
Creating a Superior Wx Allele with Temperature-Responsive Amylose Regulation and a Novel Transcriptional Pattern in Rice via CRISPR/Cas9-Mediated Promoter Editing
 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Construction of CRISPR/Cas9 Vector and Screening of Homozygous Edited Lines
2.3. Determination of Key Agronomic Traits
2.4. Determination of Physicochemical Properties of Rice Grains
2.5. Grain Transparency Calculation
2.6. Scanning Electron Microscopy Analysis
2.7. Gel Permeation Chromatography Analysis
2.8. Total RNA Extraction and Quantitative Real-Time PCR Analysis
2.9. Western Blot and Enzyme Activity Assay
2.10. Transient Assay of Promoter Activity
2.11. Full-Length Transcriptome Analysis
2.12. Statistical Analysis
3. Results
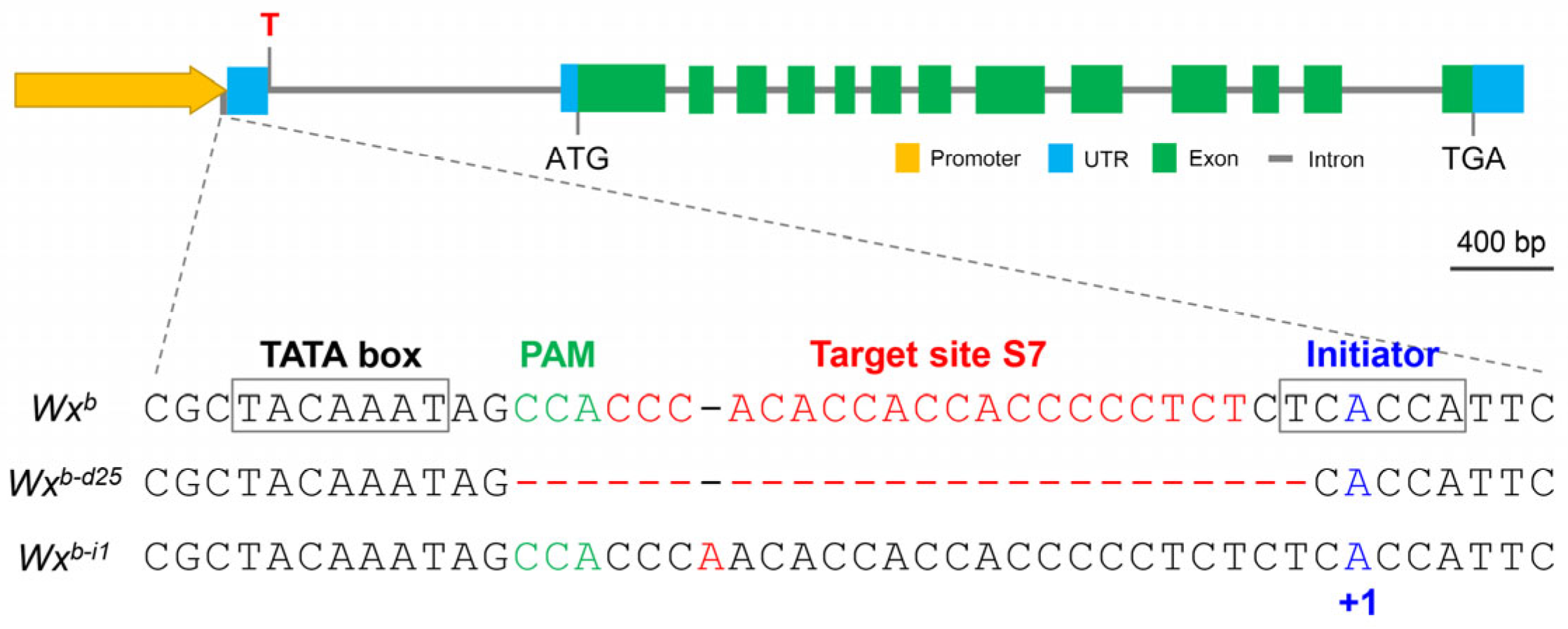
3.1. Creation of a Novel Wx Allele (Wxb-d25) via Promoter Editing
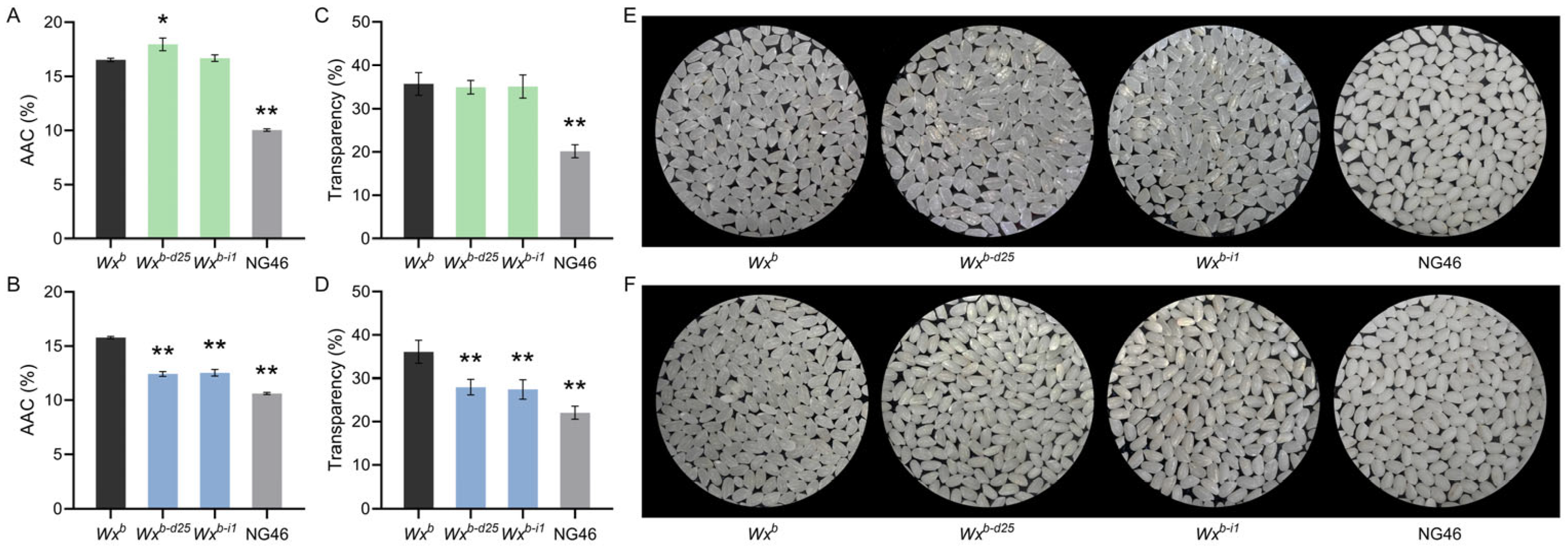
3.2. Wxb-d25 Allele Synergistically Improves Amylose Content and Grain Transparency in Rice
3.3. Wxb-d25 Allele Enhances Rice Grain Quality
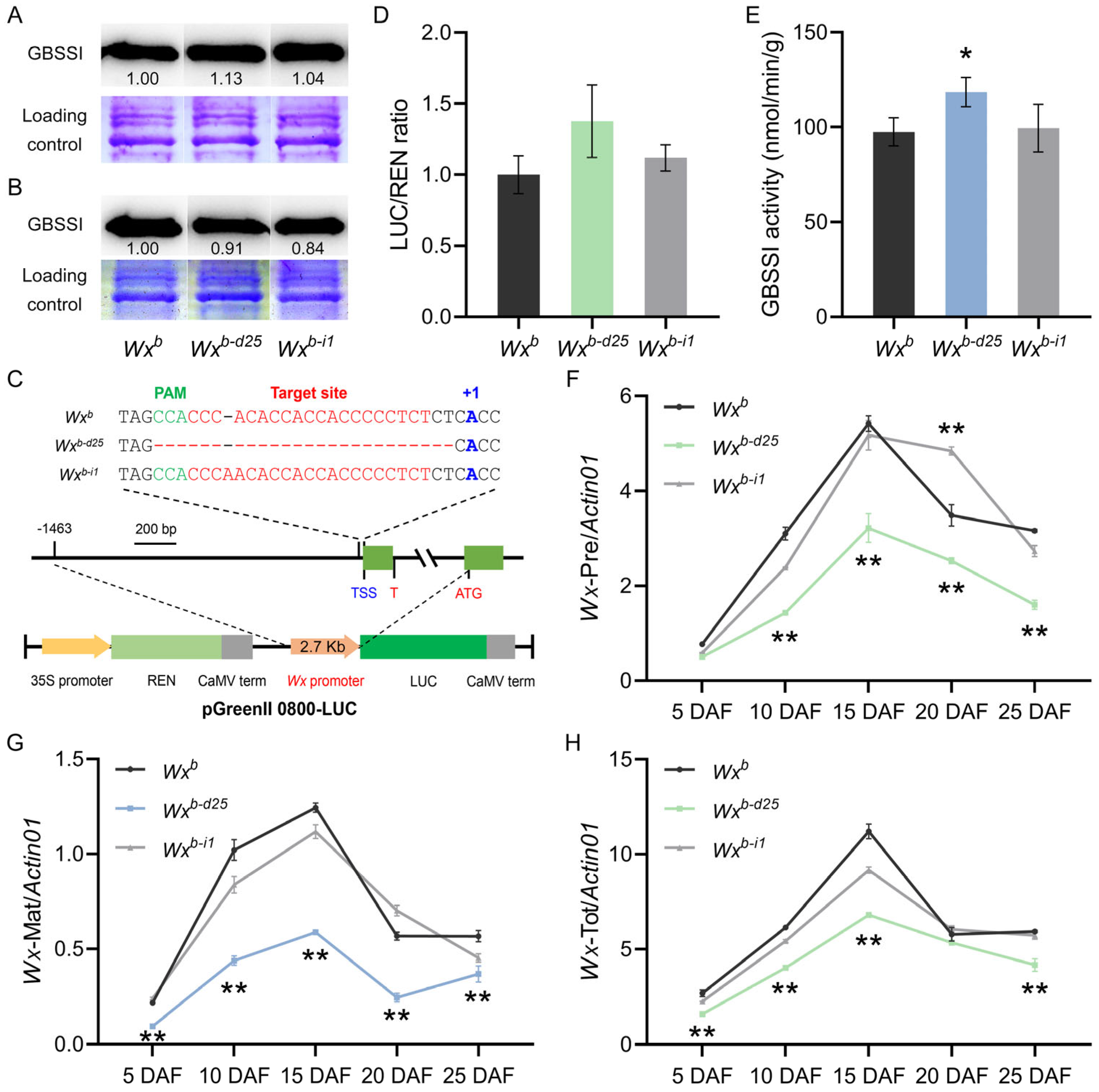
3.4. Expression and Promoter Activity Analysis of Wxb-d25
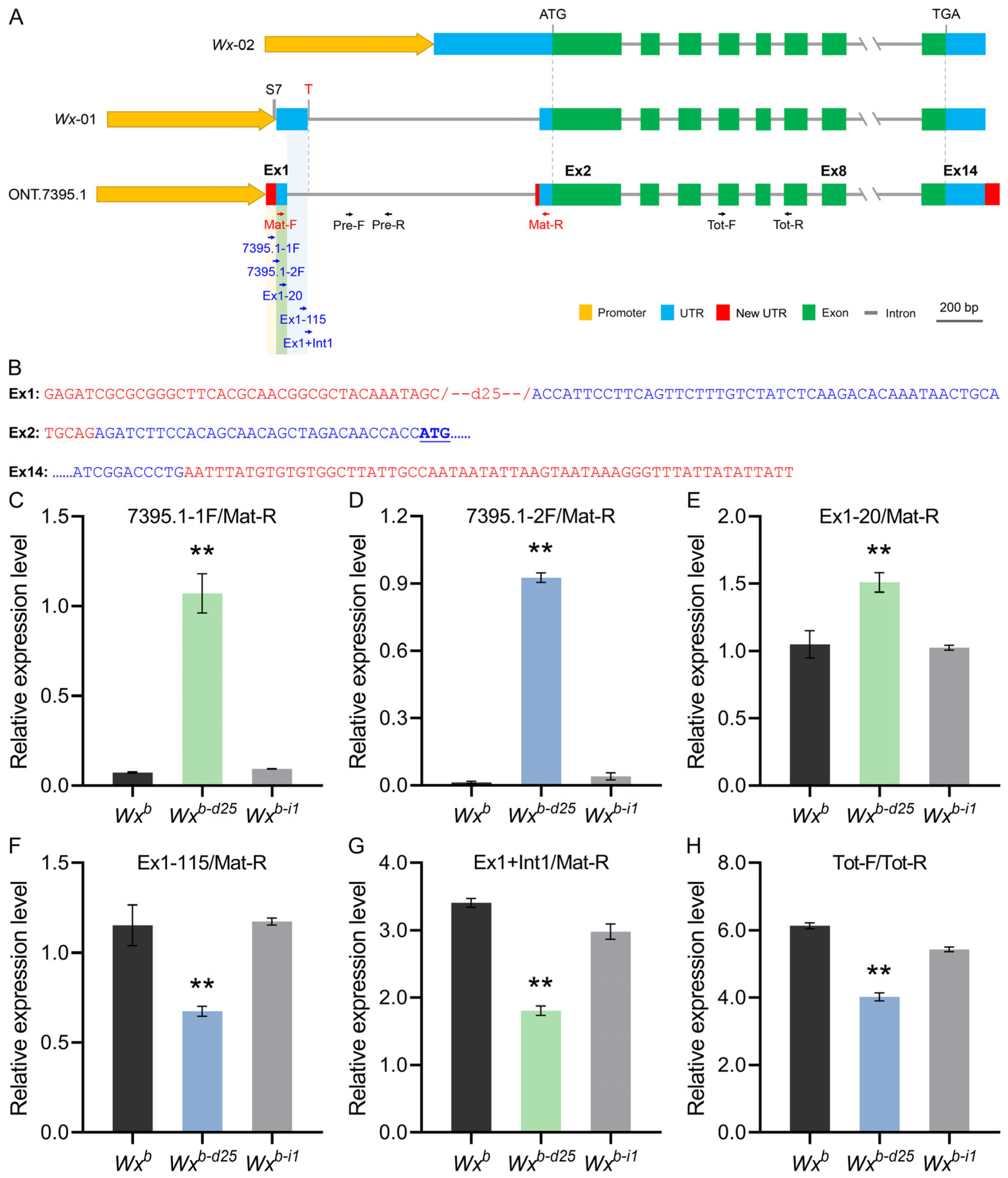
3.5. Full-Length Transcriptome Sequencing Reveals a Novel Transcription Mechanism in Wxb-d25
4. Discussion
4.1. Development of an Elite Wx Allele
4.2. Revelation of a Novel Transcriptional Pattern of the Wx Gene
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ren, D.; Ding, C.; Qian, Q. Molecular bases of rice grain size and quality for optimized productivity. Sci. Bull. 2023, 68, 314–350. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Gilbert, R.G. Starch molecular structure: The basis for an improved understanding of cooked rice texture. Carbohydr. Polym. 2018, 195, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhu, J.; Chen, S.; Fan, X.; Li, Q.; Lu, Y.; Wang, M.; Yu, H.; Yi, C.; Tang, S.; et al. Wxlv, the ancestral allele of rice Waxy gene. Mol. Plant 2019, 12, 1157–1166. [Google Scholar] [CrossRef]
- Seung, D. Amylose in starch: Towards an understanding of biosynthesis, structure and function. New Phytol. 2020, 228, 1490–1504. [Google Scholar] [CrossRef]
- Tan, Y.; Yu, S.; Xing, Y.; Xu, C.; Zhang, Q.; Li, J. The three important traits for cooking and eating quality of rice grains are controlled by a single locus in an elite rice hybrid, Shanyou 63. Theor. Appl. Genet. 1999, 99, 642–648. [Google Scholar] [CrossRef]
- Zaman, S.A.; Sarbini, S.R. The potential of resistant starch as a prebiotic. Crit. Rev. Biotechnol. 2016, 36, 578–584. [Google Scholar] [CrossRef]
- Mikami, I.; Aikawa, M.; Hirano, H.Y.; Sano, Y. Altered tissue-specific expression at the Wx gene of the opaque mutants in rice. Euphytica 1999, 105, 91–97. [Google Scholar] [CrossRef]
- Sasaki, T.; Yasui, T.; Matsuki, J. Effect of amylose content on gelatinization, retrogradation, and pasting properties of starches from waxy and nonwaxy wheat and their F1 seeds. Cereal Chem. 2000, 77, 58–63. [Google Scholar] [CrossRef]
- Tian, Z.; Qian, Q.; Liu, Q.; Yan, M.; Liu, X.; Yan, C.; Liu, G.; Gao, Z.; Tang, S.; Zeng, D.; et al. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities. Proc. Natl. Acad. Sci. USA 2009, 106, 21760–21765. [Google Scholar] [CrossRef]
- Li, H.; Prakash, S.; Nicholson, T.M.; Fitzgerald, M.A.; Gilbert, R.G. The importance of amylose and amylopectin fine structure for textural properties of cooked rice grains. Food Chem. 2016, 196, 702–711. [Google Scholar] [CrossRef]
- Mestres, C.; Briffaz, A.; Valentin, D. Rice cooking and sensory quality. In Rice, 4th ed.; Bao, J., Ed.; AACC International Press: Saint Paul, MN, USA, 2019; pp. 385–426. [Google Scholar]
- Calingacion, M.; Laborte, A.; Nelson, A.; Resurreccion, A.; Concepcion, J.C.; Daygon, V.D.; Mumm, R.; Reinke, R.; Dipti, S.; Bassinello, P.Z.; et al. Diversity of global rice markets and the science required for consumer-targeted rice breeding. PLoS ONE 2014, 9, e85106. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Zhang, Y.; He, L.; Li, C.; Liang, W.; Chen, T.; Zhao, Q.; Zhu, Z.; Zhao, L.; Zhao, C.; et al. Adjusting the amylose content of semi-glutinous japonica rice by genome editing of uORF6 in the Wx gene. Crop J. 2024, 12, 1806–1811. [Google Scholar] [CrossRef]
- Zhou, H.; Xia, D.; Zhao, D.; Li, Y.; Li, P.; Wu, B.; Gao, G.; Zhang, Q.; Wang, G.; Xiao, J.; et al. The origin of Wxla provides new insights into the improvement of grain quality in rice. J. Integr. Plant Biol. 2021, 63, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, C.; Luo, X.; Zhang, T.; Zhang, X.; Liu, P.; Yang, W.; Lei, Y.; Tang, S.; Kang, L.; et al. Fine mapping of the grain chalkiness quantitative trait locus qCGP6 reveals the involvement of Wx in grain chalkiness formation. J. Exp. Bot. 2023, 74, 3544–3559. [Google Scholar] [CrossRef]
- Tu, B.; Zhang, T.; Liu, P.; Yang, W.; Zheng, L.; Dai, Y.; Wang, H.; Lin, S.; Zhang, Z.; Zheng, X.; et al. The LCG1-OsBP5/OsEBP89-Wx module regulates the grain chalkiness and taste quality in rice. Plant Biotechnol. J. 2025, 23, 36–50. [Google Scholar] [CrossRef]
- Huang, L.; Sreenivasulu, N.; Liu, Q. Waxy editing: Old meets new. Trends Plant Sci. 2020, 25, 963–966. [Google Scholar] [CrossRef]
- Wang, A.; Jing, Y.; Cheng, Q.; Zhou, H.; Wang, L.; Gong, W.; Kou, L.; Liu, G.; Meng, X.; Chen, M.; et al. Loss of function of SSIIIa and SSIIIb coordinately confers high RS content in cooked rice. Proc. Natl. Acad. Sci. USA 2023, 120, e2220622120. [Google Scholar] [CrossRef]
- Shen, L.; Li, J.; Li, Y. Resistant starch formation in rice: Genetic regulation and beyond. Plant Commun. 2022, 3, 100329. [Google Scholar] [CrossRef]
- Deng, Z.; Liu, Y.; Gong, C.; Chen, B.; Wang, T. Waxy is an important factor for grain fissure resistance and head rice yield as revealed by a genome-wide association study. J. Exp. Bot. 2022, 73, 6942–6954. [Google Scholar] [CrossRef]
- Cai, Y.; Chen, Z.; Liu, J.; Yu, L.; Wang, Z.; Zhu, S.; Shi, W.; Pan, C.; Wu, Y.; Li, Y.; et al. Genetic improvement of eating and cooking quality of rice cultivars in southern China. Plant Biotechnol. J. 2025, 23, 518–531. [Google Scholar] [CrossRef]
- Wang, Z.; Wu, Z.; Xing, Y.; Zheng, F.; Guo, X.; Zhang, W.; Hong, M. Nucleotide sequence of rice Waxy gene. Nucleic Acids Res. 1990, 18, 5898. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Wang, Z.; Xing, Y.; Zhang, J.; Hong, M. Aberrant splicing of intron 1 leads to the heterogeneous 5′ UTR and decreased expression of Waxy gene in rice cultivars of intermediate amylose content. Plant J. 1998, 14, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Hirano, H.Y.; Eiguchi, M.; Sano, Y. A single base change altered the regulation of the Waxy gene at the posttranscriptional level during the domestication of rice. Mol. Biol. Evol. 1998, 15, 978–987. [Google Scholar] [CrossRef]
- Zeng, D.; Yan, M.; Wang, Y.; Liu, X.; Qian, Q.; Li, J. Du1, encoding a novel Prp1 protein, regulates starch biosynthesis through affecting the splicing of Wxb pre-mRNAs in rice (Oryza sativa L.). Plant Mol. Biol. 2007, 65, 501–509. [Google Scholar] [CrossRef]
- Cai, Y.; Zhang, W.; Fu, Y.; Shan, Z.; Xu, J.; Wang, P.; Kong, F.; Jin, J.; Yan, H.; Ge, X.; et al. Du13 encodes a C2H2 zinc-finger protein that regulates Wxb pre-mRNA splicing and microRNA biogenesis in rice endosperm. Plant Biotechnol. J. 2022, 20, 1387–1401. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, T.; Shi, Y.; Lu, Y.; Li, Q.; Fan, X.; Huang, L.; Chen, L.; Song, X.; Liu, Q.; et al. Introgression of lac6/tl1/du13 improves the palatability of japonica rice. Crop J. 2024, 12, 1259–1265. [Google Scholar] [CrossRef]
- Zhao, G.; Xie, S.; Zong, S.; Wang, T.; Mao, C.; Shi, J.; Li, J. Mutation of TL1, encoding a novel C2H2 zinc finger protein, improves grains eating and cooking quality in rice. Theor. Appl. Genet. 2022, 135, 3531–3543. [Google Scholar] [CrossRef]
- Dung, L.V.; Mikami, I.; Amano, E.; Sano, Y. Study on the response of dull endosperm 2-2, du2-2, to two Wx alleles in rice. Breed. Sci. 2000, 50, 215–219. [Google Scholar] [CrossRef]
- Isshiki, M.; Takasaki, M.A.; Wong, H.L.; Satoh, H.; Shimamoto, K. Du3, a mRNA cap-binding protein gene, regulates amylose content in japonica rice seeds. Plant Biotechnol. 2008, 25, 483–487. [Google Scholar] [CrossRef]
- Takemoto-Kuno, Y.; Mitsueda, H.; Suzuki, K.; Hirabayashi, H.; Ideta, O.; Aoki, N.; Umemoto, T.; Ishii, T.; Ando, I.; Kato, H.; et al. qAC2, a novel QTL that interacts with Wx and controls the low amylose content in rice (Oryza sativa L.). Theor. Appl. Genet. 2015, 128, 563–573. [Google Scholar] [CrossRef]
- Igarashi, H.; Ito, H.; Shimada, T.; Kang, D.-J.; Hamada, S. A novel rice dull gene, LowAC1, encodes an RNA recognition motif protein affecting Waxyb pre-mRNA splicing. Plant Physiol. Biochem. 2021, 162, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Xie, D.; Wang, Z.; Hong, M. Interaction of rice bZIP protein REB with the 5′-upstream region of both rice SBE1 gene and Waxy gene. Chin. Sci. Bull. 2002, 47, 310–314. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Z.; Zhang, Q.; Meng, S.; Wei, C. The NAC transcription factors OsNAC20 and OsNAC26 regulate starch and storage protein synthesis. Plant Physiol. 2020, 184, 1775–1791. [Google Scholar] [CrossRef]
- Bello, B.K.; Hou, Y.; Zhao, J.; Jiao, G.; Wu, Y.; Li, Z.; Wang, Y.; Tong, X.; Wang, W.; Yuan, W.; et al. NF-YB1-YC12-bHLH144 complex directly activates Wx to regulate grain quality in rice (Oryza sativa L.). Plant Biotechnol. J. 2019, 17, 1222–1235. [Google Scholar] [CrossRef]
- Xiong, Y.; Ren, Y.; Li, W.; Wu, F.; Yang, W.; Huang, X.; Yao, J. NF-YC12 is a key multi-functional regulator of accumulation of seed storage substances in rice. J. Exp. Bot. 2019, 70, 3765–3780. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Wang, L.; Li, L.; Liu, Y.; Chong, K.; Theißen, G.; Meng, Z. OsMADS14 and NF-YB1 cooperate in the direct activation of OsAGPL2 and Waxy during starch synthesis in rice endosperm. New Phytol. 2022, 234, 77–92. [Google Scholar] [CrossRef]
- Wang, J.-D.; Wang, J.; Huang, L.-C.; Kan, L.-J.; Wang, C.-X.; Xiong, M.; Zhou, P.; Zhou, L.-H.; Chen, C.; Zhao, D.-S.; et al. ABA-mediated regulation of rice grain quality and seed dormancy via the NF-YB1-SLRL2-bHLH144 Module. Nat. Commun. 2024, 15, 4493. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, H.; Feng, M.; Zhu, Y. Suppression of OsMADS7 in rice endosperm stabilizes amylose content under high temperature stress. Plant Biotechnol. J. 2018, 16, 18–26. [Google Scholar] [CrossRef]
- Fu, F.; Xue, H. Coexpression analysis identifies rice starch regulator1, a rice AP2/EREBP family transcription factor, as a novel rice starch biosynthesis regulator. Plant Physiol. 2010, 154, 927–938. [Google Scholar] [CrossRef]
- Zhu, M.; Liu, Y.; Jiao, G.; Yu, J.; Zhao, R.; Lu, A.; Zhou, W.; Cao, N.; Wu, J.; Hu, S.; et al. The elite eating quality alleles Wx and ALK are regulated by OsDOF18 and coordinately improve head rice yield. Plant Biotechnol. J. 2024, 22, 1582–1595. [Google Scholar] [CrossRef]
- Liu, S.; Wu, J.; Mawia, A.M.; Wei, X.; Cao, R.; Jiao, G.; Wu, Y.; Zhang, J.; Xie, L.; Sheng, Z.; et al. A novel transcription factor OsMYB73 affects grain size and chalkiness by regulating endosperm storage substances’ accumulation-mediated auxin biosynthesis signalling pathway in rice. Plant Biotechnol. J. 2025, 23, 1021–1038. [Google Scholar] [CrossRef] [PubMed]
- Anacleto, R.; Badoni, S.; Parween, S.; Butardo, V.M., Jr.; Misra, G.; Cuevas, R.P.; Kuhlmann, M.; Trinidad, T.P.; Mallillin, A.C.; Acuin, C.; et al. Integrating a genome-wide association study with a large-scale transcriptome analysis to predict genetic regions influencing the glycaemic index and texture in rice. Plant Biotechnol. J. 2019, 17, 1261–1275. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Li, Q.; Zhang, C.; Chu, R.; Gu, Z.; Tan, H.; Zhao, D.; Fan, X.; Liu, Q. Creating novel Wx alleles with fine-tuned amylose levels and improved grain quality in rice by promoter editing using CRISPR/Cas9 system. Plant Biotechnol. J. 2020, 18, 2164–2166. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Liu, T.; Ma, X.; Wang, B.; Zheng, Z.; Zhang, Y.; Xie, X.; Yang, B.; Zhao, Z.; Zhu, Q.; et al. Quantitative regulation of Waxy expression by CRISPR/Cas9-based promoter and 5′UTR-intron editing improves grain quality in rice. Plant Biotechnol. J. 2020, 18, 2385–2387. [Google Scholar] [CrossRef]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef]
- Zhong, Y.; Blennow, A.; Kofoed-Enevoldsen, O.; Jiang, D.; Hebelstrup, K.H. Protein Targeting to Starch 1 is essential for starchy endosperm development in barley. J. Exp. Bot. 2018, 70, 485–496. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, Q.; Zhu, Q.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.; Li, H.; Lin, Y.; et al. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef]
- Bull, S.E.; Seung, D.; Chanez, C.; Mehta, D.; Kuon, J.E.; Truernit, E.; Hochmuth, A.; Zurkirchen, I.; Zeeman, S.C.; Gruissem, W.; et al. Accelerated ex situ breeding of GBSS- and PTST1-edited cassava for modified starch. Sci. Adv. 2018, 4, eaat6086. [Google Scholar] [CrossRef]
- Gao, H.; Gadlage, M.J.; Lafitte, H.R.; Lenderts, B.; Yang, M.; Schroder, M.; Farrell, J.; Snopek, K.; Peterson, D.; Feigenbutz, L.; et al. Superior field performance of waxy corn engineered using CRISPR–Cas9. Nat. Biotechnol. 2020, 38, 579–581. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, R.; Song, G.; Gao, J.; Li, W.; Han, X.; Chen, M.; Li, Y.; Li, G. Targeted mutagenesis using the Agrobacterium tumefaciens-mediated CRISPR-Cas9 system in common wheat. BMC Plant Biol. 2018, 18, 302. [Google Scholar] [CrossRef]
- Li, Q.F.; Huang, L.C.; Chu, R.; Li, J.; Jiang, M.Y.; Zhang, C.Q.; Fan, X.L.; Yu, H.X.; Gu, M.H.; Liu, Q.Q. Down-regulation of SSSII-2 gene expression results in novel low-amylose rice with soft, transparent grains. J. Agric. Food Chem. 2018, 66, 9750–9760. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Shen, L.; Fu, Y.; Yan, C.; Wang, K. A simple CRISPR/Cas9 system for multiplex genome editing in rice. J. Genet. Genom. 2015, 42, 703–706. [Google Scholar] [CrossRef]
- Liu, W.; Xie, X.; Ma, X.; Li, J.; Chen, J.; Liu, Y. DSDecode: A web-based tool for decoding of sequencing chromatograms for genotyping of targeted mutations. Mol. Plant 2015, 8, 1431–1433. [Google Scholar] [CrossRef]
- Xie, X.; Ma, X.; Zhu, Q.; Zeng, D.; Li, G.; Liu, Y. CRISPR-GE: A convenient software toolkit for CRISPR-based genome editing. Mol. Plant 2017, 10, 1246–1249. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, L.; Feng, L.; Jiang, J.; Huang, L.; Liu, Q.; Zhang, Y.; Zhang, C.; Liu, Q. Deciphering the role of Waxy gene mutations in enhancing rice grain quality. Foods 2024, 13, 1624. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.J. Comparison of protease digestion at neutral pH with alkaline steeping method for rice starch isolation. Cereal Chem. 2001, 78, 690–692. [Google Scholar] [CrossRef]
- Huang, L.; Gu, Z.; Chen, Z.; Yu, J.; Chu, R.; Tan, H.; Zhao, D.; Fan, X.; Zhang, C.; Li, Q.; et al. Improving rice eating and cooking quality by coordinated expression of the major starch synthesis-related genes, SSII and Wx, in endosperm. Plant Mol. Biol. 2021, 106, 419–432. [Google Scholar] [CrossRef]
- Xiang, X.; Kang, C.; Xu, S.; Yang, B. Combined effects of Wx and SSIIa haplotypes on rice starch physicochemical properties. J. Sci. Food Agric. 2017, 97, 1229–1234. [Google Scholar] [CrossRef]
- Shu, Q.; Wu, D.; Xia, Y.; Gao, M.; McClung, A. Relationship between RVA profile character and eating quality in Oryza sativa L. Sci. Agric. Sin. 1998, 34, 25–29. [Google Scholar]
- Jia, L.; Ding, X.; Wang, P.; Deng, X. RVA profile characteristics and correlation with the physical/chemical quality. Acta Agron. Sin. 2008, 34, 790–794. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, L.; Liu, G.; Meng, X.; Jing, Y.; Shu, X.; Kong, X.; Sun, J.; Yu, H.; Smith, S.M.; et al. Critical roles of soluble starch synthase SSIIIa and granule-bound starch synthase Waxy in synthesizing resistant starch in rice. Proc. Natl. Acad. Sci. USA 2016, 113, 12844–12849. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wei, X.; Jiao, G.; Chen, W.; Wu, Y.; Sheng, Z.; Hu, S.; Xie, L.; Wang, J.; Tang, S.; et al. GBSS-BINDING PROTEIN, encoding a CBM48 domain-containing protein, affects rice quality and yield. J. Integr. Plant Biol. 2020, 62, 948–966. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Jiao, G.; Cao, R.; Li, S.; Zhao, S.; Duan, Y.; Ma, L.; Li, X.; Lu, F.; Wang, H.; et al. OsLESV and OsESV1 promote transitory and storage starch biosynthesis to determine rice grain quality and yield. Plant Commun. 2024, 5, 100893. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhang, W.; Wang, Y.; Jin, J.; Xu, H.; Fu, Y.; Shan, Z.; Wang, X.; Teng, X.; Li, X.; et al. Rice LIKE EARLY STARVATION1 cooperates with FLOURY ENDOSPERM6 to modulate starch biosynthesis and endosperm development. Plant Cell 2024, 36, 1892–1912. [Google Scholar] [CrossRef]
- Sun, Y.; Jiao, G.; Liu, Z.; Zhang, X.; Li, J.; Guo, X.; Du, W.; Du, J.; Francis, F.; Zhao, Y.; et al. Generation of high-amylose rice through CRISPR/Cas9-mediated targeted mutagenesis of starch branching enzymes. Front. Plant Sci. 2017, 8, 298. [Google Scholar] [CrossRef]
- Yan, M.; Pan, T.; Zhu, Y.; Jiang, X.; Yu, M.; Wang, R.; Zhang, F.; Luo, S.; Bao, X.; Chen, Y.; et al. FLOURY ENDOSPERM20 encoding SHMT4 is required for rice endosperm development. Plant Biotechnol. J. 2022, 20, 1438–1440. [Google Scholar] [CrossRef]
- Wu, H.; Ren, Y.; Dong, H.; Xie, C.; Zhao, L.; Wang, X.; Zhang, F.; Zhang, B.; Jiang, X.; Huang, Y.; et al. FLOURY ENDOSPERM24, a heat shock protein 101 (HSP101), is required for starch biosynthesis and endosperm development in rice. New Phytol. 2024, 242, 2635–2651. [Google Scholar] [CrossRef]
- Wu, B.; Yun, P.; Zhou, H.; Xia, D.; Gu, Y.; Li, P.; Yao, J.; Zhou, Z.; Chen, J.; Liu, R.; et al. Natural variation in WHITE-CORE RATE 1 regulates redox homeostasis in rice endosperm to affect grain quality. Plant Cell 2022, 34, 1912–1932. [Google Scholar] [CrossRef]
- Qin, P.; Zhang, G.; Hu, B.; Wu, J.; Chen, W.; Ren, Z.; Liu, Y.; Xie, J.; Yuan, H.; Tu, B.; et al. Leaf-derived ABA regulates rice seed development via a transporter-mediated and temperature-sensitive mechanism. Sci. Adv. 2021, 7, eabc8873. [Google Scholar] [CrossRef]
- Ma, B.; Zhang, L.; Gao, Q.; Wang, J.; Li, X.; Wang, H.; Liu, Y.; Lin, H.; Liu, J.; Wang, X.; et al. A plasma membrane transporter coordinates phosphate reallocation and grain filling in cereals. Nat. Genet. 2021, 53, 906–915. [Google Scholar] [CrossRef]
- Fujita, N.; Miura, S.; Crofts, N. Effects of various allelic combinations of starch biosynthetic genes on the properties of endosperm sarch in rice. Rice 2022, 15, 24. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Tan, H.; Zhang, C.; Li, Q.; Liu, Q. Starch biosynthesis in cereal endosperms: An updated review over the last decade. Plant Commun. 2021, 2, 100237. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Li, C.; Gao, C. Applications of CRISPR–Cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 2020, 21, 661–677. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, H.; Botella, J.R.; Zhu, J.K. Generation of new glutinous rice by CRISPR/Cas9-targeted mutagenesis of the Waxy gene in elite rice varieties. J. Integr. Plant Biol. 2018, 60, 369–375. [Google Scholar] [CrossRef]
- Xu, Y.; Lin, Q.; Li, X.; Wang, F.; Chen, Z.; Wang, J.; Li, W.; Fan, F.; Tao, Y.; Jiang, Y.; et al. Fine-tuning the amylose content of rice by precise base editing of the Wx gene. Plant Biotechnol. J. 2021, 19, 11–13. [Google Scholar] [CrossRef]
- Huang, X.; Su, F.; Huang, S.; Mei, F.; Niu, X.; Ma, C.; Zhang, H.; Zhu, X.; Zhu, J.-K.; Zhang, J. Novel Wx alleles generated by base editing for improvement of rice grain quality. J. Integr. Plant Biol. 2021, 63, 1632–1638. [Google Scholar] [CrossRef]
- Liu, X.; Ding, Q.; Wang, W.; Pan, Y.; Tan, C.; Qiu, Y.; Chen, Y.; Li, H.; Li, Y.; Ye, N.; et al. Targeted deletion of the first intron of the Wxb allele via CRISPR/Cas9 significantly increases grain amylose content in rice. Rice 2022, 15, 1. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, G.; Zhao, Y.; Zhang, R.; Tang, X.; Li, L.; Jia, X.; Guo, Y.; Wu, Y.; Han, Y.; et al. An efficient CRISPR–Cas12a promoter editing system for crop improvement. Nat. Plants 2023, 9, 588–604. [Google Scholar] [CrossRef]
- Haberle, V.; Stark, A. Eukaryotic core promoters and the functional basis of transcription initiation. Nat. Rev. Mol. Cell Biol. 2018, 19, 621–637. [Google Scholar] [CrossRef]
- Lee, T.I.; Young, R.A. Transcription of eukaryotic protein-coding genes. Annu. Rev. Genet. 2000, 34, 77–137. [Google Scholar] [CrossRef]
- Danino, Y.M.; Even, D.; Ideses, D.; Juven-Gershon, T. The core promoter: At the heart of gene expression. Biochim. Biophys. Acta Gene Regul. Mech. 2015, 1849, 1116–1131. [Google Scholar] [CrossRef] [PubMed]
- Juven, G.T.; Kadonaga, J.T. Regulation of gene expression via the core promoter and the basal transcriptional machinery. Dev. Biol. 2010, 339, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Sandelin, A.; Carninci, P.; Lenhard, B.; Ponjavic, J.; Hayashizaki, Y.; Hume, D.A. Mammalian RNA polymerase II core promoters: Insights from genome-wide studies. Nat. Rev. Genet. 2007, 8, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Carninci, P.; Sandelin, A.; Lenhard, B.; Katayama, S.; Shimokawa, K.; Ponjavic, J.; Semple, C.A.M.; Taylor, M.S.; Engström, P.G.; Frith, M.C.; et al. Genome-wide analysis of mammalian promoter architecture and evolution. Nat. Genet. 2006, 38, 626–635. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | PKV (cP) | HPV (cP) | BDV (cP) | CPV (cP) | SBV (cP) | PeT (Min) | PaT (°C) |
|---|---|---|---|---|---|---|---|
| Wxb-YZ | 3430.2 ± 55.7 | 1965.1 ± 127.2 | 1468.5 ± 71.5 | 3430.7 ± 117.2 | 3.9 ± 62.2 | 5.7 ± 0.1 | 79.7 ± 0.8 |
| Wxb-d25-YZ | 3413.8 ± 14.5 | 1747.6 ± 10.0 * | 1669.6 ± 22.6 * | 3077.4 ± 107.2 ** | −331.1 ± 94.2 ** | 5.8 ± 0.0 | 76.6 ± 0.0 ** |
| Wxb-i1-YZ | 3423.2 ± 72.2 | 1987.2 ± 7.4 | 1439.4 ± 77.2 | 3298.7 ± 19.6 | −121.2 ± 90.5 | 6.0 ± 0.0 ** | 78.1 ± 0.8 |
| Wxb-LS | 2978.4 ± 6.2 | 2051.0 ± 11.3 | 930.7 ± 13.6 | 3725.4 ± 33.9 | 750.4 ± 31.9 | 6.0 ± 0.0 | 87.7 ± 0.1 |
| Wxb-d25-LS | 2944.2 ± 48.6 | 1859.0 ± 10.4 * | 1088.5 ± 39.7 * | 3278.3 ± 16.2 ** | 337.4 ± 33.9 ** | 5.8 ± 0.0 ** | 81.4 ± 3.1 ** |
| Wxb-i1-LS | 2495.3 ± 242.1 ** | 2003.6 ± 141.0 | 795.0 ± 101.0 ** | 3437.8 ± 152.5 * | 645.8 ± 90.2 * | 6.1 ± 0.0 ** | 88.6 ± 0.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, J.; Yu, J.; Shen, H.; Zhou, L.; Chen, Z.; Fan, X.; Li, Q.; Zhang, C.; Liu, Q.; Huang, L.; et al. Creating a Superior Wx Allele with Temperature-Responsive Amylose Regulation and a Novel Transcriptional Pattern in Rice via CRISPR/Cas9-Mediated Promoter Editing. Foods 2025, 14, 1330. https://doi.org/10.3390/foods14081330
Yan J, Yu J, Shen H, Zhou L, Chen Z, Fan X, Li Q, Zhang C, Liu Q, Huang L, et al. Creating a Superior Wx Allele with Temperature-Responsive Amylose Regulation and a Novel Transcriptional Pattern in Rice via CRISPR/Cas9-Mediated Promoter Editing. Foods. 2025; 14(8):1330. https://doi.org/10.3390/foods14081330
Chicago/Turabian StyleYan, Jiali, Jiawen Yu, Huimin Shen, Lihui Zhou, Zhuanzhuan Chen, Xiaolei Fan, Qianfeng Li, Changquan Zhang, Qing Liu, Lichun Huang, and et al. 2025. "Creating a Superior Wx Allele with Temperature-Responsive Amylose Regulation and a Novel Transcriptional Pattern in Rice via CRISPR/Cas9-Mediated Promoter Editing" Foods 14, no. 8: 1330. https://doi.org/10.3390/foods14081330
APA StyleYan, J., Yu, J., Shen, H., Zhou, L., Chen, Z., Fan, X., Li, Q., Zhang, C., Liu, Q., Huang, L., & Liu, Q. (2025). Creating a Superior Wx Allele with Temperature-Responsive Amylose Regulation and a Novel Transcriptional Pattern in Rice via CRISPR/Cas9-Mediated Promoter Editing. Foods, 14(8), 1330. https://doi.org/10.3390/foods14081330