
Investigating the Spatial Heterogeneity of Microbial Communities and Flavor Compounds During the First Fermentation Stage of Sauce-Flavor Baijiu

Abstract
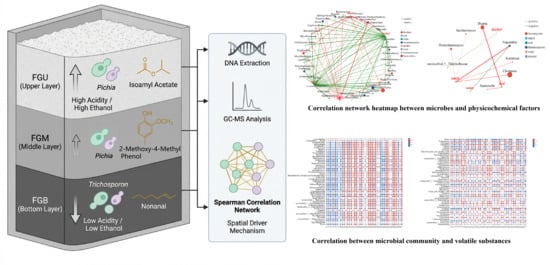
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Instruments and Equipment
2.3. Methods
2.3.1. Sample Collection
2.3.2. Determination of Physicochemical Indices of FG
2.3.3. Total DNA Extraction, PCR Amplification, and High-Throughput Sequencing of FG
2.3.4. Determination of Volatile Compounds in FG
2.4. Data Processing
3. Results
3.1. Changes in Physicochemical Parameters of Fermented Grains During the First Fermentation Cycle
3.2. Microbial Community Diversity During the First Fermentation Cycle in Baijiu Pits
3.2.1. Diversity Analysis of Bacterial and Fungal Communities in Fermented Grains
3.2.2. Analysis of Bacterial and Fungal Community Composition and Genera Divergence in Fermented Grains
3.2.3. Correlation Analysis and Functional Prediction of Microorganisms and Physicochemical Factors in Different Layers of Fermented Grains
3.3. Volatile Compounds in Fermented Grains of the First Round of Pit Fermentation During Liquor Brewing
3.4. Analysis of Microbial Community and Physicochemical Composition in Different Layers of the First-Round Pit Fermentation of Baijiu and Correlation Prediction of Their Impact on Volatile Compounds
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, X.; Du, B.; Liu, J.; Zhang, C.; Zhu, H.; Wang, K.; Sun, B.; Li, X. Exploring the significance of the 2nd and 4th round fermentations in the brewing process of Sauce-flavor Baijiu. Food Biosci. 2024, 59, 104114. [Google Scholar] [CrossRef]
- Wang, M.Y.; Yang, J.G.; Zhao, Q.S.; Zhang, K.Z.; Su, C. Research progress on flavor compounds and microorganisms of Maotai flavor baijiu. J. Food Sci. 2018, 84, 6–18. [Google Scholar] [CrossRef]
- Wang, H.; Huang, Y.; Huang, Y. Microbiome diversity and evolution in stacking fermentation during different rounds of Jiang-flavoured Baijiu brewing. LWT-Food Sci. Technol. 2021, 143, 111119. [Google Scholar] [CrossRef]
- Zhang, C.; Li, J.; Zeng, X.; Wang, Y.; Qin, H.; Song, P.; Hou, X.; Liu, S.; Ma, C.; Huang, Y.; et al. Comprehensive use of distillers’ grains derived from Chinese Baijiu: A review. J. Agric. Food Res. 2024, 18, 101439. [Google Scholar] [CrossRef]
- Hu, X.; Du, H.; Ren, C.; Xu, Y. Illuminating anaerobic microbial community and cooccurrence patterns across a quality gradient in Chinese liquor fermentation pit muds. Appl. Environ. Microbiol. 2016, 82, 2506–2515. [Google Scholar] [CrossRef]
- Jin, Y.; Li, D.; Ai, M.; Tang, Q.; Huang, J.; Ding, X.; Wu, C.; Zhou, R. Correlation between volatile profiles and microbial communities: A metabonomic approach to study Jiang-flavor liquor Daqu. Food Res. Int. 2019, 121, 422–432. [Google Scholar] [CrossRef]
- Kang, J.; Huang, X.; Li, R.; Zhang, Y.; Chen, X.; Han, B. Deciphering the core microbes and their interactions in spontaneous Baijiu fermentation: A comprehensive review. Food Res. Int. 2024, 188, 114497. [Google Scholar] [CrossRef]
- Chen, C.; Xiong, Y.; Xie, Y.; Zhang, H.; Jiang, K.; Pang, X.; Huang, M. Metabolic characteristics of Lactobacillus and interaction with yeast isolated from light-flavor Baijiu fermentation. Food Biosci. 2022, 50, 102102. [Google Scholar] [CrossRef]
- Xu, Y.; Wu, M.; Niu, J.; Lin, M.; Zhu, H.; Wang, K.; Li, X.; Sun, B. Characteristics and correlation of the microbial communities and flavor compounds during the first three rounds of fermentation in Chinese Sauce-flavor Baijiu. Foods 2023, 12, 207. [Google Scholar] [CrossRef]
- Guan, T.; Wu, X.; Hou, R.; Tian, L.; Huang, Q.; Zhao, F.; Liu, Y.; Jiao, S.; Xiang, S.; Zhang, J.; et al. Application of Clostridium butyricum, Rummeliibacillus suwonensis, and Issatchenkia orientalis for Nongxiangxing baijiu fermentation: Improves the microbial communities and flavor of upper fermented grain. Food Res. Int. 2023, 169, 112885. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wu, Y.; Zhu, H.; Wang, H.; Lu, H.; Zhang, C.; Li, X.; Xu, Y.; Li, W.; Wang, Y. Turning over fermented grains elevating heap temperature and driving microbial community succession during the heap fermentation of sauce-flavor Baijiu. LWT-Food Sci. Technol. 2022, 172, 114173. [Google Scholar] [CrossRef]
- Chen, Z.; Cao, Q.; Meng, T.; Shi, W.; Zhang, X.; Chai, L.; Shi, J.; Chen, F.; Wang, S.; Zhang, S.; et al. Yeast community in the first-round fermentation of Sauce-flavor Baijiu: Source, succession and metabolic function. Food Res. Int. 2025, 200, 115466. [Google Scholar] [CrossRef]
- T/CBJ 004-2018; The General Analysis Method of Solid-State Fermented Grains. China Alcoholic Drinks Association: Beijing, China, 2018.
- GB/T 601-2016; Chemical Reagent-Preparations of Reference Titration Solutions. State General Administration of the People’s Republic of China for Quality Supervision and Inspection and Quarantine, Standardization Administration of the People’s Republic of China: Beijing, China, 2016.
- GB/T 603-2023; Chemical Reagent-Preparations of Reagent Solutions for Use in Test Methods. State Administration for Market Regulation, Standardization Administration of the People’s Republic of China: Beijing, China, 2023.
- Liu, X.; Mu, Y.; Lv, X.; Chen, N.; Chen, L.; Wen, T.; Su, W. Analysis of fermentation characteristics in fermented grains across seven rounds of sauce-flavored Baijiu: Microbial communities structure, physicochemical parameters, volatile and non-volatile flavor compounds. Food Chem. X 2025, 25, 102228. [Google Scholar] [CrossRef]
- Lin, Y.; Yuan, D.; Zhang, S.; Wu, T. Differences in flavor characteristics of rounds of Sauce-based Baijiu based on GC-IMS, GC-MS, and electronic tongue. J. Food Compos. Anal. 2024, 135, 106696. [Google Scholar] [CrossRef]
- Hao, F.; Tan, Y.; Lv, X.; Chen, L.; Yang, F.; Wang, H.; Du, H.; Wang, L.; Xu, Y. Microbial community succession and its environment driving factors during initial fermentation of Maotai-flavor baijiu. Front. Microbiol. 2021, 12, 669201. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhang, H.; Du, H.; Yuan, C.; Xu, Y. Microorganisms and metabolic characteristics of temperature-dependent fermentation during Sauce-flavor Baijiu production. Food Biosci. 2025, 63, 105787. [Google Scholar] [CrossRef]
- Li, Y.; Qin, X.; Zeng, X.; She, M.; Chen, L.; Lu, J.; Han, Y.; Yang, F.; Tu, H.; Du, H.; et al. Comprehensive analysis of spatiotemporal heterogeneity reveals the effects of physicochemical and biological factors on temperature rise during the Moutai-flavor Baijiu stacking fermentation process. Chem. Eng. J. 2025, 508, 161049. [Google Scholar] [CrossRef]
- Fredlund, E.; Blank, L.M.; Schnürer, J.; Sauer, U.; Passoth, V. Oxygen- and glucose-dependent regulation of central carbon metabolism in Pichia anomala. Appl. Environ. Microbiol. 2004, 70, 5905–5911. [Google Scholar] [CrossRef] [PubMed]
- Basso, R.F.; Alcarde, A.R.; Portugal, C.B. Could non-Saccharomyces yeasts contribute on innovative brewing fermentations? Food Res. Int. 2016, 86, 112–120. [Google Scholar] [CrossRef]
- Chen, L.; Zheng, H.; Cheng, K.; Li, C.; Qin, X.; Wang, G.; Yang, F.; Du, H.; Wang, L.; Xu, Y. Deciphering the acidophilia and acid resistance in Acetilactobacillus jinshanensis dominating baijiu fermentation through multi-omics analysis. Food Microbiol. 2025, 125, 104655. [Google Scholar] [CrossRef]
- Fan, G.; Liu, P.; Chang, X.; Yin, H.; Cheng, L.; Teng, C.; Gong, Y.; Li, X. Isolation and identification of a high-yield ethyl caproate-producing yeast from Daqu and optimization of its fermentation. Front. Microbiol. 2021, 12, 663744. [Google Scholar] [CrossRef]
- Beckner, M.; Ivey, M.L.; Phister, T.G. Microbial contamination of fuel ethanol fermentations. Lett. Appl. Microbiol. 2011, 53, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, L.; Wang, H.; Yang, F.; Chen, L.; Hao, F.; Lv, X.; Du, H.; Xu, Y. Effects of initial temperature on microbial community succession rate and volatile flavors during Baijiu fermentation process. Food Res. Int. 2020, 141, 109887. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Zhong, H.; Zhao, D.; Du, H.; Xu, Y. Succession rate of microbial community causes flavor difference in strong-aroma Baijiu making process. Int. J. Food Microbiol. 2019, 311, 108350. [Google Scholar] [CrossRef]
- Wei, J.; Du, H.; Xu, Y. Revealing the key microorganisms producing higher alcohols and their assembly processes during Jiang-flavor Baijiu fermentation. Food Biosci. 2024, 61, 104569. [Google Scholar] [CrossRef]
- Yang, L.; Xian, C.; Li, P.; Wang, X.; Song, D.; Zhao, L.; Zhang, C. The spatio-temporal diversity and succession of microbial community and its environment driving factors during stacking fermentation of Maotai-flavor baijiu. Food Res. Int. 2023, 169, 112892. [Google Scholar] [CrossRef]
- Zhou, Y.; Hua, J. Lactic acid dynamics in Baijiu brewing: Microorganisms, roles, and control strategies. Fermentation 2025, 11, 431. [Google Scholar] [CrossRef]
- Liu, P.; Miao, L. Multiple batches of fermentation promote the formation of functional microbiota in Chinese Miscellaneous-flavor Baijiu fermentation. Front. Microbiol. 2020, 11, 75. [Google Scholar] [CrossRef]
- Pan, F.; Qiu, S.; Lv, Y.; Li, D. Exploring the controllability of the Baijiu fermentation process with microbiota orientation. Food Res. Int. 2023, 173, 113249. [Google Scholar] [CrossRef]
- Wu, Y.; Hao, F.; Lv, X.; Chen, B.; Yang, Y.; Zeng, X.; Yang, F.; Wang, H.; Wang, L. Diversity of Lactobacillus in Moutai-flavor liquor fermentation process. Food Biotechnol. 2020, 34, 212–227. [Google Scholar] [CrossRef]
- Randolph, N.K.; Her, J.; Mcaloney, C.A.; Wellman, M. Gastrointestinal colonization by Diutina (Candida) rugosain a 6-year-old Siberian Husky. Vet. Clin. Pathol. 2024, 53, 255–260. [Google Scholar] [CrossRef]
- Bian, M.; Fang, Y.; Yang, K.; Xia, Y.; Tang, H.; Feng, Y.; Xu, Q.; Zhang, S.; Ren, X.; Han, B. Comparative analysis of microbial communities and flavor compounds in fermented grains from different sorghum varieties used in Sichuan Xiaoqu liquor. LWT-Food Sci. Technol. 2025, 222, 117640. [Google Scholar] [CrossRef]
- Lei, Z.; Zhang, Z.; Huang, J.; Tang, Q.; Zhou, R. Dynamic patterns of high-temperature Daqu attributes in continuous production: Investigating the impact of Muqu attributes, production site, and process heterogeneity. Food Biosci. 2025, 68, 106417. [Google Scholar] [CrossRef]
- Gómez-Brandón, M.; Beesigamukama, D.; Probst, M.; Klammsteiner, T.; Su, J.; Zhu, Y.; Tanga, C.M. Microbial composition and bioremediation in frass fertilizers from insect-based agri-food waste valorization. J. Environ. Manag. 2025, 386, 125774. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Wu, Q.; Xu, Y. Filamentous fungal diversity and community structure associated with the solid state fermentation of Chinese Maotai-flavor liquor. Int. J. Food Microbiol. 2014, 179, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, T.; Hou, Z.; Dang, L.; Chen, M.; Bai, B.; Yang, Y.; Fan, S.; Bo, T. Germplasm resource mining of Fen-flavor Baijiu brewing micro-organisms and screening of important functional strains. Fermentation 2024, 10, 484. [Google Scholar] [CrossRef]
- Xu, S.; Hu, W.; Zhou, H.; Xu, B.; Wang, R.; Liu, W.; Zhang, H.; Xu, Q.; Liu, Y.; Li, X. Metatranscriptomic insights into the mechanism of ‘Multiple Qu’ utilization in Jian-flavor Baijiu fermentation. Food Res. Int. 2024, 200, 115463. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, J.; Yu, X.; Chen, S.; Xu, Y. Characterization of aroma differences in Jiangxiangxing Baijiu with varying ethanol concentrations: Emphasis on olfactory threshold changes of aroma compounds. Food Chem. 2025, 469, 142506. [Google Scholar] [CrossRef]
- Mou, Z.; Cai, X.; Liu, J.; Deng, R.; Liu, Z.; Fan, R.; Tang, J.; Luo, A. Elucidation of the key aroma compounds of floral and fruity aroma in sauce-flavored Baijiu by pervaporative membrane separation, GC-IMS, GC-MS, and aroma omission studies. Food Biosci. 2025, 66, 106270. [Google Scholar] [CrossRef]
- Wang, L.; Han, Y.; Zhang, X.; Gao, X.; Xu, Y.; Wu, Q.; Tang, K. Characterization of key aroma compounds of Zhuyeqing by aroma extract dilution analysis, quantitative measurements, aroma recombination, and omission studies. Foods 2025, 14, 344. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zou, M.; Tang, C.; Ao, H.; He, L.; Qiu, S.; Li, C. The insights into sour flavor and organic acids in alcoholic beverages. Food Chem. 2024, 460, 140676. [Google Scholar] [CrossRef]
- Zheng, Y.; Sun, B.; Zhao, M.; Zheng, F.; Huang, M.; Sun, J.; Sun, X.; Li, H. Characterization of the key odorants in Chinese Zhima Aroma-type Baijiu by gas chromatography–olfactometry, quantitative measurements, aroma recombination, and omission studies. J. Agric. Food. Chem. 2016, 64, 5367–5374. [Google Scholar] [CrossRef] [PubMed]
- Dharmsthiti, S.; Ammaranond, P. Purification and characterization of lipase from a raw-milk yeast (Trichosporon asteroides). Biotechnol. Appl. Biochem. 1997, 26, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulou, I.; Spanopoulos, A.; Matsakas, L. Single cell oil and ethanol production by the oleaginous yeast Trichosporon fermentans utilizing dried sweet sorghum stalks. Renew. Energy 2020, 146, 1609–1617. [Google Scholar] [CrossRef]
- Du, Y.K.; Xin, W.; Xia, Y.; Zhu, M.; Qin, J.L.; Pan, Z.F.; Wu, R.F.; Luo, G.R.; Wu, P.S.; Wu, Z.Y.; et al. Analysis of fermentation control factors on volatile compounds of primary microorganisms in Jiang-flavor Daqu. J. Food Biochem. 2022, 46, e14277. [Google Scholar] [CrossRef]
- Xu, Y.; Zhao, J.; Liu, X.; Zhang, C.; Zhao, Z.; Li, X.; Sun, B. Flavor mystery of Chinese traditional fermented baijiu: The great contribution of ester compounds. Food Chem. 2022, 369, 130920. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Bacteria | Fungi | ||||
|---|---|---|---|---|---|---|
| JCU | JCM | JCB | JCU | JCM | JCB | |
| ACE | 38.08 ± 10.51 a | 85.14 ± 35.28 a | 162.53 ± 27.95 b | 72.57 ± 12.72 a | 94.05 ± 13.42 a | 84.00 ± 9.53 a |
| Chao | 36.04 ± 9.48 a | 84.47 ± 34.74 a | 162.96 ± 29.81 b | 72.33 ± 12.50 a | 94.00 ± 13.45 a | 84.00 ± 9.54 a |
| Shannon | 1.52 ± 0.26 a | 1.83 ± 0.57 a | 2.87 ± 0.60 b | 2.07 ± 0.71 ab | 3.09 ± 0.25 b | 1.68 ± 0.52 a |
| Simpson | 0.35 ± 0.13 a | 0.33 ± 0.21 a | 0.27 ± 0.16 a | 0.25 ± 0.17 a | 0.19 ± 0.27 a | 0.36 ± 0.16 a |
| Coverage | 0.99 ± 0.00 a | 0.99 ± 0.00 a | 0.99 ± 0.00 a | 0.99 ± 0.00 a | 0.99 ± 0.00 a | 1.00 ± 0.00 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Wen, F.; Lu, F.; Zhao, Y.; You, J.; Li, S.; Yao, M.; Cao, X.; Lin, Y. Investigating the Spatial Heterogeneity of Microbial Communities and Flavor Compounds During the First Fermentation Stage of Sauce-Flavor Baijiu. Foods 2026, 15, 1849. https://doi.org/10.3390/foods15111849
Wen F, Lu F, Zhao Y, You J, Li S, Yao M, Cao X, Lin Y. Investigating the Spatial Heterogeneity of Microbial Communities and Flavor Compounds During the First Fermentation Stage of Sauce-Flavor Baijiu. Foods. 2026; 15(11):1849. https://doi.org/10.3390/foods15111849
Chicago/Turabian StyleWen, Fangling, Feifei Lu, Yu Zhao, Jiehua You, Shengfeng Li, Minna Yao, Xitao Cao, and Yonghui Lin. 2026. "Investigating the Spatial Heterogeneity of Microbial Communities and Flavor Compounds During the First Fermentation Stage of Sauce-Flavor Baijiu" Foods 15, no. 11: 1849. https://doi.org/10.3390/foods15111849
APA StyleWen, F., Lu, F., Zhao, Y., You, J., Li, S., Yao, M., Cao, X., & Lin, Y. (2026). Investigating the Spatial Heterogeneity of Microbial Communities and Flavor Compounds During the First Fermentation Stage of Sauce-Flavor Baijiu. Foods, 15(11), 1849. https://doi.org/10.3390/foods15111849