 1710. [Google Scholar] [CrossRef]
- Wong, C.K.; Wang, C.B.; Ip, W.K.; Tian, Y.P.; Lam, C.W. Role of p38 mapk and nf-kb for chemokine release in coculture of human eosinophils and bronchial epithelial cells. Clin. Exp. Immunol. 2005, 139, 90–100. [Google Scholar] [CrossRef]
- Hobbie, S.; Chen, L.M.; Davis, R.J.; Galán, J.E. Involvement of mitogen-activated protein kinase pathways in the nuclear responses and cytokine production induced by salmonella typhimurium in cultured intestinal epithelial cells. J. Immunol. 1997, 159, 5550–5559. [Google Scholar] [CrossRef]
- Li, J.; Li, Q.; Wu, Q.; Gao, N.; Wang, Z.; Yang, Y.; Shan, A. Exopolysaccharides of Lactobacillus rhamnosus gg ameliorate salmonella typhimurium-induced intestinal inflammation via the tlr4/nf-κb/mapk pathway. J. Anim. Sci. Biotechnol. 2023, 14, 23. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, J.; Duan, H.; Li, R.; Peng, W.; Wu, C.J.J. Activation of nrf2/ho-1 signaling: An important molecular mechanism of herbal medicine in the treatment of atherosclerosis via the protection of vascular endothelial cells from oxidative stress. J. Adv. Res. 2021, 34, 43–63. [Google Scholar] [CrossRef]
- Bao, H.; Zhang, H.; Zhou, Y.; Zhu, S.; Pang, M.; Zhang, X.; Wang, Y.; Wang, J.; Olaniran, A.; Xiao, Y.; et al. Dysbiosis and intestinal inflammation caused by salmonella typhimurium in mice can be alleviated by preadministration of a lytic phage. Microbiol. Res. 2022, 260, 127020. [Google Scholar] [CrossRef]
- Zhang, L.; Gui, S.; Wang, J.; Chen, Q.; Zeng, J.; Liu, A.; Chen, Z.; Lu, X. Oral administration of green tea polyphenols (tp) improves ileal injury and intestinal flora disorder in mice with salmonella typhimurium infection via resisting inflammation, enhancing antioxidant action and preserving tight junction. J. Funct. Foods 2020, 64, 103654. [Google Scholar] [CrossRef]
- Wang, B.; Ye, X.; Zhou, Y.; Zhao, P.; Mao, Y. Glycyrrhizin attenuates salmonella typhimurium-induced tissue injury, inflammatory response, and intestinal dysbiosis in c57bl/6 Mice. Front. Vet. Sci. 2021, 8, 648698. [Google Scholar] [CrossRef]
- Singh, P.; Teal, T.K.; Marsh, T.L.; Tiedje, J.M.; Mosci, R.; Jernigan, K.; Zell, A.; Newton, D.W.; Salimnia, H.; Lephart, P.; et al. Intestinal microbial communities associated with acute enteric infections and disease recovery. Microbiome 2015, 3, 45. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The firmicutes/bacteroidetes ratio: A relevant marker of gut dysbiosis in obese patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Rizzatti, G.; Lopetuso, L.R.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A common factor in human diseases. BioMed Res. Int. 2017, 2017, 9351507. [Google Scholar] [CrossRef]
- Camelo-Castillo, A.J.; Mira, A.; Pico, A.; Nibali, L.; Henderson, B.; Donos, N.; Tomás, I. Subgingival microbiota in health compared to periodontitis and the influence of smoking. Front. Microbiol. 2015, 6, 119. [Google Scholar] [CrossRef]
- Lin, H.; An, Y.; Hao, F.; Wang, Y.; Tang, H. Correlations of fecal metabonomic and microbiomic changes induced by high-fat diet in the pre-obesity state. Sci. Rep. 2016, 6, 21618. [Google Scholar] [CrossRef]
- Huang, G.; Zheng, Y.; Zhang, N.; Huang, G.; Zhang, W.; Li, Q.; Ren, X. Desulfovibrio vulgaris caused gut inflammation and aggravated dss-induced colitis in c57bl/6 mice model. Gut Pathog. 2024, 16, 39. [Google Scholar] [CrossRef] [PubMed]
- Dou, X.; Gao, N.; Yan, D.; Shan, A. Sodium butyrate alleviates mouse colitis by regulating gut microbiota dysbiosis. Animals 2020, 10, 1154. [Google Scholar] [CrossRef] [PubMed]
- Xi, L.; Wen, X.; Jia, T.; Han, J.; Qin, X.; Zhang, Y.; Wang, Z. Comparative study of the gut microbiota in three captive rhinopithecus species. BMC Genom. 2023, 24, 398. [Google Scholar] [CrossRef] [PubMed]
- Osawa, M.; Handa, O.; Fukushima, S.; Matsumoto, H.; Umegaki, E.; Inoue, R.; Naito, Y.; Shiotani, A. Reduced abundance of butyric acid-producing bacteria in the ileal mucosa-associated microbiota of ulcerative colitis patients. J. Clin. Biochem. Nutr. 2023, 73, 77–83. [Google Scholar] [CrossRef]
- Yao, Y.; Cai, X.; Fei, W.; Ye, Y.; Zhao, M.; Zheng, C. The role of short-chain fatty acids in immunity, inflammation and metabolism. Crit. Rev. Food Sci. Nutr. 2022, 62, 1–12. [Google Scholar] [CrossRef]
- Song, J.; Li, Q.; Everaert, N.; Liu, R.; Zheng, M.; Zhao, G.; Wen, J. Dietary inulin supplementation modulates short-chain fatty acid levels and cecum microbiota composition and function in chickens infected with salmonella. Front. Microbiol. 2020, 11, 584380. [Google Scholar] [CrossRef]
- Aljumaah, M.R.; Alkhulaifi, M.M.; Abudabos, A.M.; Alabdullatifb, A.; El-Mubarak, A.H.; Al Suliman, A.R.; Stanley, D. Organic acid blend supplementation increases butyrate and acetate production in salmonella enterica serovar typhimurium challenged broilers. PLoS ONE 2020, 15, e0232831. [Google Scholar] [CrossRef]
- Zhan, Z.; Tang, H.; Zhang, Y.; Huang, X.; Xu, M. Potential of gut-derived short-chain fatty acids to control enteric pathogens. Front. Microbiol. 2022, 13, 976406. [Google Scholar] [CrossRef]
- Wang, L.L.; Hu, L.J.; Yan, S.; Jiang, T.; Fang, S.G.; Wang, G.; Zhao, J.X.; Zhang, H.; Chen, W. Effects of different oligosaccharides at various dosages on the composition of gut microbiota and short-chain fatty acids in mice with constipation. Food Funct. 2017, 8, 1966–1978. [Google Scholar] [CrossRef]
- Onrust, L.; Baeyen, S.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Effect of in feed administration of different butyrate formulations on salmonella enteritidis colonization and cecal microbiota in broilers. Vet. Res. 2020, 51, 56. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, C.C.; Li, Y.R.; Han, X.L.; Luo, X.G.; Chen, L.H.; Zhang, T.C.; Wang, N.; Wang, W.M. Alginate oligosaccharides ameliorate dss-induced colitis through modulation of ampk/nf-κb pathway and intestinal microbiota. Nutrients 2022, 14, 2864. [Google Scholar] [CrossRef] [PubMed]
- Bosco, M.C. Macrophage polarization: Reaching across the aisle. J. Allergy Clin. Immunol. 2019, 143, 1348–1350. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Ji, X.G.; Chang, K.L.; Yin, H.; Zhao, M.Y.; Zhao, L.M. The regulatory effect of chitooligosaccharides on islet inflammation in t2d individuals after islet cell transplantation: The mechanism behind Candida albicans abundance and macrophage polarization. Gut Microbes 2025, 17, 2442051. [Google Scholar] [CrossRef]
- Ye, M.; Fan, M.; Zhao, Y.; Wang, F.; Yang, X.; Yao, W.; Gao, X.; Yu, J.; Liu, W. Low molecular weight Astragalus membranaceus polysaccharides alleviates dextran sulfate sodium-induced colitis in mice. Carbohydr. Polym. 2025, 367, 124050. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Wang, L.; Xiao, Z.; Yang, J.; Lu, C.; Ren, X.; Song, S.; Jiang, J.; Ai, C. Low-Molecular-Weight Fucoidan from Undaria pinnatifida Mitigates Salmonella-Induced Injury Through Gut Microbiota and Immune Regulation. Foods 2026, 15, 2135. https://doi.org/10.3390/foods15122135
Wang L, Xiao Z, Yang J, Lu C, Ren X, Song S, Jiang J, Ai C. Low-Molecular-Weight Fucoidan from Undaria pinnatifida Mitigates Salmonella-Induced Injury Through Gut Microbiota and Immune Regulation. Foods. 2026; 15(12):2135. https://doi.org/10.3390/foods15122135
Chicago/Turabian StyleWang, Lu, Zhixiu Xiao, Jiaxin Yang, Chunyan Lu, Xiaomeng Ren, Shuang Song, Jinchi Jiang, and Chunqing Ai. 2026. "Low-Molecular-Weight Fucoidan from Undaria pinnatifida Mitigates Salmonella-Induced Injury Through Gut Microbiota and Immune Regulation" Foods 15, no. 12: 2135. https://doi.org/10.3390/foods15122135
APA StyleWang, L., Xiao, Z., Yang, J., Lu, C., Ren, X., Song, S., Jiang, J., & Ai, C. (2026). Low-Molecular-Weight Fucoidan from Undaria pinnatifida Mitigates Salmonella-Induced Injury Through Gut Microbiota and Immune Regulation. Foods, 15(12), 2135. https://doi.org/10.3390/foods15122135