
Physiological Functions of the By-Products of Passion Fruit: Processing, Characteristics and Their Applications in Food Product Development

 ,
, 
Abstract
1. Introduction
2. Main Components of PFB
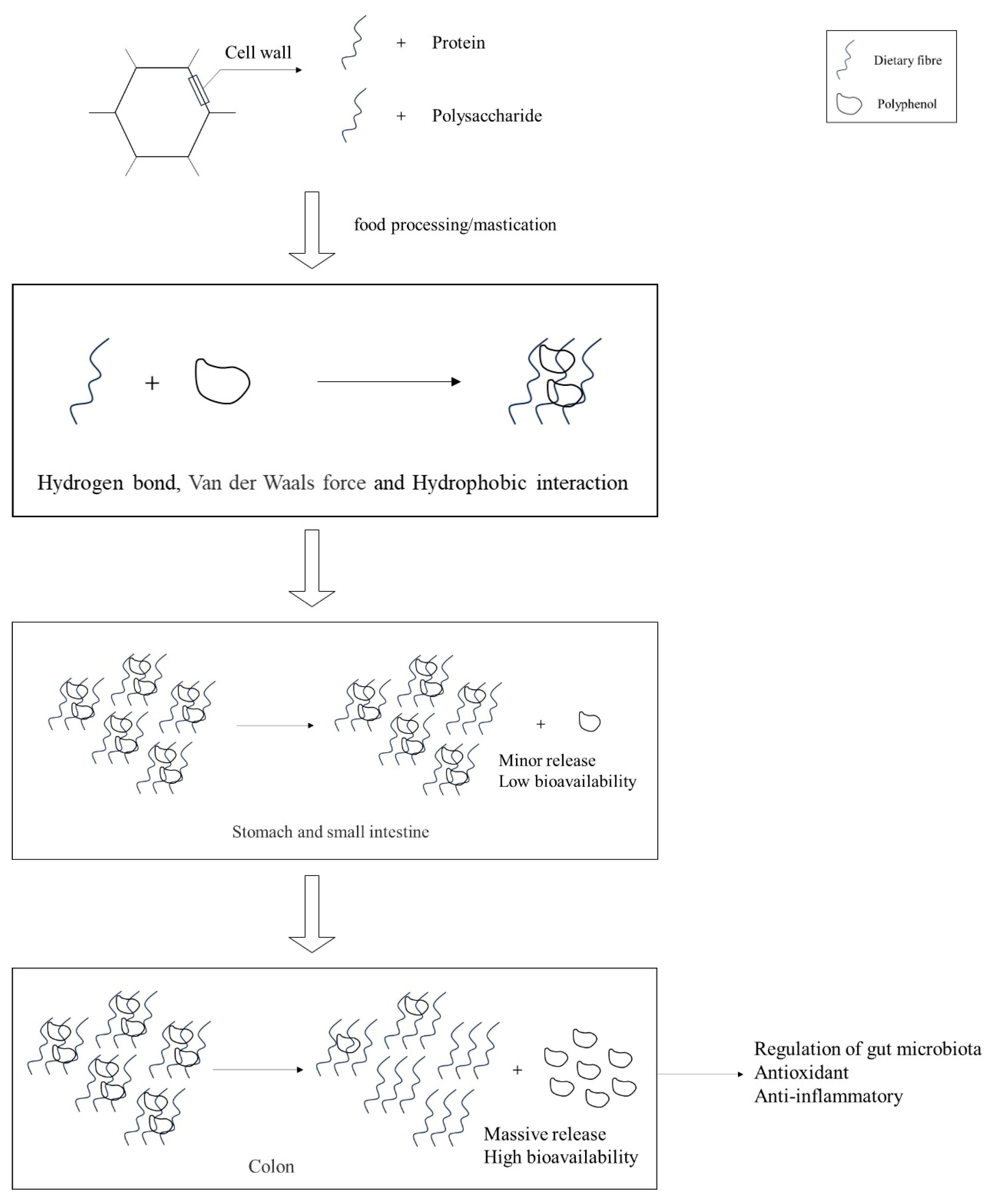
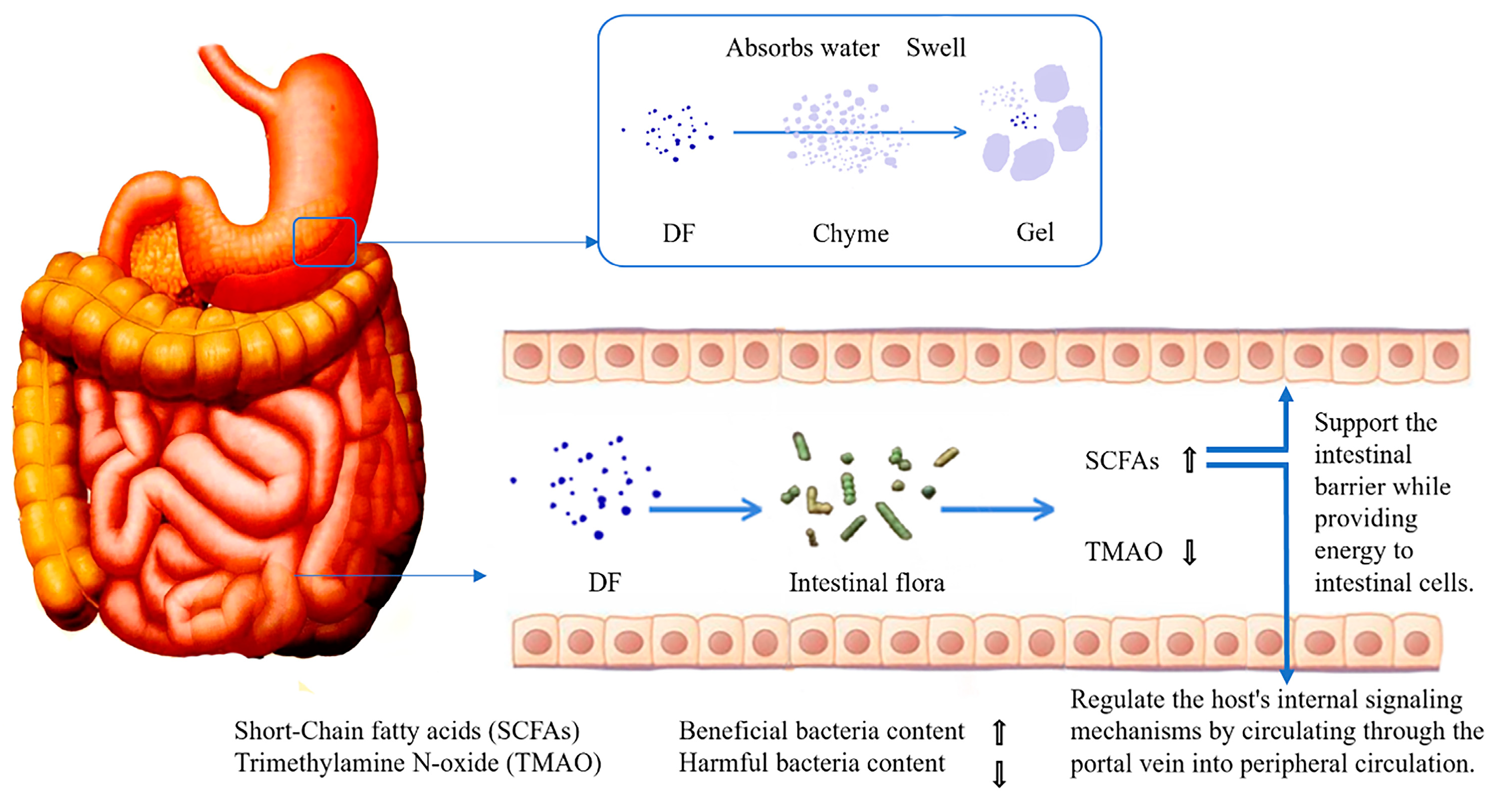
2.1. DF
2.2. Polyphenol
2.3. Other
3. Functional Characteristics of PFB
3.1. Lowers Blood Lipids
3.2. Hypoglycemia
3.3. Modulating Intestinal Flora
3.4. Slimming
4. Application of Passion Fruit DF in Food
4.1. Application in Dairy Products
4.2. Application in Noodle Products
4.3. Application in Meat Products
5. Conclusions and Prospect
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| PFB | Passion Fruit By-products |
| PFP | Passion Fruit Peel |
| PFS | Passion Fruit Seeds |
| DF | Dietary Fiber |
| LDL | Low-Density Lipoprotein |
| SDF | Soluble Dietary Fiber |
| IDF | Insoluble Dietary Fiber |
| TDF | Total Dietary Fiber |
| DPPH | 2,2-Diphenyl-1-Picrylhydrazyl |
| ABTS | Allion Bluetooth Test Suite |
| GAE | Gallic Acid Equivalents |
| WHC | Water Holding Capacity |
| OHC | Oil Holding Capacity |
| SR | Swelling Rate |
| PLE | Pressurized Liquid Extraction |
| UAE | Ultrasound-Assisted Extraction |
| MAE | Microwave-Assisted Extraction |
| ORAC | Oxygen Radical Absorbance Capacity |
| TE | Trolox Equivalent |
| SCFAs | Short-Chain Fatty Acids |
References
- Wyckhuys, K.A.; Korytkowski, C.; Martinez, J.; Herrera, B.; Rojas, M.; Ocampo, J. Species composition and seasonal occurrence of diptera associated with passionfruit crops in colombia. Crop Prot. 2012, 32, 90–98. [Google Scholar] [CrossRef]
- Zhu, X.-H.; Duan, Z.-H.; Yang, Y.-X.; Huang, X.-H.; Xu, C.-L.; Huang, Z.-Z. Development of passion fruit juice beverage. IOP Conf. Ser. Earth Environ. Sci. 2017, 100, 012080. [Google Scholar] [CrossRef]
- Oliveira, D.A.; Angonese, M.; Gomes, C.; Ferreira, S.R. Valorization of passion fruit (Passiflora edulis sp.) by-products: Sustainable recovery and biological activities. J. Supercrit. Fluids 2016, 111, 55–62. [Google Scholar] [CrossRef]
- Liu, Z.; Yang, T.; Chen, J.; Yang, C.; Niu, J.; Duan, X.; Ren, G.; Li, L. The impact of passion fruit peel powder on the physicochemical, sensory properties, and antioxidant activity of goat milk yoghurt. Int. J. Food Sci. Technol. 2024, 59, 4572–4582. [Google Scholar] [CrossRef]
- Leão, K.M.; Sampaio, K.L.; Pagani, A.A.; Da Silva, M.A.A. Odor potency, aroma profile and volatiles composition of cold pressed oil from industrial passion fruit residues. Ind. Crops Prod. 2014, 58, 280–286. [Google Scholar] [CrossRef]
- Pereira, Z.C.; dos Anjos Cruz, J.M.; Corrêa, R.F.; Sanches, E.A.; Campelo, P.H.; de Araújo Bezerra, J. Passion fruit (Passiflora spp.) pulp: A review on bioactive properties, health benefits and technological potential. Food Res. Int. 2023, 166, 112626. [Google Scholar] [CrossRef]
- Abbas, M.; Saeed, F.; Anjum, F.M.; Afzaal, M.; Tufail, T.; Bashir, M.S.; Ishtiaq, A.; Hussain, S.; Suleria, H.A.R. Natural polyphenols: An overview. Int. J. Food Prop. 2017, 20, 1689–1699. [Google Scholar] [CrossRef]
- Yang, Y.-y.; Ma, S.; Wang, X.-x.; Zheng, X.-l. Modification and application of dietary fiber in foods. J. Chem. 2017, 2017, 9340427. [Google Scholar] [CrossRef]
- Weyya, G.; Belay, A.; Tadesse, E. Passion fruit (Passiflora edulis Sims) by-products as a source of bioactive compounds for non-communicable disease prevention: Extraction methods and mechanisms of action: A systematic review. Front. Nutr. 2024, 11, 1340511. [Google Scholar] [CrossRef]
- Zibadi, S.; Farid, R.; Moriguchi, S.; Lu, Y.; Foo, L.Y.; Tehrani, P.M.; Ulreich, J.B.; Watson, R.R. Oral administration of purple passion fruit peel extract attenuates blood pressure in female spontaneously hypertensive rats and humans. Nutr. Res. 2007, 27, 408–416. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, X.; Yang, Q.; Li, L.; Cao, Y.; Zhong, R.; Miao, J. Extraction and characterization of passion fruit seed oil and investigation of its hypolipidemic activity. J. Agric. Food Res. 2025, 19, 101672. [Google Scholar] [CrossRef]
- Lupton, J.R.; Betteridge, V.A.; Pijls, L.T.J. Codex final definition of dietary fibre: Issues of implementation. Qual. Assur. Saf. Crops Foods 2009, 1, 206–212. [Google Scholar] [CrossRef]
- Feng, Z.; Dou, W.; Alaxi, S.; Niu, Y.; Yu, L. Modified soluble dietary fiber from black bean coats with its rheological and bile acid binding properties. Food Hydrocoll. 2017, 62, 94–101. [Google Scholar] [CrossRef]
- He, Y.; Wang, B.; Wen, L.; Wang, F.; Yu, H.; Chen, D.; Su, X.; Zhang, C. Effects of dietary fiber on human health. Food Sci. Hum. Wellness 2022, 11, 1–10. [Google Scholar] [CrossRef]
- Yang, C.F.; Teng, Z.; Wu, F.Y.; Liao, S.K.; Huang, K.L. Ultrasound-assisted enzymatic extraction of SDF on passion fruit seeds. Agric. Sci. Technol. 2020, 21, 28–32. [Google Scholar]
- Zhao, G.; Zhang, R.; Dong, L.; Huang, F.; Tang, X.; Wei, Z.; Zhang, M. Particle size of insoluble dietary fiber from rice bran affects its phenolic profile, bioaccessibility and functional properties. LWT 2018, 87, 450–456. [Google Scholar] [CrossRef]
- Sharma, V.; Devkota, L.; Kishore, N.; Dhital, S. Understanding the Interplay between Dietary Fiber, Polyphenols, and Digestive Enzymes. Food Hydrocoll. 2025, 166, 111310. [Google Scholar] [CrossRef]
- Wang, L.; Li, T.; Wu, C.; Fan, G.; Zhou, D.; Li, X. Unlocking the potential of plant polyphenols: Advances in extraction, antibacterial mechanisms, and future applications. Food Sci. Biotechnol. 2024, 34, 1235–1259. [Google Scholar] [CrossRef]
- Dos Reis, L.C.R.; Facco, E.M.P.; Salvador, M.; Flores, S.H.; de Oliveira Rios, A. Antioxidant potential and physicochemical characterization of yellow, purple and orange passion fruit. J. Food Sci. Technol. 2018, 55, 2679–2691. [Google Scholar] [CrossRef]
- Salgado, J.M.; Bombarde, T.A.D.; Mansi, D.N.; Piedade, S.M.D.S.; Meletti, L.M.M. Effects of different concentrations of passion fruit peel (Passiflora edulis) on the glicemic control in diabetic rat. Food Sci. Technol. 2010, 30, 784–789. [Google Scholar] [CrossRef]
- Duarte, Y.; Chaux, A.; Lopez, N.; Largo, E.; Ramírez, C.; Nuñez, H.; Simpson, R.; Vega, O. Effects of blanching and hot air drying conditions on the physicochemical and technological properties of yellow passion fruit (Passiflora edulis Var. Flavicarpa) by-products. J. Food Process Eng. 2017, 40, e12425. [Google Scholar] [CrossRef]
- Domínguez-Rodríguez, G.; García, M.C.; Plaza, M.; Marina, M.L. Revalorization of passiflora species peels as a sustainable source of antioxidant phenolic compounds. Sci. Total Environ. 2019, 696, 134030. [Google Scholar] [CrossRef]
- Hernández-Santos, B.; Vivar-Vera, M.D.L.Á.; Rodríguez-Miranda, J.; Herman-Lara, E.; Torruco-Uco, J.G.; Acevedo-Vendrell, O.; Martínez-Sánchez, C.E. Dietary fibre and antioxidant compounds in passion fruit (Passiflora edulis f. flavicarpa) peel and depectinised peel waste. Int. J. Food Sci. Technol. 2015, 50, 268–274. [Google Scholar] [CrossRef]
- Yapo, B.M.; Koffi, K.L. Dietary fiber components in yellow passion fruit rind—A potential fiber source. J. Agric. Food Chem. 2008, 56, 5880–5883. [Google Scholar] [CrossRef] [PubMed]
- Kawama, T. Constituent characteristics and functional properties of passion fruit seed extract. Life 2021, 12, 2–15. [Google Scholar] [CrossRef]
- Chutia, H.; Sharma, M.; Mahanta, D.C.L. Properties of dietary fibre from passion fruit seed obtained through individual and combined alkaline and ultrasonication extraction techniques. Waste Biomass Valorization 2024, 15, 2345–2359. [Google Scholar] [CrossRef]
- Singla, R.K.; Dubey, A.K.; Garg, A.; Sharma, R.K.; Fiorino, M.; Ameen, S.M.; Haddad, M.A.; Al-Hiary, M. Natural polyphenols: Chemical classification, definition of classes, subcategories, and structures. J. AOAC Int. 2019, 102, 1397–1400. [Google Scholar] [CrossRef]
- Tapiero, H.; Tew, K.; Ba, G.N.; Mathé, G. Polyphenols: Do they play a role in the prevention of human pathologies? Biomed. Pharmacother. 2002, 56, 200–207. [Google Scholar] [CrossRef]
- Montenegro, L.M.F.; Tapia, Q.P.; Vecino, X.; Reig, M.; Valderrama, C.; Granados, M.; Cortina, J.L.; Saurina, J. Fruit and vegetable processing wastes as natural sources of antioxidant-rich extracts: Evaluation of advanced extraction technologies by surface response methodology. J. Environ. Chem. Eng. 2021, 9, 105330. [Google Scholar] [CrossRef]
- Oliveira, A.M.B.; Viganó, J.; Sanches, V.L.; Rostagno, M.A.; Martínez, J. Extraction of potential bioactive compounds from industrial Tahiti lime (Citrus latifólia Tan.) by-product using pressurized liquids and ultrasound-assisted extraction. Food Res. Int. 2022, 157, 111381. [Google Scholar] [CrossRef]
- Kawasoe, H.; Wakamatsu, M.; Hamada, S.; Arata, Y.; Nagayoshi, K.; Uchida, R.; Yamashita, R.; Kishita, T.; Yamanouchi, H.; Minami, Y.; et al. Analysis of natural colourant extracted from the pericarp of passion fruit. LWT 2021, 136, 110412. [Google Scholar] [CrossRef]
- Ghada, B.; Pereira, E.; Pinela, J.; Prieto, M.A.; Pereira, C.; Calhelha, R.C.; Stojković, D.; Sokóvić, M.; Zaghdoudi, K.; Barros, L.; et al. Recovery of anthocyanins from passion fruit fpicarp for food colorants: Extraction process optimization and evaluation of bioactive properties. Molecules 2020, 25, 3203. [Google Scholar] [CrossRef] [PubMed]
- Malacrida, C.R.; Jorge, N. Yellow passion fruit seed oil (Passiflora edulis f. flavicarpa): Physical and chemical characteristics. Braz. Arch. Biol. Technol. 2012, 55, 127–134. [Google Scholar] [CrossRef]
- Cavalcanti-Mata, M.E.R.M.; Duarte, M.E.M.; Mendes, F.d.A.; Filho, M.T.L.; Almeida, R.D.; Cavalcanti, A.S.R.R.M.; Almeida, R.D. Residue from Passion Fruit Processing Industry: Application of Mathematical Drying Models for Seeds. Processes 2024, 12, 1752. [Google Scholar] [CrossRef]
- Marques, S.D.S.F.; Libonati, R.M.F.; Sabaa-Srur, A.U.O.; Luo, R.; Shejwalkar, P.; Hara, K.; Dobbs, T.; Smith, R.E. Evaluation of the effects of passion fruit peel flour (Passiflora edulis fo. flavicarpa) on metabolic canges in HIV patients with lipodystrophy syndrome secondary to antiretroviral therapy. Rev. Bras. Farmacogn. 2016, 26, 420–426. [Google Scholar] [CrossRef]
- Barbalho, S.M.; Damasceno, D.C.; Spada, A.P.M.; Lima, I.E.D.R.; Araújo, A.C.; Guiguer, E.L.; Martuchi, K.A.; Oshiiwa, M.; Mendes, C.G. Effects of Passiflora edulis on the metabolic profile of diabetic wistar rat offspring. J. Med. Food 2011, 14, 1490–1495. [Google Scholar] [CrossRef]
- Ramos, A.T.; Cunha, M.A.L.; Sabaa-Srur, A.U.; Pires, V.C.F.; Cardoso, A.A.; Diniz, M.D.F.; Medeiros, C.C.M. Uso de Passiflora edulis f. flavicarpa na redução do colesterol. Rev. Bras. Farmacogn. 2007, 17, 592–597. [Google Scholar] [CrossRef]
- Ishihata, A.; Maruki-Uchida, H.; Gotoh, N.; Kanno, S.; Aso, Y.; Togashi, S.; Sai, M.; Ito, T.; Katano, Y. Vascular and hepato protective effects of passion fruit seed extract containing piceatannol in chronic high-fat diet fed rats. Food Funct. 2016, 7, 4075–4081. [Google Scholar] [CrossRef]
- Chau, C.F.; Huang, Y.L. Effects of the insoluble fiber derived from Passiflora edulis seed on plasma and hepatic lipids and fecal output. Mol. Nutr. Food Res. 2010, 49, 786–790. [Google Scholar] [CrossRef]
- Chau, C.F.; Huang, Y.L. Characterization of passion fruit seed fibres—A potential fibre source. Food Chem. 2004, 85, 189–194. [Google Scholar] [CrossRef]
- Corrêa, E.; Medina, L.; Barros-Monteiro, J.; Valle, N.; Sales, R.; Magalães, A.; Souza, F.; Carvalho, T.; Lemos, J.; Lira, E.; et al. The intake of fiber mesocarp passionfruit (Passiflora edulis) lowers levels of triglyceride and cholesterol decreasing principally insulin and leptin. J. Aging Res. Clin. Pract. 2014, 3, 31–35. [Google Scholar] [PubMed]
- Li, Y.T. Compositions and Bioactivities of Extractable and Non-Extractable Polyphenols in Pyracantha Fortuneana Fruit and Passion Fruit Pomace. Master’s Thesis, Huazhong University of Science & Technology, Wuhan, China, 2019. [Google Scholar]
- Chong, F.T.; Huang, Z.Z.; Teng, J.W.; Wei, B.Y.; Huang, L.; Xia, N. Antioxidant Effect of Passion Fruit Peel and Its Effect on Inhibiting Glucose Absorption in Vitro and Regulating Intestinal Microflora Structure in Hyperglycemic Rats. Food Sci. 2021, 42, 193–200. [Google Scholar]
- Casarotti, S.N.; Borgonovi, T.F.; de Mello Tieghi, T.; Sivieri, K.; Penna, A.L.B. Probiotic low-fat fermented goat milk with passion fruit by-product: In vitro effect on obese individuals’ microbiota and on metabolites production. Food Res. Int. 2020, 136, 109453. [Google Scholar] [CrossRef]
- da Silva, J.K.; Cazarin, C.B.B.; Junior, S.B.; Augusto, F.; Junior, M.R.M. Passion fruit (Passiflora edulis) peel increases colonic production of short-chain fatty acids in Wistar rats. LWT-Food Sci. Technol. 2014, 59, 1252–1257. [Google Scholar] [CrossRef]
- Abboud, K.Y.; da Luz, B.B.; Dallazen, J.L.; de Paula Werner, M.F.; Cazarin, C.B.B.; Junior, M.R.M.; Iacomini, M.; Cordeiro, L.M. Gastroprotective effect of soluble dietary fibres from yellow passion fruit (Passiflora edulis f. flavicarpa) peel against ethanol-induced ulcer in rats. J. Funct. Foods 2019, 54, 552–558. [Google Scholar] [CrossRef]
- Lucas-González, R.; Capanoglu, E.; Pateiro, M.; Khaneghah, A.M.; Hano, C.; Lorenzo, J.M. Current trends in Passiflora genus research: Obesity and fermented foods systematic review. Trends Food Sci. Technol. 2022, 127, 143–155. [Google Scholar] [CrossRef]
- Vuolo, M.M.; Lima, G.C.; Batista, Â.G.; Carazin, C.B.B.; Cintra, D.E.; Prado, M.A.; Júnior, M.R.M. Passion fruit peel intake decreases inflammatory response and reverts lipid peroxidation and adiposity in diet-induced obese rats. Nutr. Res. 2020, 76, 106–117. [Google Scholar] [CrossRef]
- Kitada, M.; Ogura, Y.; Maruki-Uchida, H.; Sai, M.; Suzuki, T.; Kanasaki, K.; Hara, Y.; Seto, H.; Kuroshima, Y.; Monno, I.; et al. The Effect of Piceatannol from Passion Fruit (Passiflora edulis) Seeds on Metabolic Health in Humans. Nutrients 2017, 9, 1142. [Google Scholar] [CrossRef]
- Wang, N.; Mustafa, R.; Zuber, V.; Rodgers, A.; Dehghan, A. Association between systolic blood pressure and low-density lipoprotein cholesterol with coronary heart disease according to age. PLoS ONE 2023, 18, e0295004. [Google Scholar] [CrossRef]
- Bozzetto, L.; Costabile, G.; Della Pepa, G.; Ciciola, P.; Vetrani, C.; Vitale, M.; Rivellese, A.A.; Annuzzi, G. Dietary fibre as a unifying remedy for the whole spectrum of obesity-associated cardiovascular risk. Nutrients 2018, 10, 943. [Google Scholar] [CrossRef]
- Anderson, J.W.; Baird, P.; Davis, R.H., Jr.; Ferreri, S.; Knudtson, M.; Koraym, A.; Waters, V.; Williams, C.L. Health benefits of dietary fiber. Nutr. Rev. 2009, 67, 188–205. [Google Scholar] [CrossRef] [PubMed]
- Salgado, M.J.G.; Ramos, M.S.; Assis, D.J.; Otero, D.M.; Oliveira, R.L.; Ribeiro, C.V.D.M.; Costa, M.P.; Oliveira, C.A.A. Impact of fiber-rich donkey milk yogurt on apparent viscosity and sensory acceptance. LWT 2021, 145, 111494. [Google Scholar] [CrossRef]
- Pathak, P.; Xie, C.; Nichols, R.G.; Ferrell, J.M.; Boehme, S.; Krausz, K.W.; Patterson, A.D.; Gonzalez, F.J.; Chiang, J.Y.L. Intestine farnesoid X receptor agonist and the gut microbiota activate G-protein bile acid receptor-1 signaling to improve metabolism. Hepatology 2018, 68, 1574–1588. [Google Scholar] [CrossRef] [PubMed]
- Adam, A.; Leuillet, M.; Levrat-Verny, M.-A.; Lopez, H.W.; Demigné, C.; Rémésy, C. Whole wheat and triticale flours with differing viscosities stimulate cecal fermentations and lower plasma and hepatic lipids in rats. J. Nutr. 2001, 131, 1770–1776. [Google Scholar] [CrossRef]
- Seal, C.J.; Mathers, J.C. Comparative gastrointestinal and plasma cholesterol responses of rats fed on cholesterol-free diets supplemented with guar gum and sodium alginate. Br. J. Nutr. 2001, 85, 317–324. [Google Scholar] [CrossRef]
- Park, S.; Jung, S.; Yoon, H. The role of nutritional status in the relationship between diabetes and health-related quality of life. Nutr. Res. Pract. 2022, 16, 505–516. [Google Scholar] [CrossRef]
- Anderson, J.W. Dietary fiber and associated phytochemicals in prevention and reversal of diabetes. In Nutraceuticals, Glycemic Health and Type 2 Diabetes; Wiley: Hoboken, NJ, USA, 2008; Volume 7, pp. 97–125. [Google Scholar]
- Lindström, J.; Peltonen, M.; Eriksson, J.G.; Louheranta, A.; Fogelholm, M.; Uusitupa, M.; Tuomilehto, J. High-fibre, low-fat diet predicts long-term weight loss and decreased type 2 diabetes risk: The finnish diabetes prevention study. Diabetologia 2006, 49, 912–920. [Google Scholar] [CrossRef]
- Byrne, C.S.; Chambers, E.S.; Morrison, D.J.; Frost, G. The role of short chain fatty acids in appetite regulation and energy homeostasis. Int. J. Obes. 2015, 39, 1331–1338. [Google Scholar] [CrossRef]
- Psichas, A.; Sleeth, M.L.; Murphy, K.G.; Brooks, L.; Bewick, G.A.; Hanyaloglu, A.C.; Ghatei, M.A.; Bloom, S.R.; Frost, G. The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents. Int. J. Obes. 2015, 39, 424–429. [Google Scholar] [CrossRef]
- Liu, J.; Dai, Y.; Yang, W.; Chen, Z.-Y. Role of mushroom polysaccharides in modulation of GI homeostasis and protection of GI barrier. J. Agric. Food Chem. 2025, 73, 6416–6441. [Google Scholar] [CrossRef]
- Hashemi, Z.; Fouhse, J.; Im, H.S.; Chan, C.B.; Willing, B.P. Dietary pea fiber supplementation improves glycemia and induces changes in the composition of gut microbiota, serum short chain fatty acid profile and expression of mucins in glucose intolerant rats. Nutrients 2017, 9, 1236. [Google Scholar] [CrossRef] [PubMed]
- Gtarek, P.; Kauna, C.J. Trimethylamine N-oxide (TMAO) in human health. Excli J. 2021, 20, 301–319. [Google Scholar]
- Rose, D.J.; DeMeo, M.T.; Keshavarzian, A.; Hamaker, B.R. Influence of dietary fiber on inflammatory bowel disease and colon cancer: Importance of fermentation pattern. Nutr. Rev. 2007, 65, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Inan, M.S.; Rasoulpour, R.J.; Yin, L.; Hubbard, A.K.; Rosenberg, D.W.; Giardina, C. The luminal short-chain fatty acid butyrate modulates NF-κB activity in a human colonic epithelial cell line. Gastroenterology 2000, 118, 724–734. [Google Scholar] [CrossRef]
- Cesare, M.D.; Bentham, J.; Stevens, G.A.; Zhou, B.; Lu, Y.; Bixby, H.; Cowan, M.J.; Riley, L.M.; Hajifathalia, K.; Fortunato, L.; et al. Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19.2 million participants. Lancet 2016, 387, 1377–1396. [Google Scholar]
- Hammadi, E.A. Effects of dietary fiber intake on body weight and waist circumference. Arab J. Nutr. Exerc. 2016, 1, 77–84. [Google Scholar] [CrossRef]
- Espírito-Santo, A.P.D.; Lagazzo, A.; Sousa, A.L.O.P.D.; Perego, P.; Converti, A.; Oliveira, M.N. Rheology, spontaneous whey separation, microstructure and sensorial characteristics of probiotic yoghurts enriched with passion fruit fiber. Food Res. Int. 2013, 50, 224–231. [Google Scholar] [CrossRef]
- do Espírito Santo, A.P.; Perego, P.; Converti, A.; Oliveira, M.N. Influence of milk type and addition of passion fruit peel powder on fermentation kinetics, texture profile and bacterial viability in probiotic yoghurts. LWT 2012, 47, 393–399. [Google Scholar] [CrossRef]
- Rotta, E.M.; Giroux, H.J.; Lamothe, S.; Bélanger, D.; Sabik, H.; Visentainer, J.V.; Britten, M. Use of passion fruit seed extract (Passiflora edulis Sims) to prevent lipid oxidation in dairy beverages during storage and simulated digestion—ScienceDirect. LWT 2020, 123, 109088. [Google Scholar] [CrossRef]
- de Oliveira, P.V.C.; Lima, T.L.S.; de Oliveira, P.V.C.; Pereira, A.S.; de Carvalho Lima, M.H.; Júnior, E.M.Z.; da Silva Luz, K.S.; Pessoa, V.G. Frozen zero lactose com aproveitamento integral do maracujá (Passiflora edulis): Desenvolvimento de produto e elaboração de informação nutricional. Res. Soc. Dev. 2022, 11, e1611931616. [Google Scholar] [CrossRef]
- Costa, C.F.; Fusieger, A.; Andretta, M.; Camargo, A.C.; Carvalho, A.F.; Menezes, D.R.; Nero, L.A. Short communication: Potential use of passion fruit (Passiflora cincinnata) as a biopreservative in the production of coalho cheese, a traditional brazilian cheese. J. Dairy Sci. 2020, 103, 3082–3087. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, T.H.S.; Bolanho, B.C.; Montanuci, F.D.; Ruiz, S.P. Physicochemical and sensory characterization of gluten-free fresh pasta with addition of passion fruit peel flour. Ciência Rural 2018, 48, e20180508. [Google Scholar] [CrossRef]
- Ning, X.; Zhou, Y.; Wang, Z.; Zheng, X.; Pan, X.; Chen, Z.; Liu, Q.; Du, W.; Cao, X.; Wang, L. Evaluation of passion fruit mesocarp flour on the paste, dough, and quality characteristics of dried noodles. Food Sci. Nutr. 2022, 10, 1657–1666. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.V.; Milani, M.S.; Ries, E.F. Production optimization of passion fruit peel flour and its incorporation into dietary food. Food Sci. Technol. Int. 2020, 26, 132–139. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhou, M.; Mi, R.Y.; Guo, H.N. Extraction of Pectin from Passion Fruit Peel and Its Effect on the Dough and Bread Qualities. Food Res. Dev. 2020, 41, 102–107. [Google Scholar]
- de Azevedo Maia Junior, J.; da Costa Henry, F.; della Lucia, S.M.; Santos Junior, A.C.; de Oliveira Cabral, N.; Quirini, C.R.; Valle, F.R.A.F. Sensory attributes and Lipid oxidation of smoked lamb sausage formulated with passion fruit meal, potassium chloride and calcium chloride. Food Sci. Technol. 2019, 40, 423–429. [Google Scholar] [CrossRef]
- Hang, Y.Y.; Yu, S.C.; Shang, W.H.; Qi, D. Effect of Passion Fruit Peel Powder on the Gelation Characteristics of Non-rinsed Tilapia Surimi. Sci. Technol. Food Ind. 2022, 43, 92–97. [Google Scholar]
- López-Vargas, J.H.; Fernández-López, J.; Pérez-Álvarez, J.Á.; Viuda-Martos, M. Quality characteristics of pork burger added with albedo-fiber powder obtained from yellow passion fruit (Passiflora edulis var. flavicarpa) co-products. Meat Sci. 2014, 97, 270–276. [Google Scholar] [CrossRef]
- Zanetti, L.H.; Granero, L.; Da Luz, P.A.; Poletto, M.; Lala, B.; Delbem, N.L.C.; Sobral, N.C.; Brito, E.P.; Denadai, J.C.; Andrighetto, C.; et al. Chemical, physical and oxidative characteristics of broilers meat supplemented with passion fruit seed oil. Int. J. Innov. Educ. Res. 2021, 9, 69–83. [Google Scholar] [CrossRef]
- de Moura, F.A.; Macagnan, F.T.; Dos Santos, L.R.; Bizzani, M.; de Oliveira Petkowicz, C.L.; Da Silva, L.P. Characterization and physicochemical properties of pectins extracted from agroindustrial by-products. J. Food Sci. Technol. 2017, 54, 3111–3117. [Google Scholar] [CrossRef]
- Santos, E.; Andrade, R.; Gouveia, E. Utilization of the pectin and pulp of the passion fruit from Caatinga as probiotic food carriers. Food Biosci. 2017, 20, 56–61. [Google Scholar] [CrossRef]
- Ying, L.M. Passion Fruit Infused Gluten Free Noodle: Optimization of Ingredient Levels Using Response Surface Methodology. Bachelor’s Thesis, Universiti Tunku Abdul Rahman, Kampar Perak, Malaysia, 2016. [Google Scholar]
- Kim, H.J.; Paik, H.D. Functionality and application of dietary fiber in meat products. Food Sci. Anim. Resour. 2012, 32, 695–705. [Google Scholar] [CrossRef]
- Filho, E.G.A. Chemometrics applied to physical, physicochemical and sensorial attributes of chicken hamburgers blended with green banana and passion fruit epicarp biomasses. Int. J. Gastron. Food Sci. 2021, 24, 100337. [Google Scholar]
- Assunção, A.S.D.A.; Martins, R.A.; Vieira, J.C.S.; Rocha, L.C.; Krenchinski, F.K.D.L.; Buzalaf, M.A.R.; Sartori, J.R.; Padilha, P.D.M. Shotgun proteomics reveals changes in the pectoralis major muscle of broilers supplemented with passion fruit seed oil under cyclic heat stress conditions. Food Res. Int. 2023, 167, 112731. [Google Scholar] [CrossRef]
- Cao, Q.; Teng, J.; Wei, B.; Huang, L.; Xia, N. Phenolic compounds, bioactivity, and bioaccessibility of ethanol extracts from passion fruit peel based on simulated gastrointestinal digestion. Food Chem. 2021, 356, 129682. [Google Scholar] [CrossRef]
- Infante-Neta, A.A.; de Carvalho, Á.A.O.; D’Almeida, A.P.; Gonçalves, L.R.B.; de Albuquerque, T.L. Xylitol production from passion fruit peel hydrolysate: Optimization of hydrolysis and fermentation processes. Bioresour. Technol. 2024, 414, 131628. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Color | Research Object | TDF (SDF/IDF) | Phenolics | Other Active Ingredients | 2,2-Diphenyl-1-picrylhydrazyl (DPPH) | Allion Bluetooth Test Suite (ABTS) | Function Characteristics | Reference |
|---|---|---|---|---|---|---|---|---|
| yellow | Brazil-PFP | 61.16 ± 1.02% (Pectin: 37.67 ± 0.97%) | 1061.87 ± 25.00 mg/100 g | Quercetin: 760.00 ± 32.07 mg/100 g | 1.69 ± 0.03 g/100 mL | 2.22 ± 0.01 g/100 mL | — | [19] |
| Brazil-PFP | (3.49%/54.27%) | 4.2 mg/g | — | — | — | — | [20] | |
| Colombia-PFP | 63.40 ± 0.10% | 64.94 ± 0.27 mg Gallic acid equivalents (GAE)/g | — | — | — | Water holding capacity (WHC): 11.48 ± 2.54 mL/g Oil holding capacity (OHC): 5.07 ± 0.63 mL/g Swelling rate (SR): 14.72 ± 2.22 mL/g | [21] | |
| Colombia-PFP | — | 30.19 ± 3.00 mg GAE/g | — | 10.56 ± 0.80 μg/mL | — | — | [22] | |
| Colombia-PFP | — | 8.34 ± 0.83 mg GAE/g | — | 718.91 ± 40.55 μg/mL | — | — | [22] | |
| Mexico-PFP | 57.93 ± 2.72% (11.75 ± 1.21%/46.18 ± 1.21%) | 482.56 mg GAE/100 g | — | — | — | — | [23] | |
| Ecuador-PFP | — | 24.96 ± 2.00 mg GAE/g | — | 32.93 ± 2.88 μg/mL | — | — | [22] | |
| Côte d’Ivoire-PFP | 81.9 ± 1.40% (17.90 ± 0.40%/62.40 ± 0.70%) | — | — | — | — | WHC: 4.10 ± 0.10 g/g OHC: 5.20 ± 0.1 g/g | [24] | |
| Brazil-PFS | 65.60 ± 0.52% | 346.69 ± 6.58 mg/100 g | Anthocyanin: 4598.70 ± 119.73 μg/100 g Kaempferol: 375.32 ± 13.50 mg/100 g | 1.18 ± 0.03 g/100 mL | 3.84 ± 0.08 g/100 mL | — | [19] | |
| Brazil-PFS | 65.60 ± 0.52% | — | — | — | — | — | [25] | |
| India-PFS | 55.8 ± 2.1 (3.6 ± 0.6/52.2 ± 1.1)% | — | — | — | — | WHC: 2.9 ± 0.06 g/g OHC: 4.1 ± 0.05 g/g SR: 15.7 ± 0.08 mL/g | [26] | |
| purple | Brazil-PFP | 61.68 ± 1.31% (Pectin: 32.85 ± 1.20%) | 1570.80 ± 26.76 mg/100 g | Anthocyanin: 103686.48 ± 542.11 μg/100 g Kaempferol: 74.70 ± 1.44 mg/100 g | 6.98 ± 0.20 g/100 mL | 9.37 ± 0.05 g/100 mL | — | [19] |
| Colombia-PFP | — | 5.08 ± 0.48 mg GAE/g | — | 298.57 ± 18.31 μg/mL | — | — | [22] | |
| Brazil-PFS | 55.06 ± 0.35% | 325.69 ± 1.18 mg/100 g | Anthocyanin: 8232.41 ± 6.54 μg/100 g | 6.30 ± 0.08 g/100 mL | 4.76 ± 0.03 g/100 mL | — | [19] | |
| Brazil-PFS | 55.06 ± 0.35% | — | — | — | — | — | [25] | |
| orange | Brazil-PFP | 62.14 ± 2.62% (Pectin: 21.55 ± 0.55%) | 2584.91 ± 96.67 mg/100 g | Quercetin: 800.13 ± 24.18 mg/100 g Kaempferol: 229 ± 8.90 mg/100 g | 2.45 ± 0.03 g/100 mL | 2.95 ± 0.02 g/100 mL | — | [19] |
| Brazil-PFS | 51.47 ± 0.60% | 429.33 ± 0.19 mg/100 g | Anthocyanin: 293.36 ± 6.75 μg/100 g Quercetin: 120.41 ± 2.82 mg/100 g | 2.68 ± 0.03 g/100 mL | 3.87 ± 0.00 g/100 mL | — | [19] |
| Physiological Function | Research Object | Research Object | Potential Prevention Mechanisms | References |
|---|---|---|---|---|
| Lowers Blood Lipids | PFP | Human clinical trial, Animal test | The levels of triglycerides, LDL-cholesterol, and total cholesterol decreased, while the level of high-density lipoprotein cholesterol increased. | [35,36,37] |
| PFS-Ethanol extract | Animal test | Reduce the levels of triglycerides and cholesterol in rat serum. | [38] | |
| PFS-IDF | Animal test | Impede enterohepatic circulation; enhance bile acid excretion. | [39] | |
| Hypoglycemia | PFS-IDF | In vitro testing | Absorption of glucose and inhibition of amylase activity. | [40] |
| PFP-DF | Animal test | Reduce triglycerides and LDL-cholesterol, and reduce insulin or leptin levels. | [41] | |
| PFP | Animal test | Stimulate hepatic glycogen synthesis; enhance insulin sensitivity in adipose tissue. | [20] | |
| PFP-Polyphenol | In vitro testing | Inhibition of α-Glucosidase and α-Amylase Activities. | [42] | |
| Modulating Intestinal Flora | PFP-SDF | Animal test | Enrich beneficial bacteria and inhibit pathogenic bacteria. | [43] |
| PFB (PFP, PFS, Pomace) goat milk yogurt | In vitro testing | [44] | ||
| PFP | Animal test, In vitro testing | Enhance the content of short-chain fatty acids (SCFAs), exhibiting anti-inflammatory activity. | [45] | |
| PFP-SDF | Animal test | Reduced ethanol-induced gastric ulcer lesions. | [46] | |
| Slimming | PFP, PFS | Animal test | Prevent accumulation of body fat and liver damage. | [47] |
| PFP | Animal test | Improve the antioxidant defense capability of rat liver and epididymal adipose tissue; improve the inflammatory state and reduce body fat. | [48] | |
| PFS | Human clinical trial | significantly improved blood pressure and heart rate while enhancing insulin sensitivity in obese male subjects | [49] |
| Product Form | Additives | Advantages | Disadvantages | Conclusions | Reference | |
|---|---|---|---|---|---|---|
| Dairy Products | Milk yogurt | PFP | Shorten fermentation time, improve textural properties, increase the content of lactic acid bacteria, and enhance nutritional value. | Sensory evaluation has been marginally decreased | The yogurt with a 1% PFP addition exhibited the optimal characteristics. | [69] |
| Milk yogurt | PFP | Skim milk is more suitable than whole milk for the development of PFP yogurt. Physiologically active modifiers | [70] | |||
| Donkey milk yogurt | PFP-DF Apple peel-DF Inulin | Improved curd quality and enhanced nutritional value | Sensory evaluation has been marginally decreased | PFP-DF is more suitable for the development of DF yogurt compared with apple peel-DF and inulin. | [53] | |
| Milk-based compound beverage | PFS-polyphenol | Increase antioxidant activity and reduce lipid peroxidation. | — | PFS-polyphenol extract can prevent lipid oxidation in dairy beverages during storage and digestion. | [71] | |
| Lactose-free ice cream | Passion fruit pulp and PFP-pectin | Improved odor and enhanced nutritional value | — | The addition of whole passion fruit can develop products with lower processing degrees, no food additives, and higher micronutrient content. | [72] | |
| cheese | PFP | Inhibition of harmful bacteria growth, enhancement of nutritional value | — | PFP inhibits the growth of harmful bacteria while having no significant effect on lactic acid bacteria. | [73] | |
| Noodle Products | noodle | PFP Rice flour corn flour | Improve nutritional value | Damage to cooking characteristics | PFP is more suitable for the development of noodles than a blend of rice flour and corn flour | [74] |
| noodle | PFP | Improve nutritional value | Damage to cooking characteristics | Additives with an addition amount of 6% exhibited the optimal characteristics for noodles. | [75] | |
| Cookie | PFP | Inhibition of harmful microbial growth, extension of product shelf life. | — | The biscuits with an additive content of 30% exhibited the optimal characteristics. | [76] | |
| Bread | PFP-pectin Okara | Improved texture characteristics, enhanced nutritional value | — | PFP-pectin is more suitable for the development of bread compared with soybean dregs. | [77] | |
| Meat Products | Sausage | PFP | Reduce smoking loss, inhibit the growth of harmful bacteria, reduce lipid oxidation, and enhance nutritional value. | Sensory evaluation has been marginally decreased | The sausage with an additive content of 6% exhibited the optimal characteristics. | [78] |
| Surimi | PFP | Improved texture characteristics, reduced cooking loss rate, and enhanced nutritional value. | — | Additives with an addition level of 4% exhibited the optimal properties in surimi. | [79] | |
| Pork burger | PFP | Improved texture characteristics, enhanced cooking yield, inhibition of harmful bacteria growth, inhibition of intestinal bacteria growth, and enhanced nutritional value. | The pH of hamburger meat is critical at the edge of spoilage. | The burger with an additive concentration of 2.5% exhibited the optimal characteristics. | [80] | |
| PFP Green banana | PFP is more suitable than green bananas for the development of hamburger meat. | [81] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Wang, X.; Li, Q.; Kang, X.; Li, Y.; Gong, C.; Liu, Y.; Chen, H. Physiological Functions of the By-Products of Passion Fruit: Processing, Characteristics and Their Applications in Food Product Development. Foods 2025, 14, 1643. https://doi.org/10.3390/foods14091643
Liu Z, Wang X, Li Q, Kang X, Li Y, Gong C, Liu Y, Chen H. Physiological Functions of the By-Products of Passion Fruit: Processing, Characteristics and Their Applications in Food Product Development. Foods. 2025; 14(9):1643. https://doi.org/10.3390/foods14091643
Chicago/Turabian StyleLiu, Zhaohan, Xiaonan Wang, Qianwen Li, Xiaojing Kang, Yan Li, Chunmiao Gong, Yang Liu, and Han Chen. 2025. "Physiological Functions of the By-Products of Passion Fruit: Processing, Characteristics and Their Applications in Food Product Development" Foods 14, no. 9: 1643. https://doi.org/10.3390/foods14091643
APA StyleLiu, Z., Wang, X., Li, Q., Kang, X., Li, Y., Gong, C., Liu, Y., & Chen, H. (2025). Physiological Functions of the By-Products of Passion Fruit: Processing, Characteristics and Their Applications in Food Product Development. Foods, 14(9), 1643. https://doi.org/10.3390/foods14091643